
Clock at the Core of Cancer Development



Abstract
:Simple Summary
Abstract
1. Introduction
2. Method
3. Results and Discussion
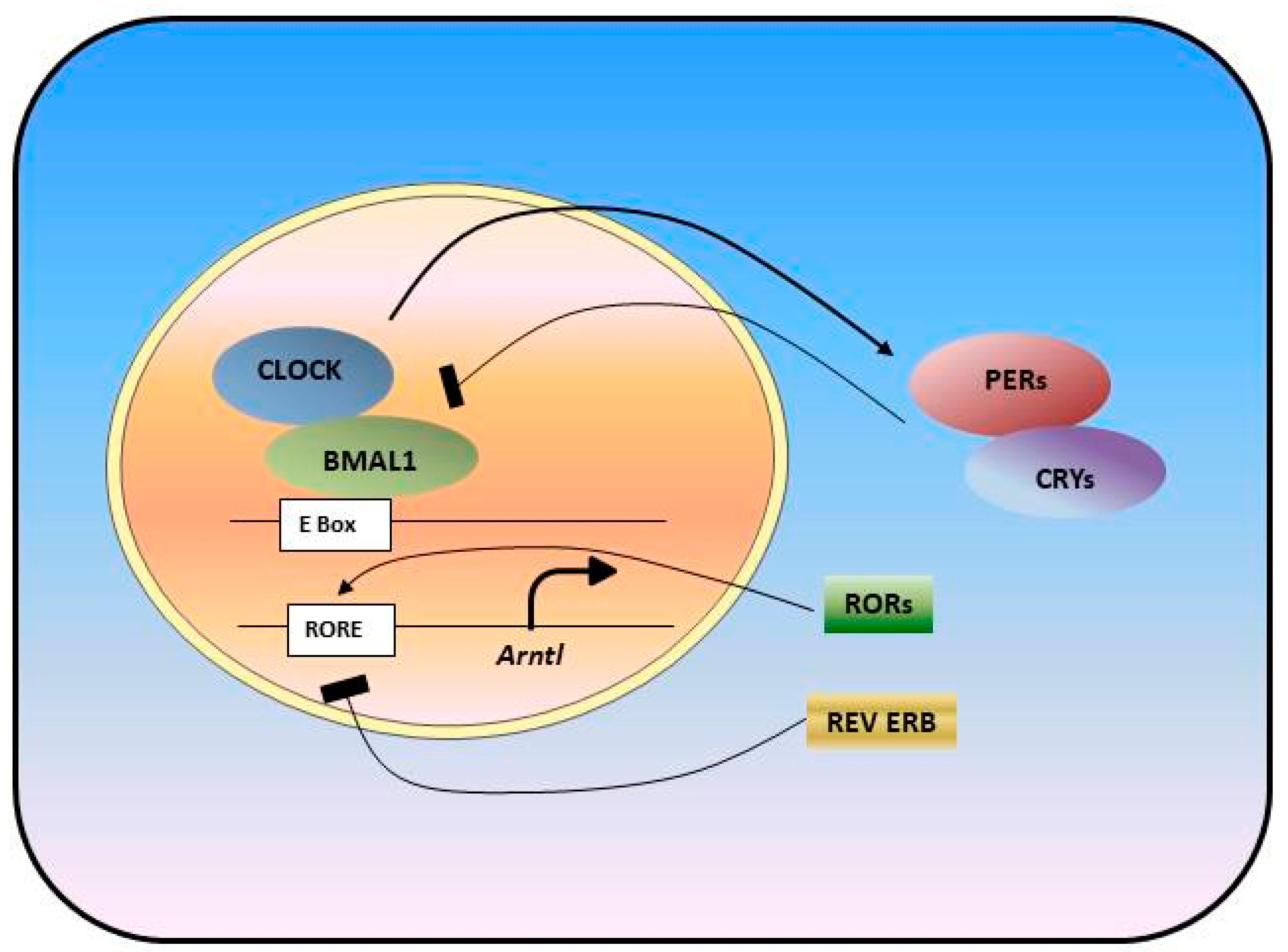
3.1. Circadian Disruption in Cancer
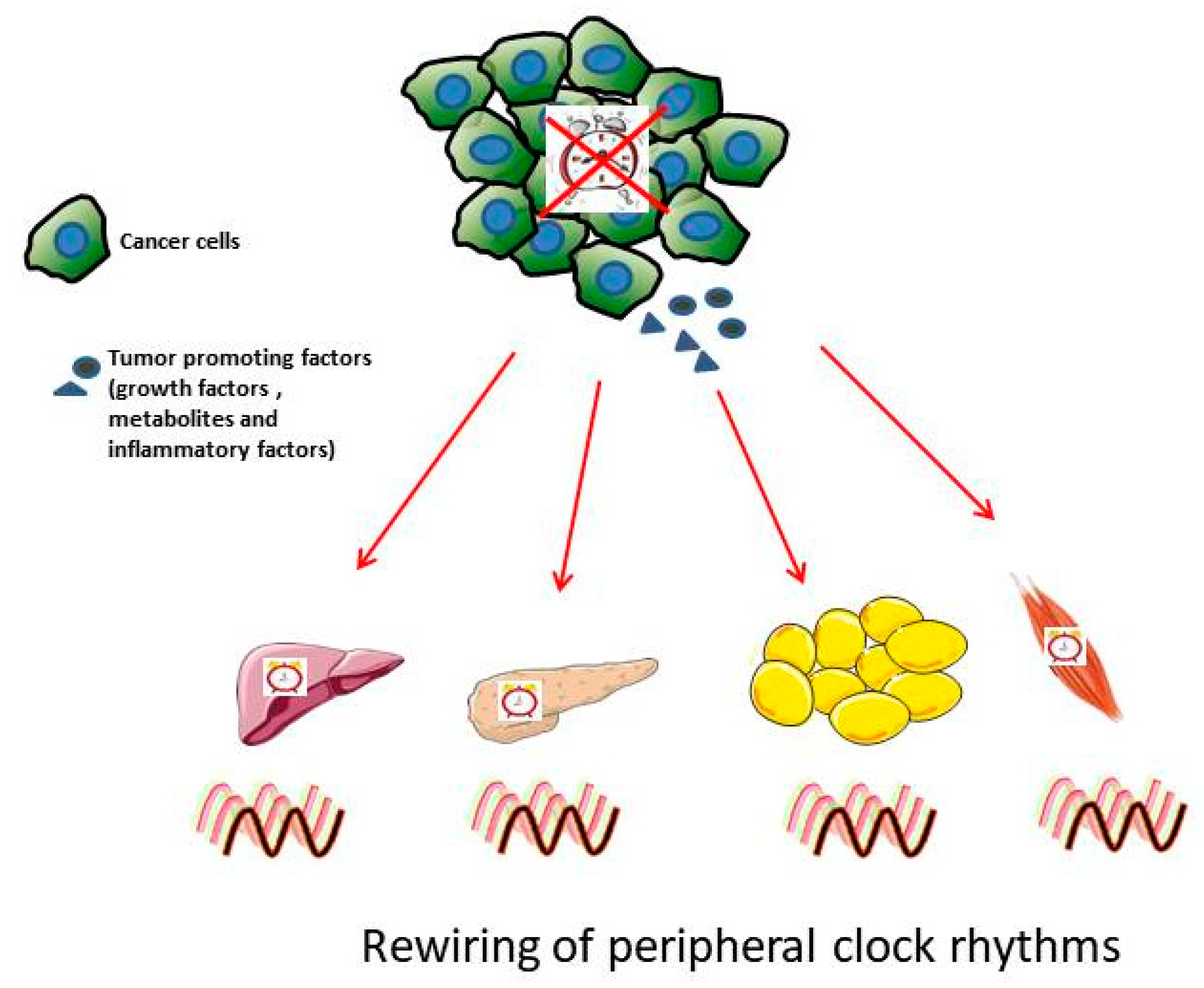
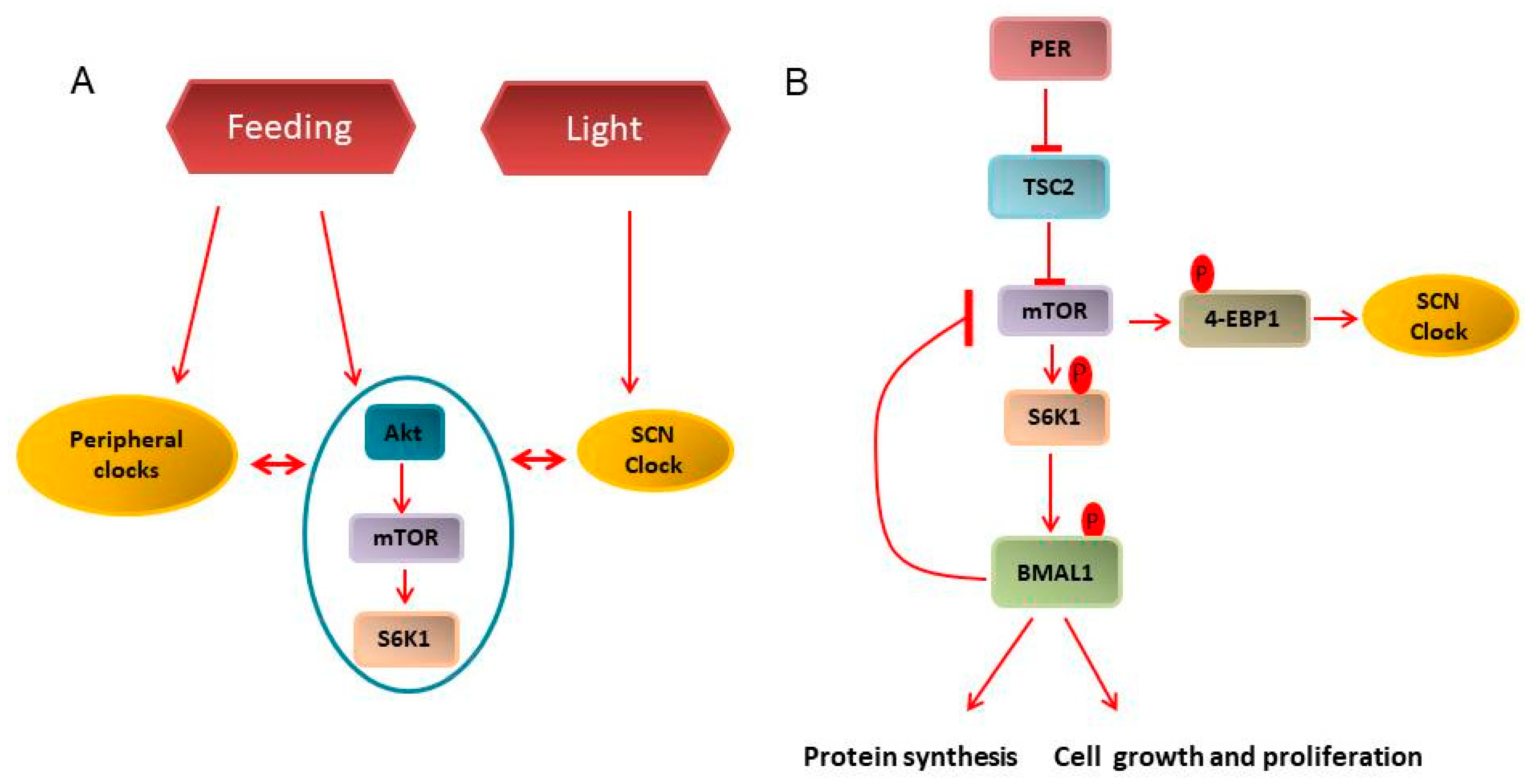
3.2. Clock and Cancer Metabolism
3.3. Circadian Strategies for Cancer Treatment
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sancar, G.; Brunner, M. Circadian Clocks and Energy Metabolism. Cell. Mol. Life Sci. 2014, 71, 2667–2680. [Google Scholar] [CrossRef]
- Fu, L.; Kettner, N.M. The Circadian Clock in Cancer Development and Therapy. Prog. Mol. Biol. Transl. Sci. 2013, 119, 221–282. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kettner, N.M.; Voicu, H.; Finegold, M.J.; Coarfa, C.; Sreekumar, A.; Putluri, N.; Katchy, C.A.; Lee, C.; Moore, D.D.; Fu, L. Circadian Homeostasis of Liver Metabolism Suppresses Hepatocarcinogenesis. Cancer Cell 2016, 30, 909–924. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Welsh, D.K.; Takahashi, J.S.; Kay, S.A. Suprachiasmatic Nucleus: Cell Autonomy and Network Properties. Annu. Rev. Physiol. 2009, 72, 551–577. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stevens, R.G.; Blask, D.E.; Brainard, G.C.; Hansen, J.; Lockley, S.W.; Provencio, I.; Rea, M.S.; Reinlib, L. Meeting Report: The Role of Environmental Lighting and Circadian Disruption in Cancer and Other Diseases. Environ. Health Perspect. 2007, 115, 1357–1362. [Google Scholar] [CrossRef] [Green Version]
- Buhr, E.D.; Takahashi, J.S. Molecular Components of the Mammalian Circadian Clock. Handb. Exp. Pharmacol. 2013, 217, 3–27. [Google Scholar] [CrossRef]
- Ko, C.H.; Takahashi, J.S. Molecular Components of the Mammalian Circadian Clock. Hum. Mol. Genet. 2006, 15. [Google Scholar] [CrossRef]
- Miller, B.H.; McDearmon, E.L.; Panda, S.; Hayes, K.R.; Zhang, J.; Andrews, J.L.; Antoch, M.P.; Walker, J.R.; Esser, K.A.; Hogenesch, J.B.; et al. Circadian and CLOCK-Controlled Regulation of the Mouse Transcriptome and Cell Proliferation. Proc. Natl. Acad. Sci. USA 2007, 104, 3342–3347. [Google Scholar] [CrossRef] [Green Version]
- Reppert, S.M.; Weaver, D.R. Molecular Analysis of Mammalian Circadian Rhythms. Annu. Rev. Physiol. 2001, 63, 647–676. [Google Scholar] [CrossRef]
- Duong, H.A.; Robles, M.S.; Knutti, D.; Weitz, C.J. A Molecular Mechanism for Circadian Clock Negative Feedback. Science 2011, 332, 1436–1439. [Google Scholar] [CrossRef] [Green Version]
- Preitner, N.; Damiola, F.; Lopez-Molina, L.; Zakany, J.; Duboule, D.; Albrecht, U.; Schibler, U. The Orphan Nuclear Receptor REV-ERBalpha Controls Circadian Transcription within the Positive Limb of the Mammalian Circadian Oscillator. Cell 2002, 110, 251–260. [Google Scholar] [CrossRef]
- Sato, T.K.; Panda, S.; Miraglia, L.J.; Reyes, T.M.; Rudic, R.D.; McNamara, P.; Naik, K.A.; FitzGerald, G.A.; Kay, S.A.; Hogenesch, J.B. A Functional Genomics Strategy Reveals Rora as a Component of the Mammalian Circadian Clock. Neuron 2004, 43, 527–537. [Google Scholar] [CrossRef] [Green Version]
- Asher, G.; Sassone-Corsi, P. Time for Food: The Intimate Interplay between Nutrition, Metabolism, and the Circadian Clock. Cell 2015, 161, 84–92. [Google Scholar] [CrossRef] [Green Version]
- Masri, S.; Sassone-Corsi, P. The Emerging Link between Cancer, Metabolism, and Circadian Rhythms. Nat. Med. 2018, 24, 1795–1803. [Google Scholar] [CrossRef] [PubMed]
- Schernhammer, E.S.; Laden, F.; Speizer, F.E.; Willet, W.C.; Hunter, D.J.; Kawachi, I.; Fuchs, C.S.; Colditz, G.A. Night-Shift Work and Risk of Colorectal Cancer in the Nurses’ Health Study. J. Natl. Cancer Inst. 2003, 95, 825–828. [Google Scholar] [CrossRef] [Green Version]
- Srour, B.; Plancoulaine, S.; Andreeva, V.A.; Fassier, P.; Julia, C.; Galan, P.; Hercberg, S.; Deschasaux, M.; Latino-Martel, P.; Touvier, M. Circadian Nutritional Behaviours and Cancer Risk: New Insights from the NutriNet-Santé Prospective Cohort Study: Disclaimers. Int. J. Cancer 2018, 143, 2369–2379. [Google Scholar] [CrossRef] [Green Version]
- Papantoniou, K.; Castaño-Vinyals, G.; Espinosa, A.; Aragonés, N.; Pérez-Gõmez, B.; Burgos, J.; Gõmez-Acebo, I.; Llorca, J.; Peirõ, R.; Jimenez-Moleõn, J.J.; et al. Night Shift Work, Chronotype and Prostate Cancer Risk in the MCC-Spain Case-Control Study. Int. J. Cancer 2015, 137, 1147–1157. [Google Scholar] [CrossRef]
- Knutsson, A.; Alfredsson, L.; Karlsson, B.; Åkerstedt, T.; Fransson, E.I.; Westerholm, P.; Westerlund, H. Breast Cancer among Shift Workers: Results of the WOLF Longitudinal Cohort Study. Scand. J. Work. Environ. Health 2013, 39, 170–177. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sancar, A.; Lindsey-Boltz, L.A.; Gaddameedhi, S.; Selby, C.P.; Ye, R.; Chiou, Y.Y.; Kemp, M.G.; Hu, J.; Lee, J.H.; Ozturk, N. Circadian Clock, Cancer, and Chemotherapy. Biochemistry 2015, 54, 110–123. [Google Scholar] [CrossRef]
- Sancar, A. Mechanisms of DNA Repair by Photolyase and Excision Nuclease (Nobel Lecture). Angew. Chemie Int. Ed. 2016, 55, 8502–8527. [Google Scholar] [CrossRef] [PubMed]
- Papagiannakopoulos, T.; Bauer, M.R.; Davidson, S.M.; Heimann, M.; Subbaraj, L.; Bhutkar, A.; Bartlebaugh, J.; Vander Heiden, M.G.; Jacks, T. Circadian Rhythm Disruption Promotes Lung Tumorigenesis. Cell Metab. 2016, 24, 324–331. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, F.; Li, C.; Yongluo; Chen, L. The Circadian Gene Clock Plays an Important Role in Cell Apoptosis and the DNA Damage Response In Vitro. Technol. Cancer Res. Treat. 2015, 15, 480–486. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.; Donehower, L.A.; Herron, A.J.; Moore, D.D.; Fu, L. Disrupting Circadian Homeostasis of Sympathetic Signaling Promotes Tumor Development in Mice. PLoS ONE 2010, 5. [Google Scholar] [CrossRef] [Green Version]
- Wendeu-Foyet, M.G.; Menegaux, F. Circadian Disruption and Prostate Cancer Risk: An Updated Review of Epidemiological Evidences. Cancer Epidemiol. Biomark. Prev. 2017, 26, 985–991. [Google Scholar] [CrossRef] [Green Version]
- Dickerman, B.A.; Markt, S.C.; Koskenvuo, M.; Hublin, C.; Pukkala, E.; Mucci, L.A.; Kaprio, J. Sleep Disruption, Chronotype, Shift Work, and Prostate Cancer Risk and Mortality: A 30-Year Prospective Cohort Study of Finnish Twins. Cancer Causes Control. 2016, 27, 1361–1370. [Google Scholar] [CrossRef]
- Markt, S.C.; Flynn-Evans, E.E.; Valdimarsdottir, U.A.; Sigurdardottir, L.G.; Tamimi, R.M.; Batista, J.L.; Haneuse, S.; Lockley, S.W.; Stampfer, M.; Wilson, K.M.; et al. Sleep Duration and Disruption and Prostate Cancer Risk: A 23-Year Prospective Study. Cancer Epidemiol. Biomark. Prev. 2016, 25, 302–308. [Google Scholar] [CrossRef] [Green Version]
- Sigurdardottir, L.G.; Valdimarsdottir, U.A.; Fall, K.; Rider, J.R.; Lockley, S.W.; Schernhammer, E.; Mucci, L.A. Circadian Disruption, Sleep Loss, and Prostate Cancer Risk: A Systematic Review of Epidemiologic Studies. Cancer Epidemiol. Biomark. Prev. 2012, 21, 1002–1011. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Erren, T.C.; Morfeld, P.; Groß, J.V.; Wild, U.; Lewis, P. IARC 2019: “Night Shift Work” Is Probably Carcinogenic: What about Disturbed Chronobiology in All Walks of Life? J. Occup. Med. Toxicol. 2019, 14. [Google Scholar] [CrossRef] [PubMed]
- Hanahan, D.; Weinberg, R.A. Hallmarks of Cancer: The next Generation. Cell 2011, 144, 646–674. [Google Scholar] [CrossRef] [Green Version]
- Hansen, J. Increased Breast Cancer Risk among Women Who Work Predominantly at Night. Epidemiology 2001, 12, 74–77. [Google Scholar] [CrossRef]
- Schernhammer, E.S.; Kroenke, C.H.; Laden, F.; Hankinson, S.E. Night Work and Risk of Breast Cancer. Epidemiology 2006, 17, 108–111. [Google Scholar] [CrossRef]
- Hansen, J. Night Shift Work and Risk of Breast Cancer. Curr. Environ. Health Rep. 2017, 4, 325–339. [Google Scholar] [CrossRef]
- Leonardi, G.C.; Rapisarda, V.; Marconi, A.; Scalisi, A.; Catalano, F.; Proietti, L.; Travali, S.; Libra, M.; Fenga, C. Correlation of the Risk of Breast Cancer and Disruption of the Circadian Rhythm (Review). Oncol. Rep. 2012, 28, 418–428. [Google Scholar] [CrossRef]
- Stevens, R.G.; Rea, M.S. Light in the Built Environment: Potential Role of Circadian Disruption in Endocrine Disruption and Breast Cancer. Cancer Causes Control. 2001, 12, 279–287. [Google Scholar] [CrossRef]
- Berson, D.M.; Dunn, F.A.; Takao, M. Phototransduction by Retinal Ganglion Cells That Set the Circadian Clock. Science 2002, 295, 1070–1073. [Google Scholar] [CrossRef] [Green Version]
- Stevens, R. Circadian Disruption and Breast Cancer. Epidemiology 2005, 16, 711. [Google Scholar] [CrossRef]
- Kos-Kudla, B.; Ostrowska, Z.; Kozlowski, A.; Marek, B.; Ciesielska-Kopacz, N.; Kudla, M.; Kajdaniuk, D.; Strzelczyk, J.; Staszewicz, P. Circadian Rhythm of Melatonin in Patients with Colorectal Carcinoma. Neuroendocrinol. Lett. 2002, 23, 239–242. [Google Scholar] [PubMed]
- Kakizaki, M.; Inoue, K.; Kuriyama, S.; Sone, T.; Matsuda-Ohmori, K.; Nakaya, N.; Fukudo, S.; Tsuji, I. Sleep Duration and the Risk of Prostate Cancer: The Ohsaki Cohort Study. Br. J. Cancer 2008, 99, 176–178. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rafnsson, V.; Tulinius, H.; Jonasson, J.G.; Hrafnkelsson, J. Risk of Breast Cancer in Female Flight Attendants: A Population-Based Study (Iceland). Cancer Causes Control. 2001, 12, 95–101. [Google Scholar] [CrossRef] [PubMed]
- Tokumaru, O.; Haruki, K.; Bacal, K.; Katagiri, T.; Yamamoto, T.; Sakurai, Y. Incidence of Cancer among Female Flight Attendants: A Meta-Analysis. J. Travel Med. 2006, 13, 127–132. [Google Scholar] [CrossRef] [PubMed]
- Pukkala, E.; Aspholm, R.; Auvinen, A.; Eliasch, H.; Gundestrup, M.; Haldorsen, T.; Hammar, N.; Ukkala, P.E.; Spholm, A.R.; Uvinen, A.A.; et al. Cancer Incidence Among 10,211 Airline Pilots: A Nordic study. Aviat. Space Environ. Med. 2003, 74, 699–706. [Google Scholar]
- Filipski, E.; King, V.M.; Li, X.; Granda, G.T.; Mormont, M.-C.; Liu, X.H.; Claustrat, B.; Hastings, M.H. Host Circadian Clock as a Control Point in Tumor Progression. J. Natl. Cancer Inst. 2002, 94, 690–697. [Google Scholar] [CrossRef]
- Filipski, E.; Delaunay, F.; King, V.M.; Wu, M.W.; Claustrat, B.; Gréchez-Cassiau, A.; Guettier, C.; Hastings, M.H.; Francis, L. Effects of Chronic Jet Lag on Tumor Progression in Mice. Cancer Res. 2004, 64, 7879–7885. [Google Scholar] [CrossRef] [Green Version]
- Fu, L.; Pelicano, H.; Liu, J.; Huang, P.; Lee, C.C. The Circadian Gene Period2 Plays an Important Role in Tumor Suppression and DNA Damage Response in Vivo. Cell 2002, 111, 41–50. [Google Scholar] [CrossRef] [Green Version]
- Reszka, E.; Zienolddiny, S. Epigenetic Basis of Circadian Rhythm Disruption in Cancer. Methods Mol. Biol. 2018, 1856, 173–201. [Google Scholar] [CrossRef]
- Masri, S.; Kinouchi, K.; Sassone-Corsi, P. Circadian Clocks, Epigenetics, and Cancer. Curr. Opin. Oncol. 2015, 27, 50–56. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Joska, T.M.; Zaman, R.; Belden, W.J. Regulated DNA Methylation and the Circadian Clock: Implications in Cancer. Biology 2014, 3, 560–577. [Google Scholar] [CrossRef] [Green Version]
- Ye, Y.; Xiang, Y.; Ozguc, F.M.; Kim, Y.; Liu, C.J.; Park, P.K.; Hu, Q.; Diao, L.; Lou, Y.; Lin, C.; et al. The Genomic Landscape and Pharmacogenomic Interactions of Clock Genes in Cancer Chronotherapy. Cell Syst. 2018, 6, 314–328.e2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.; Zou, J.X.; Xue, X.; Cai, D.; Zhang, Y.; Duan, Z.; Xiang, Q.; Yang, J.C.; Louie, M.C.; Borowsky, A.D.; et al. ROR-γ Drives Androgen Receptor Expression and Represents a Therapeutic Target in Castration-Resistant Prostate Cancer. Nat. Med. 2016, 22, 488–496. [Google Scholar] [CrossRef]
- Anafi, R.C.; Francey, L.J.; Hogenesch, J.B.; Kim, J. CYCLOPS Reveals Human Transcriptional Rhythms in Health and Disease. Proc. Natl. Acad. Sci. USA 2017, 114, 5312–5317. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Assis, L.V.M.; Kinker, G.S.; Moraes, M.N.; Markus, R.P.; Fernandes, P.A.; De Lauro Castrucci, A.M. Expression of the Circadian Clock Gene BMAL1 Positively Correlates with Antitumor Immunity and Patient Survival in Metastatic Melanoma. Front. Oncol. 2018, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Patel, S.A.; Velingkaar, N.; Makwana, K.; Chaudhari, A.; Kondratov, R. Calorie Restriction Regulates Circadian Clock Gene Expression through BMAL1 Dependent and Independent Mechanisms. Sci. Rep. 2016, 6, 25970. [Google Scholar] [CrossRef] [PubMed]
- Astafev, A.A.; Patel, S.A.; Kondratov, R.V. Calorie Restriction Effects on Circadian Rhythms in Gene Expression Are Sex Dependent. Sci. Rep. 2017, 7. [Google Scholar] [CrossRef] [Green Version]
- Hojo, H.; Enya, S.; Arai, M.; Suzuki, Y.; Nojiri, T.; Kangawa, K.; Koyama, S.; Kawaoka, S. Remote Reprogramming of Hepatic Circadian Transcriptome by Breast Cancer. Oncotarget 2017, 8, 34128–34140. [Google Scholar] [CrossRef] [Green Version]
- Kiessling, S.; Beaulieu-Laroche, L.; Blum, I.D.; Landgraf, D.; Welsh, D.K.; Storch, K.F.; Labrecque, N.; Cermakian, N. Enhancing Circadian Clock Function in Cancer Cells Inhibits Tumor Growth. BMC Biol. 2017, 15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iurisci, I.; Filipski, E.; Reinhardt, J.; Bach, S.; Gianella-Borradori, A.; Iacobelli, S.; Meijer, L.; Lévi, F. Improved Tumor Control through Circadian Clock Induction by Seliciclib, a Cyclin-Dependent Kinase Inhibitor. Cancer Res. 2006, 66, 10720–10728. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, X.M.; Delaunay, F.; Dulong, S.; Claustrat, B.; Zampera, S.; Fujii, Y.; Teboul, M.; Beau, J.; Lévi, F. Cancer Inhibition through Circadian Reprogramming of Tumor Transcriptome with Meal Timing. Cancer Res. 2010, 70, 3351–3360. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huisman, S.A.; Oklejewicz, M.; Ahmadi, A.R.; Tamanini, F.; Ijzermans, J.N.M.; Van Der Horst, G.T.J.; De Bruin, R.W.F. Colorectal Liver Metastases with a Disrupted Circadian Rhythm Phase Shift the Peripheral Clock in Liver and Kidney. Int. J. Cancer 2015, 136, 1027–1032. [Google Scholar] [CrossRef] [PubMed]
- Masri, S.; Papagiannakopoulos, T.; Kinouchi, K.; Liu, Y.; Cervantes, M.; Baldi, P.; Jacks, T.; Sassone-Corsi, P. Lung Adenocarcinoma Distally Rewires Hepatic Circadian Homeostasis. Cell 2016, 165, 896–909. [Google Scholar] [CrossRef] [Green Version]
- Koike, N.; Yoo, S.H.; Huang, H.C.; Kumar, V.; Lee, C.; Kim, T.K.; Takahashi, J.S. Transcriptional Architecture and Chromatin Landscape of the Core Circadian Clock in Mammals. Science 2012, 338, 349–354. [Google Scholar] [CrossRef] [Green Version]
- Patel, S.A.; Velingkaar, N.S.; Kondratov, R.V. Transcriptional Control of Antioxidant Defense by the Circadian Clock. Antioxid. Redox Signal. 2014, 20, 2997–3006. [Google Scholar] [CrossRef]
- Antoch, M.P.; Gorbacheva, V.Y.; Vykhovanets, O.; Toshkov, I.A.; Kondratov, R.V.; Kondratova, A.A.; Lee, C.; Nikitin, A.Y. Disruption of the Circadian Clock Due to the Clock Mutation Has Discrete Effects on Aging and Carcinogenesis. Cell Cycle 2008, 7, 1197–1204. [Google Scholar] [CrossRef] [Green Version]
- Gauger, M.A.; Sancar, A. Cryptochrome, Circadian Cycle, Cell Cycle Checkpoints, and Cancer. Cancer Res. 2005, 65, 6828–6834. [Google Scholar] [CrossRef] [Green Version]
- Huber, A.L.; Papp, S.J.; Chan, A.B.; Henriksson, E.; Jordan, S.D.; Kriebs, A.; Nguyen, M.; Wallace, M.; Li, Z.; Metallo, C.M.; et al. CRY2 and FBXL3 Cooperatively Degrade C-MYC. Mol. Cell 2016, 64, 774–789. [Google Scholar] [CrossRef] [Green Version]
- Puram, R.V.; Kowalczyk, M.S.; De Boer, C.G.; Schneider, R.K.; Miller, P.G.; McConkey, M.; Tothova, Z.; Tejero, H.; Heckl, D.; Järås, M.; et al. Core Circadian Clock Genes Regulate Leukemia Stem Cells in AML. Cell 2016, 165, 303–316. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ozturk, N.; Lee, J.H.; Gaddameedhi, S.; Sancar, A. Loss of Cryptochrome Reduces Cancer Risk in P53 Mutant Mice. Proc. Natl. Acad. Sci. USA 2009, 106, 2841–2846. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sulli, G.; Lam, M.T.Y.; Panda, S. Interplay between Circadian Clock and Cancer: New Frontiers for Cancer Treatment. Trends Cancer 2019, 5, 475–494. [Google Scholar] [CrossRef] [PubMed]
- Asher, G.; Schibler, U. Crosstalk between Components of Circadian and Metabolic Cycles in Mammals. Cell Metab. 2011, 13, 125–137. [Google Scholar] [CrossRef] [Green Version]
- Reinke, H.; Asher, G. Crosstalk between Metabolism and Circadian Clocks. Nat. Rev. Mol. Cell Biol. 2019, 20, 227–241. [Google Scholar] [CrossRef]
- Sinturel, F.; Petrenko, V.; Dibner, C. Circadian Clocks Make Metabolism Run. J. Mol. Biol. 2020, 432, 3680–3699. [Google Scholar] [CrossRef]
- Marcheva, B.; Ramsey, K.M.; Peek, C.B.; Affinati, A.; Maury, E.; Bass, J. Circadian Clocks and Metabolism. Handb. Exp. Pharmacol. 2013, 217, 127–155. [Google Scholar] [CrossRef]
- Kinouchi, K.; Sassone-Corsi, P. Metabolic Rivalry: Circadian Homeostasis and Tumorigenesis. Nat. Rev. Cancer 2020, 20, 645–661. [Google Scholar] [CrossRef] [PubMed]
- Verlande, A.; Masri, S. Circadian Clocks and Cancer: Timekeeping Governs Cellular Metabolism. Trends Endocrinol. Metab. 2019, 30, 445–458. [Google Scholar] [CrossRef]
- Altman, B.J. Cancer Clocks out for Lunch: Disruption of Circadian Rhythm and Metabolic Oscillation in Cancer. Front. Cell Dev. Biol. 2016, 4. [Google Scholar] [CrossRef] [Green Version]
- Tian, T.; Li, X.; Zhang, J. MTOR Signaling in Cancer and Mtor Inhibitors in Solid Tumor Targeting Therapy. Int. J. Mol. Sci. 2019, 20, 755. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Porta, C.; Paglino, C.; Mosca, A. Targeting PI3K/Akt/MTOR Signaling in Cancer. Front. Oncol. 2014, 4. [Google Scholar] [CrossRef] [Green Version]
- Khapre, R.V.; Kondratova, A.A.; Patel, S.; Dubrovsky, Y.; Wrobel, M.; Antoch, M.P.; Kondratov, R.V. BMAL1-Dependent Regulation of the MTOR Signaling Pathway Delays Aging. Aging 2014, 6, 48–57. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khapre, R.V.; Patel, S.A.; Kondratova, A.A.; Chaudhary, A.; Velingkaar, N.; Antoch, M.P.; Kondratov, R.V. Metabolic Clock Generates Nutrient Anticipation Rhythms in MTOR Signaling. Aging 2014, 6, 675–689. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cao, R. MTOR Signaling, Translational Control, and the Circadian Clock. Front. Genet. 2018, 9. [Google Scholar] [CrossRef] [PubMed]
- Cao, R.; Robinson, B.; Xu, H.; Gkogkas, C.; Khoutorsky, A.; Alain, T.; Yanagiya, A.; Nevarko, T.; Liu, A.C.; Amir, S.; et al. Translational Control of Entrainment and Synchrony of the Suprachiasmatic Circadian Clock by MTOR/4E-BP1 Signaling. Neuron 2013, 79, 712–724. [Google Scholar] [CrossRef] [Green Version]
- Patel, S.A.; Chaudhari, A.; Gupta, R.; Velingkaar, N.; Kondratov, R.V. Circadian Clocks Govern Calorie Restriction-Mediated Life Span Extension through BMAL1-and IGF-1-Dependent Mechanisms. FASEB J. 2015, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Tulsian, R.; Velingkaar, N.; Kondratov, R. Caloric Restriction Effects on Liver MTOR Signaling Are Time-of-Day Dependent. Aging 2018, 10, 1640–1648. [Google Scholar] [CrossRef] [PubMed]
- Velingkaar, N.; Mezhnina, V.; Poe, A.; Makwana, K.; Tulsian, R.; Kondratov, R.V. Reduced Caloric Intake and Periodic Fasting Independently Contribute to Metabolic Effects of Caloric Restriction. Aging Cell 2020, 19. [Google Scholar] [CrossRef]
- Yoon, M.S. The Role of Mammalian Target of Rapamycin (MTOR) in Insulin Signaling. Nutrients 2017, 9, 1176. [Google Scholar] [CrossRef]
- Vander Haar, E.; Lee, S.I.; Bandhakavi, S.; Griffin, T.J.; Kim, D.H. Insulin Signalling to MTOR Mediated by the Akt/PKB Substrate PRAS. Nat. Cell Biol. 2007, 9, 316–323. [Google Scholar] [CrossRef]
- Marcheva, B.; Ramsey, K.M.; Buhr, E.D.; Kobayashi, Y.; Su, H.; Ko, C.H.; Ivanova, G.; Omura, C.; Mo, S.; Vitaterna, M.H.; et al. Disruption of the Clock Components CLOCK and BMAL1 Leads to Hypoinsulinaemia and Diabetes. Nature 2010, 466, 627–631. [Google Scholar] [CrossRef] [Green Version]
- Chaudhari, A.; Gupta, R.; Patel, S.; Velingkaar, N.; Kondratov, R. Cryptochromes Regulate IGF-1 Production and Signaling through Control of JAK2-Dependent STAT5B Phosphorylation. Mol. Biol. Cell 2017, 28, 834–842. [Google Scholar] [CrossRef]
- Wu, R.; Dang, F.; Li, P.; Wang, P.; Xu, Q.; Liu, Z.; Li, Y.; Wu, Y.; Chen, Y.; Liu, Y. The Circadian Protein Period2 Suppresses MTORC1 Activity via Recruiting Tsc1 to MTORC1 Complex. Cell Metab. 2019, 29, 653–667.e6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lipton, J.O.; Yuan, E.D.; Boyle, L.M.; Ebrahimi-Fakhari, D.; Kwiatkowski, E.; Nathan, A.; Güttler, T.; Davis, F.; Asara, J.M.; Sahin, M. The Circadian Protein BMAL1 Regulates Translation in Response to S6K1-Mediated Phosphorylation. Cell 2015, 161, 1138–1151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lipton, J.O.; Boyle, L.M.; Yuan, E.D.; Hochstrasser, K.J.; Chifamba, F.F.; Nathan, A.; Tsai, P.T.; Davis, F.; Sahin, M. Aberrant Proteostasis of BMAL1 Underlies Circadian Abnormalities in a Paradigmatic MTOR-Opathy. Cell Rep. 2017, 20, 868–880. [Google Scholar] [CrossRef] [Green Version]
- Makwana, K.; Gosai, N.; Poe, A.; Kondratov, R.V. Calorie Restriction Reprograms Diurnal Rhythms in Protein Translation to Regulate Metabolism. FASEB J. 2019, 33, 4473–4489. [Google Scholar] [CrossRef]
- Liu, D.; Stowie, A.; De Zavalia, N.; Leise, T.; Pathak, S.S.; Drewes, L.R.; Davidson, A.J.; Amir, S.; Sonenberg, N.; Cao, R. MTOR Signaling in VIP Neurons Regulates Circadian Clock Synchrony and Olfaction. Proc. Natl. Acad. Sci. USA. 2018, 115, E3296–E3304. [Google Scholar] [CrossRef] [Green Version]
- Cao, R.; Li, A.; Cho, H.Y.; Lee, B.; Obrietan, K. Mammalian Target of Rapamycin Signaling Modulates Photic Entrainment of the Suprachiasmatic Circadian Clock. J. Neurosci. 2010, 30, 6302–6314. [Google Scholar] [CrossRef]
- Ramanathan, C.; Kathale, N.D.; Liu, D.; Lee, C.; Freeman, D.A.; Hogenesch, J.B.; Cao, R.; Liu, A.C. MTOR Signaling Regulates Central and Peripheral Circadian Clock Function. PLoS Genet. 2018, 14. [Google Scholar] [CrossRef] [PubMed]
- Zhang, E.E.; Zhang, E.E.; Liu, A.C.; Liu, A.C.; Hirota, T.; Hirota, T.; Miraglia, L.J.; Miraglia, L.J.; Welch, G.; Welch, G.; et al. A Genome-Wide RNAi Screen for Modifiers of the Circadian Clock in Human Cells. Cell 2009, 139, 199–210. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Okazaki, H.; Matsunaga, N.; Fujioka, T.; Okazaki, F.; Akagawa, Y.; Tsurudome, Y.; Ono, M.; Kuwano, M.; Koyanagi, S.; Ohdo, S. Circadian Regulation of MTOR by the Ubiquitin Pathway in Renal Cell Carcinoma. Cancer Res. 2014, 74, 543–551. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Devocelle, A.; Souza, L.; Foudi, A.; Bento, S.T.; Desterke, C.; Sherrard, R.; Ballesta, A.; Adam, R.; Giron-Michel, J.; et al. BMAL1 Knockdown Triggers Different Colon Carcinoma Cell Fates by Altering the Delicate Equilibrium between AKT/MTOR and P53/P21 Pathways. Aging 2020, 12, 8067–8083. [Google Scholar] [CrossRef] [PubMed]
- Chen, B.; Tan, Y.; Li, Y.; Chen, L.; Wu, S.; Xu, W.; Wang, Y.; Zhao, W.; Wu, J. Per2 Participates in AKT-Mediated Drug Resistance in A549/DDP Lung Adenocarcinoma Cells. Oncol. Lett. 2017, 13, 423–428. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, H.; Gong, X.; Yang, K. Overexpression of the Clock Gene Per2 Suppresses Oral Squamous Cell Carcinoma Progression by Activating Autophagy via the PI3K/AKT/MTOR Pathway. J. Cancer 2020, 11, 3655–3666. [Google Scholar] [CrossRef] [Green Version]
- Yang, G.; Yang, Y.; Tang, H.; Yang, K. Loss of the Clock Gene Per1 Promotes Oral Squamous Cell Carcinoma Progression via the AKT/MTOR Pathway. Cancer Sci. 2020, 111, 1542–1554. [Google Scholar] [CrossRef]
- Matsumoto, C.S.; Almeida, L.O.; Guimarães, D.M.; Martins, M.D.; Papagerakis, P.; Papagerakis, S.; Leopoldino, A.M.; Castilho, R.M.; Squarize, C.H. PI3K-PTEN Dysregulation Leads to MTOR-Driven Upregulation of the Core Clock Gene BMAL1 in Normal and Malignant Epithelial Cells. Oncotarget 2016, 7, 42393–42407. [Google Scholar] [CrossRef] [Green Version]
- Chan, S.; Zhang, L.; Rowbottom, L.; McDonald, R.; Bjarnason, G.A.; Tsao, M.; Barnes, E.; Danjoux, C.; Popovic, M.; Lam, H.; et al. Effects of Circadian Rhythms and Treatment Times on the Response of Radiotherapy for Painful Bone Metastases. Ann. Palliat. Med. 2017, 6, 14–25. [Google Scholar] [CrossRef]
- Ballesta, A.; Innominato, P.F.; Dallmann, R.; Rand, D.A.; Lévi, F.A. Systems Chronotherapeutics. Pharmacol. Rev. 2017, 69, 161–199. [Google Scholar] [CrossRef] [Green Version]
- Dallmann, R.; Okyar, A.; Lévi, F. Dosing-Time Makes the Poison: Circadian Regulation and Pharmacotherapy. Trends Mol. Med. 2016, 22, 430–445. [Google Scholar] [CrossRef] [Green Version]
- Shuboni-Mulligan, D.D.; Breton, G.; Smart, D.D.; Gilbert, M.; Armstrong, T.S. Radiation Chronotherapy—Clinical Impact of Treatment Time-of-Day: A Systematic Review. J. Neurooncol. 2019, 145, 415–427. [Google Scholar] [CrossRef]
- Coudert, B.; Focan, C.; Genet, D.; Giacchetti, S.; Cvickovic, F.; Zambelli, A.; Fillet, G.; Chollet, P.; Amoroso, D.; Van Der Auwera, J.; et al. A Randomized Multicenter Study of Optimal Circadian Time of Vinorelbine Combined with Chronomodulated 5-Fluorouracil in Pretreated Metastatic Breast Cancer Patients: EORTC Trial. Chronobiol. Int. 2008, 25, 680–696. [Google Scholar] [CrossRef] [PubMed]
- Gallion, H.H.; Brunetto, V.L.; Cibull, M.; Lentz, S.S.; Reid, G.; Soper, J.T.; Burger, R.A.; Andersen, W.; Mackey, D. Randomized Phase III Trial of Standard Timed Doxorubicin plus Cisplatin versus Circadian Timed Doxorubicin plus Cisplatin in Stage III and IV or Recurrent Endometrial Carcinoma: A Gynecologic Oncology Group Study. J. Clin. Oncol. 2003, 21, 3808–3813. [Google Scholar] [CrossRef]
- Kobayashi, M.; Wood, P.A.; Hrushesky, W.J.M. Circadian Chemotherapy for Gynecological and Genitourinary Cancers. Chronobiol. Int. 2002, 19, 237–251. [Google Scholar] [CrossRef] [PubMed]
- Lévi, F.A.; Zidani, R.; Vannetzel, J.M.; Perpoint, B.; Focan, C.; Faggiuolo, R.; Chollet, P.; Garufi, C.; Itzhaki, M.; Dogliotti, L.; et al. Chronomodulated versus Fixed-Infusion-Rate Delivery of Ambulatory Chemotherapy with Oxaliplatin, Fluorouracil, and Folinic Acid (Leucovorin) in Patients with Colorectal Cancer Metastases: A Randomized Multi-Institutional Trial. J. Natl. Cancer Inst. 1994, 86, 1608–1617. [Google Scholar] [CrossRef] [PubMed]
- Lévi, F.; Zidani, R.; Misset, J.L. Randomised Multicentre Trial of Chronotherapy with Oxaliplatin, Fluorouracil, and Folinic Acid in Metastatic Colorectal Cancer. Lancet 1997, 350, 681–686. [Google Scholar] [CrossRef]
- Giacchetti, S. Chronotherapy of Colorectal Cancer. Chronobiol. Int. 2002, 19, 207–219. [Google Scholar] [CrossRef] [PubMed]
- Bouchahda, M.; Adam, R.; Giacchetti, S.; Castaing, D.; Brezault-Bonnet, C.; Hauteville, D.; Innominato, P.F.; Focan, C.; Machover, D.; Lévi, F. Rescue Chemotherapy Using Multidrug Chronomodulated Hepatic Arterial Infusion for Patients with Heavily Pretreated Metastatic Colorectal Cancer. Cancer 2009, 115, 4990–4999. [Google Scholar] [CrossRef]
- Gorbacheva, V.Y.; Kondratov, R.V.; Zhang, R.; Cherukuri, S.; Gudkov, A.V.; Takahashi, J.S.; Antoch, M.P. Circadian Sensitivity to the Chemotherapeutic Agent Cyclophosphamide Depends on the Functional Status of the CLOCK/BMAL1 Transactivation Complex. Proc. Natl. Acad. Sci. USA 2005, 10, 3407–3412. [Google Scholar] [CrossRef] [Green Version]
- Vitaterna, M.H.; Selby, C.P.; Todo, T.; Niwa, H.; Thompson, C.; Fruechte, E.M.; Hitomi, K.; Thresher, R.J.; Ishikawa, T.; Miyazaki, J.; et al. Differential Regulation of Mammalian Period Genes and Circadian Rhythmicity by Cryptochromes 1 and 2. Proc. Natl. Acad. Sci. USA 1999, 96, 12114–12119. [Google Scholar] [CrossRef] [Green Version]
- Sulli, G.; Manoogian, E.N.C.; Taub, P.R.; Panda, S. Training the Circadian Clock, Clocking the Drugs, and Drugging the Clock to Prevent, Manage, and Treat Chronic Diseases. Trends Pharmacol. Sci. 2018, 39, 812–827. [Google Scholar] [CrossRef]
- Lee, Y.; Lahens, N.F.; Zhang, S.; Bedont, J.; Field, J.M.; Sehgal, A. G1/S Cell Cycle Regulators Mediate Effects of Circadian Dysregulation on Tumor Growth and Provide Targets for Timed Anticancer Treatment. PLoS Biol. 2019, 17. [Google Scholar] [CrossRef] [Green Version]
- Ozturk, N.; Ozturk, D.; Pala-Kara, Z.; Kaptan, E.; Sancar-Bas, S.; Ozsoy, N.; Cinar, S.; Deniz, G.; Li, X.M.; Giacchetti, S.; et al. The Immune System as a Chronotoxicity Target of the Anticancer MTOR Inhibitor Everolimus. Chronobiol. Int. 2018, 35, 705–718. [Google Scholar] [CrossRef]
- Puzanov, I.; Diab, A.; Abdallah, K.; Bingham, C.O.; Brogdon, C.; Dadu, R.; Hamad, L.; Kim, S.; Lacouture, M.E.; LeBoeuf, N.R.; et al. Managing Toxicities Associated with Immune Checkpoint Inhibitors: Consensus Recommendations from the Society for Immunotherapy of Cancer (SITC) Toxicity Management Working Group. J. Immunother. Cancer 2017, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deng, W.; Zhu, S.; Zeng, L.; Liu, J.; Kang, R.; Yang, M.; Cao, L.; Wang, H.; Billiar, T.R.; Jiang, J.; et al. The Circadian Clock Controls Immune Checkpoint Pathway in Sepsis. Cell Rep. 2018, 24, 366–378. [Google Scholar] [CrossRef] [Green Version]
- Guerrero-Vargas, N.N.; Salgado-Delgado, R.; Basualdo, M.C.; García, J.; Guzmán-Ruiz, M.; Carrero, J.C.; Escobar, C.; Buijs, R.M. Reciprocal Interaction between the Suprachiasmatic Nucleus and the Immune System Tunes down the Inflammatory Response to Lipopolysaccharide. J. Neuroimmunol. 2014, 273, 22–30. [Google Scholar] [CrossRef]
- Liu, J.; Malkani, G.; Shi, X.; Meyer, M.; Cunningham-Runddles, S.; Ma, X.; Zhong, S.S. Erratum: The Circadian Clock Period 2 Gene Regulates Gamma Interferon Production of NK Cells in Host Response to Lipopolysaccharide-Induced Endotoxic Shock. Infect. Immun. 2006, 74, 4750–4756. [Google Scholar] [CrossRef] [Green Version]
- Marinac, C.R.; Natarajan, L.; Sears, D.D.; Gallo, L.C.; Hartman, S.J.; Arredondo, E.; Patterson, R.E. Prolonged Nightly Fasting and Breast Cancer Risk: Findings from NHANES (2009–2010). Cancer Epidemiol. Biomarkers Prev. 2015, 24, 783–789. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marinac, C.R.; Sears, D.D.; Natarajan, L.; Gallo, L.C.; Breen, C.I.; Patterson, R.E. Frequency and Circadian Timing of Eating May Influence Biomarkers of Inflammation and Insulin Resistance Associated with Breast Cancer Risk. PLoS ONE 2015, 10. [Google Scholar] [CrossRef] [Green Version]
- Wu, M.W.; Li, X.M.; Xian, L.J.; Lévi, F. Effects of Meal Timing on Tumor Progression in Mice. Life Sci. 2004, 75, 1181–1193. [Google Scholar] [CrossRef]
- Filipski, E.; Lévi, F. Circadian Disruption in Experimental Cancer Processes. Integr. Cancer Ther. 2009, 8, 298–302. [Google Scholar] [CrossRef] [Green Version]
- James, P.; Bertrand, K.A.; Hart, J.E.; Schernhammer, E.S.; Tamimi, R.M.; Laden, F. Outdoor Light at Night and Breast Cancer Incidence in the Nurses’ Health Study II. Environ. Health Perspect. 2017, 125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johns, L.E.; Jones, M.E.; Schoemaker, M.J.; McFadden, E.; Ashworth, A.; Swerdlow, A.J. Domestic Light at Night and Breast Cancer Risk: A Prospective Analysis of 105 000 UK Women in the Generations Study. Br. J. Cancer 2018, 118, 600–606. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, Y.J.; Park, M.S.; Lee, E.; Choi, J.W. High Incidence of Breast Cancer in Light-Polluted Areas with Spatial Effects in Korea. Asian Pac. J. Cancer Prev. 2016, 17, 361–367. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kloog, I.; Haim, A.; Stevens, R.G.; Barchana, M.; Portnov, B.A. Light at Night Co-Distributes with Incident Breast but Not Lung Cancer in the Female Population of Israel. Chronobiol. Int. 2008, 25, 65–81. [Google Scholar] [CrossRef] [PubMed]
- Ritonja, J.; McIsaac, M.A.; Sanders, E.; Kyba, C.C.M.; Grundy, A.; Cordina-Duverger, E.; Spinelli, J.J.; Aronson, K.J. Outdoor Light at Night at Residences and Breast Cancer Risk in Canada. Eur. J. Epidemiol. 2020, 35, 579–589. [Google Scholar] [CrossRef] [PubMed]
- Stevens, R.G. Light-at-Night, Circadian Disruption and Breast Cancer: Assessment of Existing Evidence. Int. J. Epidemiol. 2009, 38, 963–970. [Google Scholar] [CrossRef]
- Wu, J.; Dauchy, R.T.; Tirrell, P.C.; Wu, S.S.; Lynch, D.T.; Jitawatanarat, P.; Burrington, C.M.; Dauchy, E.M.; Blask, D.E.; Greene, M.W. Light at Night Activates IGF-1R/PDK1 Signaling and Accelerates Tumor Growth in Human Breast Cancer Xenografts. Cancer Res. 2011, 71, 2622–2631. [Google Scholar] [CrossRef] [Green Version]
- Blask, D.E.; Dauchy, R.T.; Sauer, L.A.; Krause, J.A.; Brainard, G.C. Growth and Fatty Acid Metabolism of Human Breast Cancer (MCF-7) Xenografts in Nude Rats: Impact of Constant Light-Induced Nocturnal Melatonin Suppression. Breast Cancer Res. Treat. 2003, 79, 313–320. [Google Scholar] [CrossRef]
- Schwimmer, H.; Metzer, A.; Pilosof, Y.; Szyf, M.; Machnes, Z.M.; Fares, F.; Harel, O.; Haim, A. Light at Night and Melatonin Have Opposite Effects on Breast Cancer Tumors in Mice Assessed by Growth Rates and Global DNA Methylation. Chronobiol. Int. 2014, 31, 144–150. [Google Scholar] [CrossRef] [PubMed]
- Zubidat, A.E.; Fares, B.; Fares, F.; Haim, A. Artificial Light at Night of Different Spectral Compositions Differentially Affects Tumor Growth in Mice: Interaction With Melatonin and Epigenetic Pathways. Cancer Control. 2018, 25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zubidat, A.E. Melatonin Through DNA Methylation Constricts Breast Cancer Growth Accelerated By Blue LED Light-At-Night In 4T1 Tumor-Bearing Mice. J. Cancer Biol. Ther. 2015, 1. [Google Scholar] [CrossRef]
- Jung-Hynes, B.; Huang, W.; Reiter, R.J.; Ahmad, N. Melatonin Resynchronizes Dysregulated Circadian Rhythm Circuitry in Human Prostate Cancer Cells. J. Pineal Res. 2010, 49, 60–68. [Google Scholar] [CrossRef] [Green Version]
- Savvidis, C.; Koutsilieris, M. Circadian Rhythm Disruption in Cancer Biology. Mol. Med. 2012, 18, 1249–1260. [Google Scholar] [CrossRef]
- Grubisic, M.; Haim, A.; Bhusal, P.; Dominoni, D.M.; Gabriel, K.M.A.; Jechow, A.; Kupprat, F.; Lerner, A.; Marchant, P.; Riley, W.; et al. Light Pollution, Circadian Photoreception, and Melatonin in Vertebrates. Sustainability 2019, 11, 6400. [Google Scholar] [CrossRef] [Green Version]
- Baturin, D.A.; Alimova, I.N.; Anisimov, V.N.; Popovich, I.G.; Zabezhinski, M.A.; Provinciali, M.; Mancini, R.; Franceschi, C. The Effect of Light Regimen and Melatonin on the Development of Spontaneous Mammary Tumors in HER-2/Neu Transgenic Mice Is Related to a Downregulation of HER-2/Neu Gene Expression. Neuroendocrinol. Lett. 2001, 22, 441–447. [Google Scholar]
- Anisimov, V.N.; Alimova, I.N.; Baturin, D.A.; Popovich, I.G.; Zabezhinski, M.A.; Manton, K.G.; Semenchenko, A.V.; Yashin, A.I. The Effect of Melatonin Treatment Regimen on Mammary Adenocarcinoma Development in HER-2/Neu Transgenic Mice. Int. J. Cancer 2003, 103, 300–305. [Google Scholar] [CrossRef]
- Haus, E.L.; Smolensky, M.H. Shift Work and Cancer Risk: Potential Mechanistic Roles of Circadian Disruption, Light at Night, and Sleep Deprivation. Sleep Med. Rev. 2013, 17, 273–284. [Google Scholar] [CrossRef] [PubMed]
- Oshima, T.; Niwa, Y.; Kuwata, K.; Srivastava, A.; Hyoda, T.; Tsuchiya, Y.; Kumagai, M.; Tsuyuguchi, M.; Tamaru, T.; Sugiyama, A.; et al. Cell-Based Screen Identifies a New Potent and Highly Selective CK2 Inhibitor for Modulation of Circadian Rhythms and Cancer Cell Growth. Sci. Adv. 2019, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, J.; Azmi, A.S.; Aboukameel, A.; Kauffman, M.; Shacham, S.; Abou-Samra, A.B.; Mohammad, R.M. Nuclear Retention of Fbw7 by Specific Inhibitors of Nuclear Export Leads to Notch1 Degradation in Pancreatic Cancer. Oncotarget 2014, 5, 3444–3454. [Google Scholar] [CrossRef] [Green Version]
- Sulli, G.; Rommel, A.; Wang, X.; Kolar, M.J.; Puca, F.; Saghatelian, A.; Plikus, M.V.; Verma, I.M.; Panda, S. Pharmacological Activation of REV-ERBs Is Lethal in Cancer and Oncogene-Induced Senescence. Nature 2018, 553, 351–355. [Google Scholar] [CrossRef] [PubMed]
- Hu, X.; Liu, X.; Moisan, J.; Wang, Y.; Lesch, C.A.; Spooner, C.; Morgan, R.W.; Zawidzka, E.M.; Mertz, D.; Bousley, D.; et al. Synthetic RORγ Agonists Regulate Multiple Pathways to Enhance Antitumor Immunity. Oncoimmunology 2016, 5. [Google Scholar] [CrossRef] [PubMed]
- Lellupitiyage Don, S.S.; Robertson, K.L.; Lin, H.H.; Labriola, C.; Harrington, M.E.; Taylor, S.R.; Farkas, M.E. Nobiletin Affects Circadian Rhythms and Oncogenic Characteristics in a Cell-Dependent Manner. PLoS ONE 2020, 15. [Google Scholar] [CrossRef]
- He, B.; Nohara, K.; Park, N.; Park, Y.S.; Guillory, B.; Zhao, Z.; Garcia, J.M.; Koike, N.; Lee, C.C.; Takahashi, J.S.; et al. The Small Molecule Nobiletin Targets the Molecular Oscillator to Enhance Circadian Rhythms and Protect against Metabolic Syndrome. Cell Metab. 2016, 23, 610–621. [Google Scholar] [CrossRef] [Green Version]
- Gaddameedhi, S.; Selby, C.P.; Kaufmann, W.K.; Smart, R.C.; Sancar, A. Control of Skin Cancer by the Circadian Rhythm. Proc. Natl. Acad. Sci. USA 2011, 108, 18790–18795. [Google Scholar] [CrossRef] [Green Version]
- Kuo, T.T.; Ladurner, A.G. Exploiting the Circadian Clock for Improved Cancer Therapy: Perspective from a Cell Biologist. Front. Genet. 2019, 10. [Google Scholar] [CrossRef]
- Hill, R.J.W.; Innominato, P.F.; Lévi, F.; Ballesta, A. Optimizing Circadian Drug Infusion Schedules towards Personalized Cancer Chronotherapy. PLoS Comput. Biol. 2020, 16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, X.M.; Mohammad-Djafari, A.; Dumitru, M.; Dulong, S.; Filipski, E.; Siffroi-Fernandez, S.; Mteyrek, A.; Scaglione, F.; Guettier, C.; Delaunay, F.; et al. A Circadian Clock Transcription Model for the Personalization of Cancer Chronotherapy. Cancer Res. 2013, 73, 7176–7188. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mormont, M.C.; Waterhouse, J. Contribution of the Rest-Activity Circadian Rhythm to Quality of Life in Cancer Patients. Chronobiol. Int. 2002, 19, 313–323. [Google Scholar] [CrossRef]
- Focan, C.; Gorlia, T.; Moreau, T.; Garufi, C.; Waterhouse, J.; Giacchetti, S.; Coudert, B.; Iacobelli, S.; Genet, D.; Tampellini, M.; et al. Circadian Rhythm in Rest and Activity: A Biological Correlate of Quality of Life and a Predictor of Survival in Patients with Metastatic Colorectal Cancer. Cancer Res. 2009, 69, 4700–4707. [Google Scholar]
- Innominato, P.F.; Giacchetti, S.; Bjarnason, G.A.; Focan, C.; Garufi, C.; Coudert, B.; Iacobelli, S.; Tampellini, M.; Durando, X.; Mormont, M.C.; et al. Prediction of Overall Survival through Circadian Rest-Activity Monitoring during Chemotherapy for Metastatic Colorectal Cancer. Int. J. Cancer 2012, 131, 2684–2692. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Clock Genes | Animal Cancer Model | Outcome | References |
|---|---|---|---|
| Rev Erbα, Rorγ and Per2 | 4T1 Breast Cancer | Downregulation | [54] |
| Per1, Per2 and Bmal1 | B16F10 Melanoma | Downregulation | [55] |
| Rev Erbα, Per2 and Bmal1 | Glasgow osteosarcoma | Downregulation | [56] |
| Rev Erbα, Per2 and Bmal1 | P03 Pancreatic adenocarcinoma | Downregulation | [57] |
| Rev-Erbα, Per1, Per2, and Bmal1 | C26 Colorectal liver metastases | Downregulation | [58] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Patel, S.A.; Kondratov, R.V. Clock at the Core of Cancer Development. Biology 2021, 10, 150. https://doi.org/10.3390/biology10020150
Patel SA, Kondratov RV. Clock at the Core of Cancer Development. Biology. 2021; 10(2):150. https://doi.org/10.3390/biology10020150
Chicago/Turabian StylePatel, Sonal A., and Roman V. Kondratov. 2021. "Clock at the Core of Cancer Development" Biology 10, no. 2: 150. https://doi.org/10.3390/biology10020150
APA StylePatel, S. A., & Kondratov, R. V. (2021). Clock at the Core of Cancer Development. Biology, 10(2), 150. https://doi.org/10.3390/biology10020150