
Unpredictable, Counter-Intuitive Geoclimatic and Demographic Correlations of COVID-19 Spread Rates



Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
3. Results
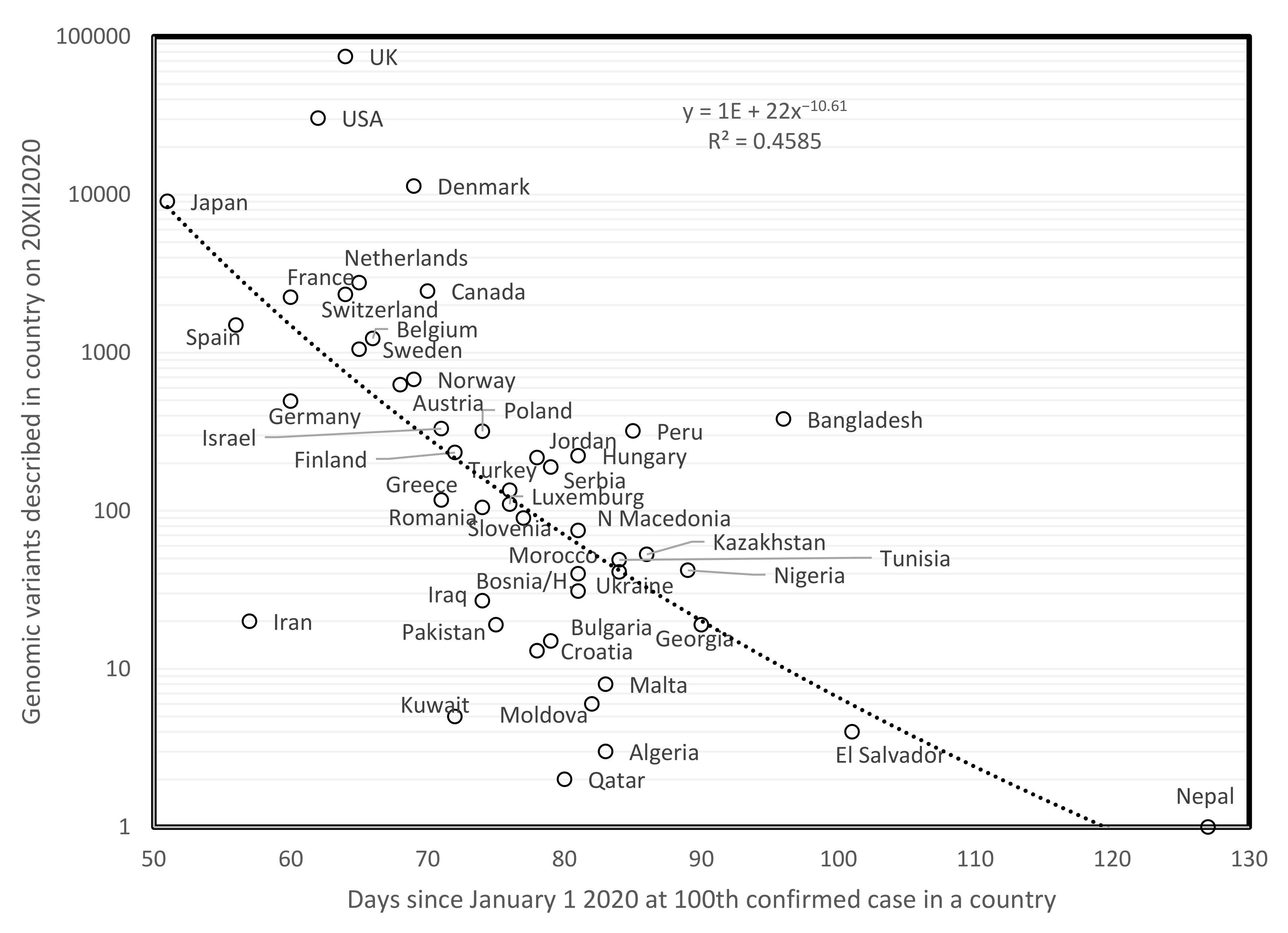
3.1. Mutations Cumulate over Time during the COVID-19 Outbreak
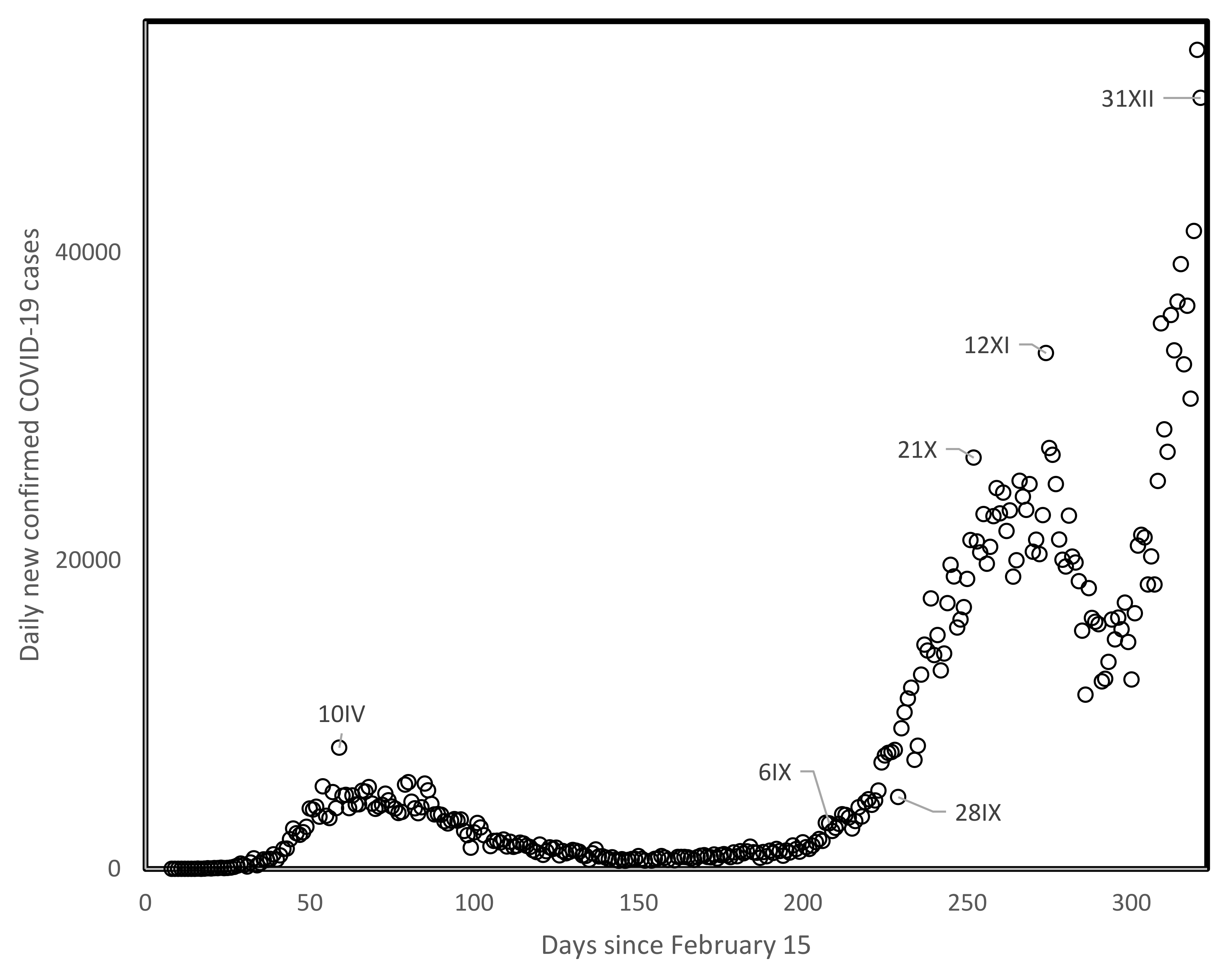
3.2. Spread Parameters for First and Second Waves in the USA
3.3. Visual Inspection vs. Objective Statistical Analysis
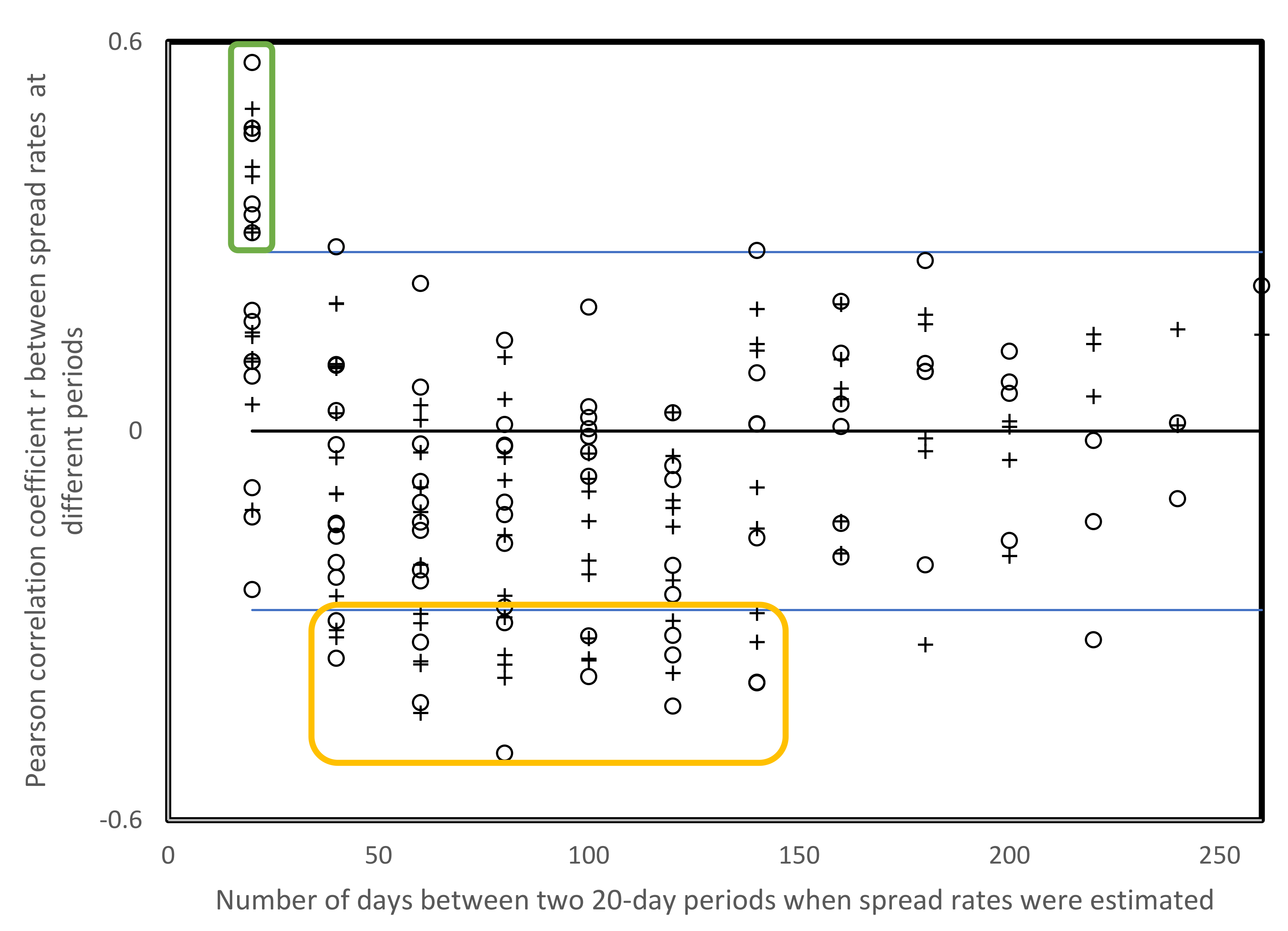
3.4. Spread Rates for Consecutive Nonoverlapping 20-Day Windows
3.5. Environmental Correlates of Spread Rates across Time
4. Discussion
4.1. A Potential Epidemiological Explanation for Spread Rate Inversions
4.2. Negative Correlations between Spread Rates Are Not between Ascending and Descending Parts of the Same Wave
4.3. Does Confinement Increase Ulterior Spread Rates?
4.4. Intrinsic vs. Extrinsic Constraints
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Demongeot, J.; Flet-Berliac, Y.; Seligmann, H. Temperature decreases spread parameters of the new covid-19 cases dynamics. Biology 2020, 9, 94. [Google Scholar] [CrossRef] [PubMed]
- Seligmann, H.; Iggui, S.; Rachdi, M.; Vuillerme, N.; Demongeot, J. Inverted covariate effects for first vs mutated second wave Covid-19: High temperature spread biased for young. Biology 2020, 9, 226. [Google Scholar] [CrossRef] [PubMed]
- Malki, Z.; Atlam, E.-S.; Hassanien, A.E.; Dagnew, G.; Elhosseini, M.A.; Gad, I. Association between weather data and COVID-19 pandemic predicting mortality rate: Machine learning approaches. Chaos Solution Fractals 2020, 138, 110137. [Google Scholar] [CrossRef]
- Forster, P.; Forster, L.; Renfrew, C.; Forster, M. Phylogenetic network analysis of SARS-CoV-2 genomes. Proc. Natl. Acad. Sci. USA 2020, 117, 9241–9243. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Esteban, D. Quasispecies dynamics in disease prevention and control. Virus Popul. 2020, 263–299. [Google Scholar]
- Liu, Z.; Magal, P.; Seydi, O.; Webb, G. Understanding Unreported Cases in the COVID-19 Epidemic Outbreak in Wuhan, China, and the Importance of Major Public Health Interventions. Biology 2020, 9, 50. [Google Scholar] [CrossRef] [Green Version]
- Lund, R.; Reeves, J. Detection of Undocumented Changepoints: A Revision of the Two-Phase Regression Model. J. Climate Sci. 2002, 15, 2547–2554. [Google Scholar] [CrossRef]
- Percival, D.B.; Rothrock, D.A. “Eyeballing” trends in climate time series: A cautionary tale. J. Climate Sci. 2005, 18, 886–891. [Google Scholar] [CrossRef] [Green Version]
- Ghorbanzadeh, D.; Picard, D. Etude asymptomatique et pratique du comportement de deux tests de détection de rupture. Statistique et Analyse de Données 1992, 16, 63–84. [Google Scholar]
- Ondo, J.C. Étude comparative des tests de stationnarité. Ph.D. Thesis, Québec University, Quebec City, QC, Canada, 2002; p. 288. [Google Scholar]
- Krishnan, N.M.; Seligmann, H.; Raina, S.D.; Pollock, D.D. Detecting gradients of asymmetry in site-specific substitutions in mitochondrial genomes. DNA Cell Biol. 2004, 23, 707–714. [Google Scholar] [CrossRef]
- Manzourolajdad, A.; Xu, Z.; Ebrahimi, D. Novel Polybasic Cleavage Site in SARS-CoV-2 Genome Is Likely to Induce a Major Change in the RNA Secondary Structure. Preprints 2020. [Google Scholar] [CrossRef]
- Janssen, G.M.; de Jong, G.; Joosse, E.N.G.; Scharloo, W. A negative maternal effect in spring tails. Evolution 1988, 42, 828–834. [Google Scholar]
- Haldane, J.B.S. The negative heritability of neonatal jaundice. Ann. Hum. Genet. 1996, 60, 3–5. [Google Scholar] [CrossRef] [PubMed]
- Steinsaltz, D.; Dahl, A.; Wachter, K.W. On negative heritability and negative estimates of heritability. Genetics 2020, 215, 343–357. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stam, E.; Isaaks, A.; Ernsting, G. Negative maternal effect revisited: A test on two populations of Orchesella cincta L. (Collembola: Entomobryidae). Evolution 2017, 52, 1839–1843. [Google Scholar] [CrossRef] [PubMed]
- Bonduriansky, R.; Crean, A.J.; Day, T. The implications of nongenetic inheritance for evolution in changing environments. Evol. App. 2011, 5, 192–201. [Google Scholar] [CrossRef]
- Seligmann, H.; Amzallag, G.N. Adaptive determinism during salt-adaptation in Sorghum bicolor. Biosystems 1995, 36, 71–77. [Google Scholar] [CrossRef]
- Seligmann, H. Transmission of acquired adjustments to salinity in Sorghum bicolor. Biosystems 1997, 40, 257–261. [Google Scholar] [CrossRef]
- Seligmann H 1998 Effect of environmental complexity on salt-adaptation in Sorghum bicolor. Biosystems 1998, 45, 213–220. [CrossRef]
- Saadeh, D.; Feeney, S.M.; Pontzen, A.; Peiris, H.V.; McEwen, J.D. How isotropic is the universe? Phys. Rev. Lett. 2016, 117. [Google Scholar] [CrossRef] [PubMed]
- Horn, H.S. Forest succession. Sci. Am. 1975, 232, 90–101. [Google Scholar] [CrossRef]
- Wardle, P. Primary succession in Westland National park and its vicinity. New Zealand J. Bot. 1980, 18, 221. [Google Scholar] [CrossRef]
- Reyes, H.A.; Ferreira, P.F.A.; Silva, L.C.; da Costa, M.G.; Nobre, C.P.; Gehring, C. Arbuscular mycorrhizal fungi along secondary forest succession at the eastern periphery of Amazonia: Seasonal variability and impacts of soil fertility. App. Soil Ecol. 2019, 136, 1–10. [Google Scholar] [CrossRef]
- Seligmann, H. Resource partition history and evolutionary specialization of subunits in complex systems. Biosystems 1999, 51, 31–39. [Google Scholar] [CrossRef]
- Hsiao, C.; Mohan, S.; Kalahar, B.K.; Williams, L.D. Peeling the onion: Ribosomes are ancient molecular fossils. Mol. Biol. Evol. 2009, 26, 2415–2425. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Petrov, A.S.; Gulen, B.; Norris, A.M.; Kovacs, N.A.; Bernier, C.R.; Lanier, K.A.; Fox, G.E.; Harvey, S.C.; Wartell, R.M.; Hud, N.V.; et al. History of the ribosome and the origin of translation. Proc. Natl. Acad. Sci. USA 2015, 112, 15396–15401. [Google Scholar] [CrossRef] [Green Version]
- Gulen, B.; Petrov, A.S.; Okafor, C.D.; Vander Wood, D.; O’Neill, E.B.; Hud, N.V.; Williams, L.D. Ribosomal small subunit domains radiate from a central core. Sci. Rep. 2016, 6, 20885. [Google Scholar] [CrossRef]
- Lanier, K.A.; Athavale, S.S.; Petrov, A.S.; Wartell, R.; Williams, L.D. Imprint of ancient evolution on rRNA folding. Biochemistry 2016, 55, 4603–4613. [Google Scholar] [CrossRef]
- Demongeot, J.; Seligmann, H. Accretion history of large ribosomal subunits deduced from theoretical minimal RNA rings is congruent with histories derived from phylogenetic and structural methods. Gene 2020, 738, 144436. [Google Scholar] [CrossRef]
- Demongeot, J.; Seligmann, H. Comparisons between small ribosomal RNA and theoretical minimal RNA ring secondary structures confirm phylogenetic and structural accretion histories. Sci. Rep. 2020, 10, 7693. [Google Scholar] [CrossRef]
- Lovtrup, S. On von Baerian and Haeckelian recapitulation. Syst. Zool. 1978, 27, 348–352. [Google Scholar] [CrossRef]
- Kalinka, A.T.; Tomancak, P. The evolution of early animal embryos: Conservation or divergence. Trends Ecol. Evol. 2012, 27, 385–393. [Google Scholar] [CrossRef]
- Seligmann, H. Evolution and ecology of developmental processes and of the resulting morphology: Directional asymmetry in hindlimbs of Agamidae and Lacertidae (Reptilia: Lacertilia). Biol. J. Linn. Soc. 2000, 69, 461–481. [Google Scholar] [CrossRef]
- Johnson, D.B.F.; Wang, L. Imprints of the genetic code in the ribosome. Proc. Natl. Acad. Sci. USA 2010, 107, 8298–8303. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seligmann, H. Protein sequences recapitulate genetic code evolution. Comput. Struct. Biotechnol. J. 2018, 16, 177–189. [Google Scholar] [CrossRef] [PubMed]
- Gaudart, J.; Landier, J.; Huiart, L.; Legendre, E.; Lehot, L.; Bendiane, M.K.; Chiche, L.; Petitjean, A.; Mosnier, E.; Kirakoya-Samadoulougou, F.; et al. Factors associated with spatial heterogeneity of Covid-19 in France: A nationwide ecological study. Lancet Public Health 2021, 6, e89. [Google Scholar] [CrossRef]
- Lau, H.; Khosrawipour, V.; Kocbach, P.; Mikolajczyk, A.; Schubert, J.; Bania, J.; Khosrawipour, T. The positive impact of lockdown in Wuhan on containing the COVID-19 outbreak in China. J. Travel. Med. 2020, 27, taaa037. [Google Scholar] [CrossRef] [Green Version]
- Bendavid, E.; Oh, C.; Bhattacharya, J.; Ioannidis, J.P.A. Assessing mandatory stay-at-home and business closure effects on the spread of COVID-19. Eur. J. Clin. Investig. 2021, e13484. [Google Scholar]
- Bonacini, L.; Gallo, G.; Patriarca, F. Identifying policy challenges of COVID-19 in hardly reliable data and judging the success of lockdown measures. J. Popul. Econ. 2021, 34, 275–301. [Google Scholar] [CrossRef]
- Oraby, T.; Tyshenko, M.G.; Maldonado, J.C.; Vatcheva, K.; Elsaadany, S.; Alali, W.G.; Longenecker, J.C.; Al-Zoughool, M. Modeling the effect of lockdown timing as a COVID-19 control measure in countries with differing social contacts. Sci. Rep. 2021, 11, 3354. [Google Scholar] [CrossRef]
- Timelli, L.; Girardi, E. Effect of timing of implementation of containment measures on Covid-19 epidemic. The case of the first wave in Italy. PLoS ONE 2021, 16, e0245656. [Google Scholar] [CrossRef] [PubMed]
- Demongeot, J.; Seligmann, H. Covid-19 and miRNA-like inhibition power. Med. Hypotheses 2020, 144C, 110245. [Google Scholar] [CrossRef] [PubMed]
- Demongeot, J.; Griette, Q.; Magal, P. Computations of the transmission rates in SI epidemic model applied to COVID-19 data in mainland China. R. Soc. Open Sci. 2020, 7, 201878. [Google Scholar] [CrossRef] [PubMed]
- Soubeyrand, S.; Demongeot, J.; Roques, L. Towards unified and real-time analyses of outbreaks at country-level during pandemics. One Health 2020, 11, 100187. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| State | T | E | D | A | Date 1 | Slope 1 | Date 2 | Dis | Slope 2 |
|---|---|---|---|---|---|---|---|---|---|
| Alabama | 17.1 | 150 | 37 | 39.2 | 21.03 | 0.3338 | |||
| Alaska | −3 | 580 | 1 | 34.6 | 16.03 | 0.175 | 23.05 | 7 | 0.051 |
| Arizona | 15.7 | 1250 | 23 | 37.9 | 22.03 | 0.1841 | 25.05 | 6 | 0.0733 |
| Arkansas | 15.8 | 200 | 22 | 38.3 | 19.03 | 0.0967 | 10.05 | 8 | 0.0502 |
| California | 15.2 | 880 | 95 | 36.8 | 14.03 | 0.1662 | 10.05 | −4 | 0.0223 |
| Colorado | 7.3 | 2070 | 20 | 36.9 | 20.03 | 0.1989 | 11.06 | 2 | 0.0408 |
| Connecticut | 9.4 | 150 | 286 | 41 | 18.03 | 0.2759 | |||
| DC | 50 | 34 | 21.03 | 0.1011 | 30.06 | 6 | 0.0719 | ||
| Delaware | 12.9 | 20 | 187 | 40.7 | 22.03 | 0.1463 | 6.06 | −6 | 0.0412 |
| Florida | 21.5 | 105 | 145 | 42.2 | 15.03 | 0.2584 | 26.05 | −11 | 0.0785 |
| Georgia | 17.5 | 180 | 68 | 36.9 | 15.03 | 0.2536 | 25.05 | 0 | 0.0293 |
| Hawaii | 21.1 | 920 | 86 | 39.2 | 23.03 | 0.1433 | 2.05 | −21 | 0.0906 |
| Idaho | 6.9 | 1520 | 7 | 36.6 | 24.03 | 0.2793 | 5.06 | −3 | 0.0991 |
| Illinois | 11 | 180 | 89 | 38.3 | 16.03 | 0.4308 | 15.06 | 1 | 0.0289 |
| Indiana | 10.9 | 286 | 71 | 37.9 | 24.03 | 0.1093 | 22.06 | 1 | 0.0348 |
| Iowa | 8.8 | 340 | 21 | 38.2 | 24.03 | 0.1017 | 16.04 | 2 | 0.0893 |
| Kansas | 12.4 | 610 | 14 | 36.9 | 23.03 | 0.0876 | 12.04 | 1 | 0.096 |
| Kentucky | 13.1 | 230 | 43 | 38.9 | 23.03 | 0.1743 | 10.6 | −7 | 0.0344 |
| Louisiana | 19.1 | 30 | 41 | 37.7 | 15.03 | 0.2022 | 23.05 | −6 | 0.0382 |
| Maine | 5 | 180 | 16 | 44.9 | 21.03 | 0.1092 | 18.04 | 0 | 0.0373 |
| Maryland | 12.3 | 110 | 238 | 38.8 | 25.03 | 0.1216 | 14.04 | 0 | 0.0127 |
| Massachusetts | 8.8 | 150 | 336 | 39.4 | 13.03 | 0.3015 | 15.06 | −33 | 0.0129 |
| Michigan | 6.9 | 270 | 67 | 39.8 | 18.03 | 0.4173 | 15.06 | 2 | 0.0612 |
| Minnesota | 5.1 | 370 | 26 | 38.1 | 20.03 | 0.1194 | 14.04 | −6 | 0.1172 |
| Mississippi | 17.4 | 90 | 24 | 37.7 | 20.03 | 0.0863 | |||
| Missouri | 12.5 | 240 | 34 | 38.7 | 21.03 | 0.1762 | 29.05 | 14 | 0.0307 |
| Montana | 5.9 | 1040 | 2 | 39.9 | 14.03 | 0.2485 | 5.06 | 4 | 0.0965 |
| North Carolina | 15 | 210 | 79 | 38.9 | 19.03 | 0.1458 | 4.05 | 0 | 0.0272 |
| North Dakota | 4.7 | 580 | 4 | 35.2 | 27.03 | 0.0545 | 8.05 | 7 | 0.0914 |
| Nebraska | 9.3 | 790 | 9 | 36.6 | 29.03 | 0.0775 | 19.06 | −7 | 0.0248 |
| Nevada | 9.9 | 1680 | 10 | 38.1 | 18.03 | 0.1992 | 23.06 | −2 | 0.054 |
| New Hampshire | 6.6 | 300 | 57 | 43 | 21.03 | 0.1426 | |||
| New Jersey | 11.5 | 80 | 470 | 40 | 16.03 | 0.3797 | |||
| New Mexico | 11.9 | 1740 | 17 | 38.1 | 24.03 | 0.1126 | 9.06 | −5 | 0.0347 |
| New York | 7.4 | 300 | 162 | 39 | 10.03 | 0.5503 | |||
| Ohio | 10.4 | 260 | 109 | 39.4 | 19.03 | 0.2369 | 15.04 | 1 | 0.2485 |
| Oklahoma | 15.3 | 400 | 22 | 36.7 | 24.03 | 0.1267 | 8.06 | 2 | 0.1285 |
| Oregon | 9.1 | 1010 | 16 | 39.4 | 19.03 | 0.106 | 17.05 | −9 | 0.0564 |
| Pennsylvania | 9.3 | 340 | 110 | 40.8 | 17.03 | 0.2935 | 14.06 | 1 | 0.0472 |
| Rhode Island | 10.1 | 60 | 394 | 40.1 | 21.03 | 0.146 | 24.05 | 1 | 0.0435 |
| South Carolina | 16.9 | 110 | 62 | 39.6 | 18.03 | 0.1536 | 24.05 | 0 | 0.0609 |
| South Dakota | 7.3 | 670 | 4 | 37.1 | 29.03 | 0.1716 | |||
| Tennessee | 14.2 | 270 | 61 | 38.8 | 17.03 | 0.2258 | |||
| Texas | 18.2 | 520 | 40 | 34.8 | 17.03 | 0.241 | 20.4 | 1 | 0.0281 |
| Utah | 9.2 | 1860 | 14 | 31 | 19.03 | 0.1146 | 11.05 | −6 | 0.0329 |
| Vermont | 6.1 | 300 | 26 | 42.8 | 22.03 | 0.0546 | |||
| Virginia | 12.8 | 290 | 81 | 38.4 | 19.03 | 0.1789 | 15.06 | 0 | 0.0173 |
| West Virginia | 11 | 460 | 29 | 42.7 | 23.03 | 0.0985 | 5.06 | 1 | 0.0565 |
| Washington | 9.1 | 520 | 41 | 37.2 | 14.03 | 0.0657 | 14.04 | 0 | 0.0494 |
| Wisconsin | 6.2 | 320 | 41 | 39.6 | 18.03 | 0.0910 | 12.04 | 0 | 0.0200 |
| Wyoming | 5.6 | 2040 | 2 | 38 | 28.03 | 0.2346 | 29.05 | 0 | 0.0646 |
| P K-S | 0.907 | 0.008 | 0.007 | 0.530 | 0.274 | 0.228 |
| State\Day | 20 | 40 | 60 | 80 | 100 | 120 | 140 | 160 | 180 | 200 | 220 | 240 | 260 | 280 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Alabama | 24.82 | 0.51 | 2.67 | 4.61 | 6.49 | 5.17 | −0.59 | −4.30 | −0.26 | 2.37 | 0.88 | 1.71 | 1.07 | 2.13 |
| Alaska | 14.70 | −8.70 | −2.76 | 14.64 | 0.52 | 6.41 | 5.18 | 0.60 | 0.35 | 1.64 | 2.81 | 3.03 | 0.29 | −4.71 |
| Arizona | 28.61 | −0.07 | 2.09 | 1.79 | 8.31 | 1.97 | −0.58 | −6.02 | −3.77 | 3.92 | 4.05 | 4.03 | 6.06 | 2.25 |
| Arkansas | 23.24 | 1.66 | −4.17 | 12.99 | 3.53 | −0.57 | 0.39 | −1.08 | 1.24 | 1.19 | −0.71 | −0.15 | 0.72 | 1.30 |
| California | 26.21 | 0.06 | 0.55 | 2.83 | 1.54 | 3.88 | 0.30 | 0.94 | −1.79 | −0.50 | −0.06 | 3.14 | 7.85 | 6.02 |
| Colorado | 23.83 | −0.17 | 0.42 | −2.01 | −4.76 | 2.25 | 1.56 | −2.61 | 0.59 | 5.25 | 2.59 | 5.33 | 1.36 | −2.47 |
| Connecticut | 32.48 | 3.46 | −3.92 | −3.20 | −4.99 | 3.62 | 3.10 | 1.25 | 5.22 | −1.33 | 2.86 | 4.44 | 0.24 | −0.64 |
| DC | 18.43 | 1.18 | 2.00 | −2.08 | −4.00 | 2.50 | 1.52 | −1.83 | −1.70 | 0.98 | 4.37 | 4.06 | 2.96 | 1.71 |
| Delaware | 28.25 | −4.30 | 9.34 | −4.42 | −2.68 | 5.02 | −0.38 | 0.44 | −0.16 | 1.33 | 2.04 | 2.78 | 5.10 | 3.54 |
| Florida | 22.08 | −2.26 | −0.88 | −0.08 | 7.87 | 5.21 | −1.21 | −3.72 | −1.81 | 0.11 | 1.26 | 3.72 | 2.18 | 1.86 |
| Georgia | 25.76 | −0.58 | 0.03 | 0.60 | 2.28 | 5.68 | 0.41 | −1.63 | −1.06 | −1.14 | −0.25 | 1.87 | 2.48 | 4.07 |
| Hawaii | 22.85 | −7.89 | −9.60 | −3.84 | 16.15 | 5.10 | 3.19 | 6.53 | 0.84 | −3.67 | −2.12 | 0.70 | −0.43 | 1.59 |
| Idaho | 32.63 | −9.28 | −5.88 | 10.37 | 4.65 | 6.68 | −0.40 | −1.76 | −1.81 | 4.39 | 1.93 | 1.50 | 1.40 | −0.50 |
| Illinois | 23.16 | 1.72 | 2.51 | −1.82 | −5.17 | 1.47 | 1.68 | 0.61 | 0.53 | 0.47 | 2.26 | 4.61 | −0.67 | −0.52 |
| Indiana | 34.25 | 0.06 | 0.50 | −0.26 | −0.38 | 3.05 | 1.49 | −0.26 | 0.08 | 0.83 | 3.39 | 4.90 | 1.09 | 0.23 |
| Iowa | 26.93 | 6.86 | 1.39 | −2.82 | −0.96 | 4.40 | 1.35 | 0.95 | −1.62 | 2.78 | 2.55 | 6.94 | −4.71 | −1.99 |
| Kansas | 24.21 | 1.49 | 3.91 | −4.31 | 3.20 | 5.06 | −3.57 | −2.44 | −2.71 | 1.11 | 2.23 | 0.11 | 2.03 | −1.89 |
| Kentucky | 23.89 | 4.07 | 0.83 | −1.77 | −3.16 | 3.47 | 2.67 | −0.02 | 1.91 | 1.60 | 0.86 | 2.50 | 2.57 | 0.39 |
| Louisiana | 29.89 | −7.88 | −2.11 | −0.57 | 2.96 | 9.53 | 0.65 | −2.44 | 0.31 | −1.71 | −4.58 | −0.96 | 4.92 | 1.24 |
| Maine | 22.35 | −0.72 | 5.22 | 2.02 | −3.99 | −2.01 | 1.06 | −1.42 | 0.17 | 2.29 | 0.41 | 7.93 | 1.97 | 5.99 |
| Maryland | 30.02 | 5.99 | 2.67 | −1.11 | −5.58 | 0.26 | 3.50 | −1.58 | 0.94 | −1.87 | 1.25 | 2.11 | 3.17 | 1.53 |
| Massachusetts | 33.10 | 2.80 | −2.26 | −3.74 | −8.44 | 0.53 | 2.26 | −1.50 | 0.58 | 1.50 | 1.08 | 3.73 | 1.75 | 2.75 |
| Michigan | 37.94 | −4.00 | −2.67 | −2.12 | −4.92 | 3.78 | 1.31 | −0.70 | 0.28 | 0.30 | 2.46 | 5.67 | −0.76 | −5.05 |
| Minnesota | 19.01 | 4.60 | 10.34 | 0.17 | −2.17 | 2.23 | 0.68 | −0.94 | 2.25 | 2.73 | 1.34 | 5.57 | 1.74 | −4.48 |
| Mississippi | 32.25 | 4.00 | 0.99 | 2.18 | 0.03 | 4.08 | 3.05 | −2.57 | 0.33 | −1.73 | 0.05 | −2.05 | 0.65 | 1.93 |
| Missouri | 27.29 | −0.48 | −2.61 | 0.99 | 0.45 | 2.81 | 3.59 | 4.79 | −0.07 | 1.11 | 1.81 | 2.43 | −2.17 | −2.36 |
| Montana | 23.86 | −6.12 | −12.88 | 0.37 | 1.80 | 6.77 | 2.18 | 1.47 | 3.11 | 2.09 | 5.24 | 1.53 | 2.02 | 0.28 |
| Nebraska | 19.14 | 6.55 | 4.39 | 0.15 | −3.17 | 1.24 | 2.96 | −1.57 | −0.87 | 3.00 | 3.52 | 5.69 | −3.28 | −4.55 |
| Nevada | 26.55 | −0.48 | −1.13 | 1.49 | 3.87 | 5.54 | 1.94 | −2.36 | −3.26 | −0.86 | 0.93 | 1.23 | 3.48 | 1.25 |
| New Hampshire | 21.28 | 0.15 | 1.25 | −1.06 | −5.27 | −0.77 | −0.26 | −2.45 | 2.87 | −1.76 | 3.97 | 4.10 | 3.68 | 2.58 |
| New Jersey | 37.46 | 0.15 | −3.57 | −2.64 | −4.82 | −0.34 | 0.72 | 1.42 | 1.39 | 2.89 | 3.09 | 4.41 | 2.64 | 0.98 |
| New Mexico | 14.28 | 2.46 | 1.26 | −1.04 | −1.64 | 3.72 | 1.49 | −3.76 | −1.33 | 4.51 | 5.04 | 2.87 | 3.75 | −1.11 |
| New York | 24.05 | −2.39 | −3.75 | −2.75 | −3.60 | −1.07 | −1.39 | 0.34 | 2.43 | −0.17 | 2.13 | 2.71 | 2.96 | 1.72 |
| North Carolina | 29.43 | 1.70 | 2.24 | 2.55 | 2.25 | 1.67 | −0.78 | −4.92 | −0.54 | 1.87 | 1.86 | 1.48 | 3.16 | 3.14 |
| North Dakota | 16.30 | 6.56 | −1.08 | −3.77 | −1.37 | 6.40 | 1.57 | 0.54 | 1.57 | 4.27 | 3.21 | 4.11 | −1.41 | −4.33 |
| Ohio | 28.97 | 5.10 | −1.13 | −0.19 | 0.40 | 2.70 | −0.40 | −1.75 | 2.29 | −1.13 | 2.33 | 3.81 | 3.65 | 1.40 |
| Oklahoma | 28.27 | −3.56 | 0.02 | −1.77 | 7.42 | 3.31 | 2.65 | −2.22 | 0.92 | 2.81 | 0.65 | 1.26 | 0.55 | 0.97 |
| Oregon | 20.27 | −1.79 | 1.86 | −4.64 | 7.45 | 2.49 | 0.80 | −0.93 | −0.16 | 1.03 | 0.64 | 3.44 | 2.56 | −0.62 |
| Pennsylvania | 36.04 | 0.85 | 0.71 | −2.20 | −2.91 | 3.20 | 1.79 | −0.87 | 1.90 | 0.38 | 2.64 | 3.76 | 3.93 | 2.27 |
| Rhode Island | 22.39 | 3.75 | −2.60 | −4.36 | −4.45 | −0.31 | 4.36 | 0.15 | −2.40 | 2.90 | 4.50 | 3.31 | 1.90 | −3.86 |
| South Carolina | 25.64 | −1.69 | 0.97 | 1.62 | 5.17 | 2.57 | −1.47 | −3.29 | 2.79 | −0.94 | −0.61 | 0.54 | 2.35 | 4.14 |
| South Dakota | 28.01 | 5.63 | 3.39 | −1.53 | −1.63 | 0.91 | 3.10 | 1.72 | 0.78 | 5.42 | 5.22 | 2.97 | −1.94 | −2.23 |
| Tennessee | 26.84 | 6.02 | 0.08 | 0.87 | 1.11 | 5.39 | 2.35 | −1.62 | 0.78 | 0.35 | 2.85 | 0.38 | −0.82 | 3.97 |
| Texas | 27.73 | 0.05 | 2.81 | −0.58 | 4.55 | 4.74 | −1.22 | −2.03 | −2.72 | 1.57 | 2.34 | 4.11 | 1.29 | 3.50 |
| Utah | 27.54 | 0.26 | 0.87 | 0.11 | 1.53 | 0.46 | −1.55 | −1.20 | 1.68 | 4.67 | 0.97 | 1.45 | 0.94 | −0.63 |
| Vermont | 22.07 | −8.53 | −0.13 | 8.59 | −2.92 | −2.88 | −3.21 | 8.34 | 1.07 | −1.86 | 9.30 | 4.99 | 9.21 | 3.59 |
| Virginia | 23.59 | 3.99 | 2.08 | 1.06 | −4.81 | 0.55 | 1.80 | −0.06 | 1.31 | −1.40 | 1.70 | 1.57 | 3.72 | 3.33 |
| W Virginia | 26.40 | −2.56 | −1.81 | 1.89 | 4.96 | 8.56 | 1.41 | 0.19 | 3.71 | 1.37 | 3.18 | 4.71 | 1.54 | −0.02 |
| Washington | 8.22 | −6.72 | 1.03 | 0.59 | 0.89 | 3.15 | 0.85 | −1.31 | −2.75 | 2.27 | 2.45 | 4.66 | 2.43 | 0.46 |
| Wisconsin | 23.99 | 0.01 | 4.04 | 4.10 | −0.82 | 3.90 | 0.40 | −0.82 | 1.67 | 4.20 | 0.65 | 2.05 | −3.06 | −1.76 |
| Wyoming | 17.98 | −3.48 | 8.95 | 0.17 | 9.48 | 3.00 | 3.04 | −4.67 | −2.25 | 5.93 | 3.27 | 3.86 | 1.90 | −3.35 |
| N > 0 | 51 | 29 | 32 | 24 | 26 | 44 | 37 | 16 | 31 | 37 | 45 | 48 | 41 | 31 |
| Mean | 25.38 | −0.038 | 0.323 | 0.316 | 0.397 | 3.069 | 1.086 | −0.831 | 0.253 | 1.321 | 2.075 | 3.033 | 1.765 | 0.491 |
| P K-S | 0.776 | 0.399 | 0.529 | 0.113 | 0.754 | 0.989 | 0.883 | 0.545 | 0.867 | 0.920 | 0.794 | 0.790 | 0.654 | 0.670 |
| Country\Day | 20 | 40 | 60 | 80 | 100 | 120 | 140 | 160 | 180 | 200 | 220 | 240 | 260 | 280 | 300 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Afghanistan | 15.28 | 5.00 | 7.26 | 5.32 | −1.29 | −2.26 | −0.48 | −6.00 | −6.08 | −1.22 | 11.19 | 1.60 | 4.80 | 0.36 | |
| Albania | 14.49 | −0.84 | −5.79 | −2.20 | 5.02 | 0.97 | 1.30 | 1.88 | −1.00 | 1.65 | 0.89 | 2.66 | 4.13 | 1.11 | |
| Armenia | 16.65 | −0.79 | 5.72 | 7.07 | 2.12 | −0.68 | −4.33 | −0.87 | 0.48 | 1.45 | 3.92 | 1.69 | −4.83 | −3.31 | −5.01 |
| Australia | 25.07 | −4.71 | −11.82 | −0.04 | 0.83 | 11.06 | 9.07 | 2.56 | −7.88 | −4.05 | −7.59 | −0.08 | −3.59 | −1.24 | 7.08 |
| Austria | 30.23 | 4.16 | −10.29 | −4.76 | −6.94 | 1.50 | 5.69 | 2.13 | 5.72 | 0.12 | 2.43 | 4.22 | 7.34 | −2.42 | −2.19 |
| Bangladesh | 5.26 | 24.62 | 4.00 | 3.71 | 2.10 | −0.57 | −0.49 | 1.25 | −2.11 | −1.02 | 0.53 | 1.10 | 2.44 | −1.72 | |
| Belgium | 29.94 | 5.88 | −2.79 | −3.27 | −5.26 | −1.73 | 1.74 | 6.97 | −0.49 | 2.05 | 3.52 | 9.38 | −8.02 | −4.66 | −0.16 |
| Bolivia | 14.65 | 7.18 | 8.70 | 5.72 | 1.63 | 2.12 | 1.10 | 0.23 | −2.76 | −0.96 | −2.08 | −1.72 | 1.90 | 6.46 | |
| Bosnia | 18.43 | 4.10 | −0.38 | −5.19 | 5.36 | 6.15 | 0.90 | 0.32 | 1.22 | −0.42 | −2.27 | 7.65 | −2.09 | −2.28 | −3.57 |
| Brazil | 33.73 | 8.57 | 5.18 | 6.82 | 2.17 | 2.05 | −0.93 | 1.46 | −0.73 | −2.52 | −0.86 | 0.53 | 1.65 | 2.28 | 0.04 |
| Canada | 25.04 | 14.58 | 1.87 | −1.82 | −2.58 | −4.04 | 0.10 | 1.97 | 0.88 | 2.09 | 4.29 | 1.54 | 2.68 | 1.28 | 1.28 |
| Chile | 31.06 | 6.44 | 1.73 | 5.41 | 1.47 | −1.95 | −2.31 | −0.97 | −1.28 | 0.70 | 1.06 | 0.00 | 2.05 | 0.88 | 1.54 |
| Czechia | 30.07 | 2.34 | −6.20 | −1.42 | −2.70 | 2.91 | −4.90 | 1.87 | 0.34 | 8.70 | 2.59 | 8.27 | −2.65 | −1.15 | 5.81 |
| Georgia | 7.56 | 5.78 | −0.16 | 0.08 | 2.65 | −6.34 | 3.13 | −6.75 | −2.31 | 15.53 | 6.30 | 8.63 | 4.03 | 0.63 | −6.27 |
| Germany | 35.16 | 7.43 | −5.98 | −5.09 | −5.06 | 3.36 | −2.52 | 4.94 | 4.69 | −2.04 | 2.07 | 6.73 | 4.30 | 0.30 | 3.30 |
| Greece | 18.28 | −1.52 | −3.81 | −2.05 | 6.38 | 0.93 | 1.25 | 12.28 | 1.55 | 0.62 | 0.30 | 5.47 | 2.79 | −3.74 | −4.50 |
| Hungary | 19.48 | 6.03 | −0.99 | −3.38 | −7.37 | −1.73 | 6.18 | 1.61 | 2.73 | 10.63 | 1.61 | 4.88 | 3.25 | 1.92 | −4.47 |
| India | 19.78 | 14.27 | 4.25 | 5.48 | 3.25 | 3.71 | 3.24 | 2.23 | 0.40 | 1.75 | −0.67 | −1.73 | 0.01 | −0.20 | −2.43 |
| Indonesia | 31.30 | 6.62 | 0.21 | 3.57 | 3.00 | 1.29 | 0.54 | 0.85 | 2.18 | 1.26 | −0.09 | −0.51 | 2.31 | 1.57 | 1.37 |
| Iran | 40.40 | 6.66 | −4.41 | −0.19 | 1.85 | −0.87 | 0.10 | 0.02 | −0.48 | −1.72 | 2.88 | 1.14 | 4.16 | 2.47 | −2.79 |
| Iraq | 14.82 | 8.94 | −5.01 | 3.21 | 9.96 | 3.94 | 1.20 | 2.04 | 3.05 | 0.49 | −0.95 | −1.32 | −0.96 | −2.68 | −2.93 |
| Ireland | 30.39 | 6.56 | −5.86 | −7.61 | −10.15 | −4.52 | 4.66 | 9.26 | 1.82 | 4.31 | 3.79 | 3.43 | −3.89 | −1.98 | 8.65 |
| Israel | 28.61 | 12.23 | −2.47 | −11.53 | 12.25 | 5.22 | 7.81 | −0.06 | 1.18 | 3.82 | 1.80 | −10.38 | 0.55 | 1.70 | 3.79 |
| Italy | 35.43 | 6.45 | −1.99 | −5.16 | −5.06 | −2.65 | −2.39 | 1.62 | 4.41 | 6.08 | 0.35 | 7.27 | 6.76 | −0.68 | −2.62 |
| Kazakhstan | 21.10 | 5.98 | 0.55 | 6.71 | −0.22 | 8.67 | −0.94 | −6.96 | −7.08 | −1.47 | 2.07 | 9.07 | 1.27 | −0.50 | |
| Kuwait | 4.17 | 6.19 | 3.04 | 7.45 | −0.56 | −2.01 | 0.33 | 0.15 | 1.49 | 0.99 | −1.78 | 2.36 | −0.29 | −4.74 | −1.18 |
| Lithuania | 21.70 | −3.11 | −9.79 | −2.26 | −0.20 | 2.46 | 6.22 | 4.34 | −0.17 | 5.18 | 1.44 | 8.25 | 0.67 | 1.40 | |
| Luxemburg | 29.22 | −8.42 | −6.55 | −6.81 | 0.46 | 14.02 | 5.36 | −5.47 | 0.30 | 2.60 | 4.25 | 7.57 | −2.28 | 0.16 | |
| Malaysia | 31.13 | 1.47 | −6.38 | −2.08 | 4.30 | −3.72 | 2.53 | 3.78 | 3.42 | 6.77 | 4.92 | 5.48 | 0.82 | 1.23 | 2.00 |
| Mexico | 26.38 | 8.11 | 4.82 | 3.36 | 1.52 | 0.79 | 0.89 | −0.24 | 0.39 | −1.99 | 1.88 | 0.09 | −0.83 | 1.38 | 0.45 |
| Moldova | 19.26 | 1.80 | 0.03 | 1.23 | 4.22 | −4.23 | 0.68 | 1.60 | 0.83 | −0.64 | 0.09 | −0.30 | 1.41 | −0.78 | |
| Morocco | 25.02 | 6.01 | −1.31 | −5.95 | 4.98 | 6.50 | 4.17 | 3.80 | −0.23 | 2.65 | 2.04 | 0.16 | −1.43 | −1.66 | |
| N Macedonia | 19.60 | 5.10 | −8.86 | 1.80 | 10.53 | 0.38 | 0.31 | −0.69 | −4.68 | 3.92 | 5.90 | 5.55 | 1.80 | −3.23 | |
| Netherlands | 33.96 | 6.11 | −1.67 | −4.50 | −2.12 | −6.07 | −1.05 | 8.25 | 1.24 | 2.96 | 6.15 | 4.91 | −3.62 | −0.79 | 5.22 |
| Norway | 32.24 | 3.04 | −4.34 | −4.61 | −5.32 | −4.10 | −8.34 | 10.91 | 2.80 | 5.89 | −0.32 | 1.04 | 5.25 | −2.75 | 1.26 |
| Pakistan | 32.05 | 5.03 | 6.72 | 2.79 | 7.68 | −2.83 | −5.12 | −5.37 | −3.39 | 2.12 | −0.64 | 3.03 | 4.91 | 0.68 | −3.49 |
| Panama | 25.65 | 2.63 | 0.32 | 1.01 | 4.09 | 1.48 | −1.03 | −0.60 | 0.20 | −0.35 | 1.03 | 1.06 | 4.05 | 3.42 | 1.44 |
| Philippines | 23.02 | 1.90 | −0.50 | −0.89 | 3.50 | 3.59 | 2.59 | 5.20 | −1.10 | 0.90 | −1.60 | −2.30 | −1.77 | −0.62 | 0.97 |
| Poland | 32.90 | 6.28 | −0.37 | 1.45 | 1.00 | −2.38 | 0.93 | 4.22 | 1.28 | −3.03 | 6.77 | 7.00 | 1.16 | −4.49 | −0.04 |
| Portugal | 32.57 | 3.86 | −4.33 | −1.82 | 2.48 | 1.43 | 0.19 | −2.99 | −0.43 | 4.72 | 1.36 | 7.84 | 3.59 | −4.45 | −1.16 |
| Romania | 25.79 | 3.61 | −0.81 | −4.61 | 0.44 | 1.88 | 5.16 | 0.03 | −0.50 | 1.01 | 3.58 | 2.86 | 1.94 | −1.54 | −3.23 |
| Russia | 24.49 | 15.79 | 5.52 | −0.90 | −0.10 | −1.62 | −0.46 | −0.76 | −0.46 | 0.81 | 3.51 | 1.89 | 1.35 | 1.18 | 0.40 |
| S Korea | 39.90 | −5.49 | −8.29 | −5.98 | 7.20 | 3.22 | 0.91 | −2.40 | −3.56 | 13.72 | −5.90 | −0.99 | 1.88 | 5.77 | 3.62 |
| Saudi Arabia | 26.61 | 6.47 | 6.99 | 3.68 | 1.52 | −0.50 | −2.70 | −3.71 | −1.80 | −2.88 | −2.03 | −0.83 | −1.19 | −1.99 | 0.01 |
| Serbia | 24.03 | 8.49 | −5.76 | −1.45 | 5.40 | 9.67 | 1.79 | −3.79 | −6.40 | 0.43 | 4.49 | 14.24 | 7.72 | 0.85 | −3.55 |
| Turkey | 42.42 | 3.06 | −4.63 | −2.49 | 3.35 | −1.02 | −0.54 | 1.67 | 1.82 | 0.18 | 0.31 | 1.41 | 6.30 | 1.44 | |
| UAE | 5.12 | 16.42 | 1.89 | 2.99 | −2.70 | −0.51 | −2.32 | −1.13 | 3.67 | 4.10 | 2.32 | 1.31 | −0.34 | 0.03 | 0.45 |
| UK | 35.66 | 14.62 | 0.24 | −1.45 | −4.09 | −2.13 | −3.28 | 0.73 | 2.28 | 4.96 | 4.96 | 3.90 | 0.68 | −3.83 | 4.09 |
| Ukraine | 27.04 | 10.09 | −0.42 | −0.49 | 3.69 | −0.96 | 2.53 | 2.75 | 1.24 | 1.99 | 2.88 | 3.10 | 2.34 | −1.29 | |
| USA | 35.93 | 24.23 | 0.89 | −0.29 | −0.67 | 0.17 | 3.35 | 0.60 | −0.80 | −1.64 | 0.54 | 0.79 | 3.63 | 2.38 | 1.99 |
| Uzbekistan | 14.33 | 1.90 | 1.02 | 3.81 | 4.71 | 4.62 | 1.32 | −0.89 | −3.71 | 2.43 | −2.68 | −2.47 | −1.17 | 1.02 | |
| N > 0 | 51.00 | 44.00 | 21.00 | 21.00 | 33.00 | 27.00 | 33.00 | 33.00 | 27.00 | 36.00 | 37.00 | 39.00 | 35.00 | 25.00 | 21.00 |
| Mean | 25.15 | 6.02 | −1.20 | −0.35 | 1.35 | 0.88 | 0.85 | 1.06 | −0.15 | 2.03 | 1.58 | 2.87 | 1.31 | −0.33 | 0.14 |
| P K-S | 0.730 | 0.109 | 0.927 | 0.871 | 0.880 | 0.657 | 0.532 | 0.324 | 0.430 | 0.224 | 0.994 | 0.599 | 0.933 | 0.962 | 0.868 |
| Period | 20 | 40 | 60 | 80 | 100 | 120 | 140 | 160 | 180 | 200 | 220 | 240 | 260 | 280 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 20 | 8.44 | −20.28 | −14.06 | −12.91 | 2.06 | −5.35 | 1.08 | 19.97 | −20.66 | −16.90 | −13.99 | 1.25 | 22.38 | |
| 40 | −12.19 | 34.97 | −35.06 | −41.88 | −27.16 | 19.09 | −20.75 | 8.95 | 11.97 | 9.16 | 12.29 | −32.21 | −10.46 | |
| 60 | −30.70 | 46.88 | −13.30 | −14.46 | −21.45 | −17.35 | −37.87 | −25.24 | 27.82 | 4.15 | 26.25 | 5.78 | −1.48 | |
| 80 | −43.48 | 19.60 | 60.91 | 18.56 | 3.12 | −7.80 | 0.98 | −3.24 | 2.78 | 1.03 | −19.44 | 9.14 | 7.52 | |
| 100 | −16.05 | −12.48 | 2.74 | 14.58 | 45.82 | −16.25 | −15.34 | −29.57 | −0.86 | −34.56 | −38.75 | 0.67 | 10.38 | |
| 120 | −3.47 | −28.73 | −29.64 | −9.71 | 30.60 | 10.69 | −14.23 | −11.02 | −2.38 | −31.61 | −31.53 | −16.50 | −14.27 | |
| 140 | −10.69 | −13.90 | −38.06 | −35.52 | 10.33 | 39.24 | 16.86 | 10.11 | 6.74 | −2.21 | 3.73 | −42.42 | −38.85 | |
| 160 | 18.81 | −3.84 | −31.98 | −36.01 | −28.21 | −25.49 | 4.07 | 33.33 | −29.27 | 22.71 | 13.97 | −7.00 | −7.52 | |
| 180 | 19.53 | 13.42 | −11.88 | −35.10 | −34.54 | −36.00 | −9.64 | 49.67 | −24.47 | −2.14 | −2.01 | −10.99 | 0.32 | |
| 200 | −1.15 | −18.88 | −28.08 | −37.32 | −7.39 | −25.39 | 1.72 | −4.11 | 15.19 | 30.54 | 28.37 | −32.57 | −49.68 | |
| 220 | 0.65 | 17.93 | 6.56 | −8.7 | −23.01 | −35.36 | −7.58 | −8.67 | 10.00 | 11.19 | 46.64 | 10.23 | −23.15 | |
| 240 | 5.33 | −19.27 | −32.94 | −13.92 | −32.56 | 2.86 | −9.33 | −4.04 | −3.33 | 19.67 | 40.71 | −8.76 | −22.56 | |
| 260 | 15.67 | 14.91 | −4.48 | −3.11 | 4.91 | −15.03 | −14.76 | −22.09 | 4.90 | 3.98 | 9.66 | 10.70 | 56.77 | |
| 280 | 14.85 | 0.86 | 13.4 | 1.48 | 16.47 | 11.04 | 12.40 | −29.26 | −19.98 | 11.38 | −20.61 | −31.81 | 31.23 | |
| 300 | 43.39 | 2.23 | −32.48 | −29.66 | −26.21 | 4.74 | 4.05 | 33.80 | −4.15 | −4.35 | −10.73 | −26.07 | −35.84 | 12.88 |
| Variable\Day | 20 | 40 | 60 | 80 | 100 | 120 | 140 | 160 | 180 | 200 | 220 | 240 | 260 | 280 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Temp. USA | 25.12 | 4.70 | −7.16 | −16.21 | 47.06 | 21.18 | −21.98 | −19.91 | −21.60 | −40.10 | −50.69 | −49.46 | 16.29 | 51.98 |
| World | −19.04 | 4.51 | 23.03 | 34.31 | 31.66 | 15.94 | 7.09 | −6.62 | −5.28 | −23.11 | −38.76 | −38.72 | −10.68 | 13.34 |
| Altitude USA | −23.09 | −18.78 | −1.66 | 6.53 | 32.71 | 11.81 | 1.28 | −19.04 | −29.20 | 49.98 | 14.48 | 1.34 | 3.90 | −28.26 |
| rs | −4.8 | −19.9 | 1.5 | 12.8 | 32.31 | 3.85 | −8.56 | 0.52 | −6.69 | 42.04 | 21.23 | 6.97 | −1.97 | −23.1 |
| World | −11.64 | −10.61 | 29.92 | 33.46 | 18.73 | −3.51 | −13.12 | −35.17 | −30.11 | −5.92 | 7.80 | −24.34 | 12.08 | 22.81 |
| rs | −4.9 | −8.9 | 22.88 | 19.79 | 27.61 | 6.34 | −5.22 | −36.68 | −32.67 | −13.16 | −1.60 | −21.75 | 26.36 | 25.92 |
| Density USA | 38.85 | 17.62 | −17.62 | −36.69 | −41.00 | −31.03 | 14.32 | 13.56 | 14.52 | −18.43 | 3.41 | 10.16 | 14.58 | 12.67 |
| rs | 11.54 | 26.24 | −10.30 | −36.64 | −34.65 | −5.49 | 7.61 | 16.86 | 12.70 | −38.32 | −9.86 | 10.74 | 8.77 | 20.96 |
| World | −19.90 | 24.74 | 6.45 | −4.70 | 4.56 | −4.78 | 1.62 | 10.29 | 7.00 | 13.47 | −13.05 | −10.58 | −11.70 | −14.60 |
| rs | 8.63 | −5.25 | −20.88 | −25.96 | 7.27 | 0.91 | 4.33 | 19.35 | 32.83 | 36.69 | 0.22 | 12.44 | −10.41 | −23.4 |
| Age USA | 15.17 | −9.38 | −5.15 | −7.11 | −13.10 | −21.44 | −7.13 | 17.25 | 32.06 | −33.31 | 11.68 | 25.91 | 21.84 | 30.23 |
| World | 33.71 | −16.10 | −50.50 | −54.53 | −39.52 | −2.62 | 9.08 | 30.14 | 25.74 | 29.10 | 12.27 | 57.83 | 7.62 | −22.37 |
| Country | Temperature | Elevation | Density | Age |
|---|---|---|---|---|
| Afghanistan | 12.6 | 1885 | 49 | 18.9 |
| Albania | 11.4 | 708 | 99.73 | 32.9 |
| Armenia | 7.15 | 1792 | 99 | 35.1 |
| Australia | 21.65 | 330 | 3 | 38.7 |
| Austria | 6.35 | 910 | 106 | 44 |
| Bangladesh | 25 | 85 | 1175 | 26.7 |
| Belgium | 9.55 | 181 | 378 | 41.4 |
| Bolivia | 21.55 | 1192 | 10 | 24.3 |
| Bosnia | 9.85 | 500 | 69 | 42.1 |
| Brazil | 24.95 | 320 | 25 | 32.6 |
| Canada | −5.35 | 487 | 4 | 42.2 |
| Chile | 8.45 | 1871 | 23 | 34.4 |
| Czech | 7.55 | 433 | 135 | 42.1 |
| Georgia | 5.8 | 1432 | 53.51 | 38.1 |
| Germany | 8.5 | 263 | 233 | 47.1 |
| Greece | 15.4 | 498 | 210 | 44.5 |
| Hungary | 9.75 | 143 | 272 | 42.3 |
| India | 23.65 | 621 | 412 | 28.1 |
| Indonesia | 25.85 | 367 | 141 | 30.2 |
| Iran | 17.25 | 1305 | 51 | 30.3 |
| Iraq | 14.03 | 312 | 90 | 20 |
| Ireland | 9.3 | 118 | 70 | 36.8 |
| Israel | 19.2 | 508 | 417 | 29.9 |
| Italy | 12.45 | 538 | 200 | 45.5 |
| Kazakhstan | 6.4 | 387 | 7 | 30.6 |
| Kuwait | 25.35 | 108 | 642 | 29.3 |
| Lithuania | 6.2 | 110 | 73 | 43.7 |
| Luxemburg | 8.65 | 325 | 237.39 | 39.3 |
| Malaysia | 25.4 | 538 | 99 | 28.5 |
| Mexico | 21 | 1111 | 64 | 28.3 |
| Moldova | 9.45 | 139 | 79.27 | 36.7 |
| Morocco | 17.1 | 909 | 80 | 29.3 |
| N Macedonia | 9.8 | 741 | 81 | 37.9 |
| Netherlands | 9.25 | 30 | 421 | 42.6 |
| Norway | 1.5 | 460 | 17 | 39.2 |
| Pakistan | 20.2 | 900 | 274 | 23.8 |
| Panama | 25.4 | 360 | 56 | 29.2 |
| Philippines | 25.85 | 442 | 362 | 23.5 |
| Poland | 7.85 | 173 | 123 | 40.7 |
| Portugal | 15.15 | 372 | 112 | 42.2 |
| Romania | 8.8 | 414 | 81.4 | 41.1 |
| Russia | −5.1 | 600 | 9 | 39.6 |
| S Korea | 11.5 | 282 | 517 | 41.8 |
| Saudi Arabia | 24.65 | 665 | 16 | 27.5 |
| Serbia | 10.55 | 473 | 89.08 | 42.6 |
| Turkey | 11.1 | 1132 | 106.12 | 30.9 |
| UAE | 27 | 149 | 116.87 | 30.3 |
| UK | 8.45 | 162 | 280 | 40.5 |
| Ukraine | 8.3 | 175 | 69.49 | 40.6 |
| USA | 8.55 | 760 | 34 | 38.1 |
| Uzbekistan | 12.05 | 2750 | 73 | 28.6 |
| P K-S | 0.199 | 0.024 | 0.002 | 0.259 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Seligmann, H.; Vuillerme, N.; Demongeot, J. Unpredictable, Counter-Intuitive Geoclimatic and Demographic Correlations of COVID-19 Spread Rates. Biology 2021, 10, 623. https://doi.org/10.3390/biology10070623
Seligmann H, Vuillerme N, Demongeot J. Unpredictable, Counter-Intuitive Geoclimatic and Demographic Correlations of COVID-19 Spread Rates. Biology. 2021; 10(7):623. https://doi.org/10.3390/biology10070623
Chicago/Turabian StyleSeligmann, Hervé, Nicolas Vuillerme, and Jacques Demongeot. 2021. "Unpredictable, Counter-Intuitive Geoclimatic and Demographic Correlations of COVID-19 Spread Rates" Biology 10, no. 7: 623. https://doi.org/10.3390/biology10070623
APA StyleSeligmann, H., Vuillerme, N., & Demongeot, J. (2021). Unpredictable, Counter-Intuitive Geoclimatic and Demographic Correlations of COVID-19 Spread Rates. Biology, 10(7), 623. https://doi.org/10.3390/biology10070623