
Interleukin-10 Deficiency Impacts on TNF-Induced NFκB Regulated Responses In Vivo

 ,
,  , ,
, , 

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Mice
2.2. RNA Extraction and qPCR
2.3. Immunohistochemistry
2.4. NF-κB p65[RelA] Activity ELISA
2.5. ELISA for Serum and Tissue Cytokines, Chemokines and Immunoglobulins
2.6. Immunoblot Analysis of A20 and IκBα Protein in Small Intestinal Tissue Lysates
2.7. Data Analyses
3. Results
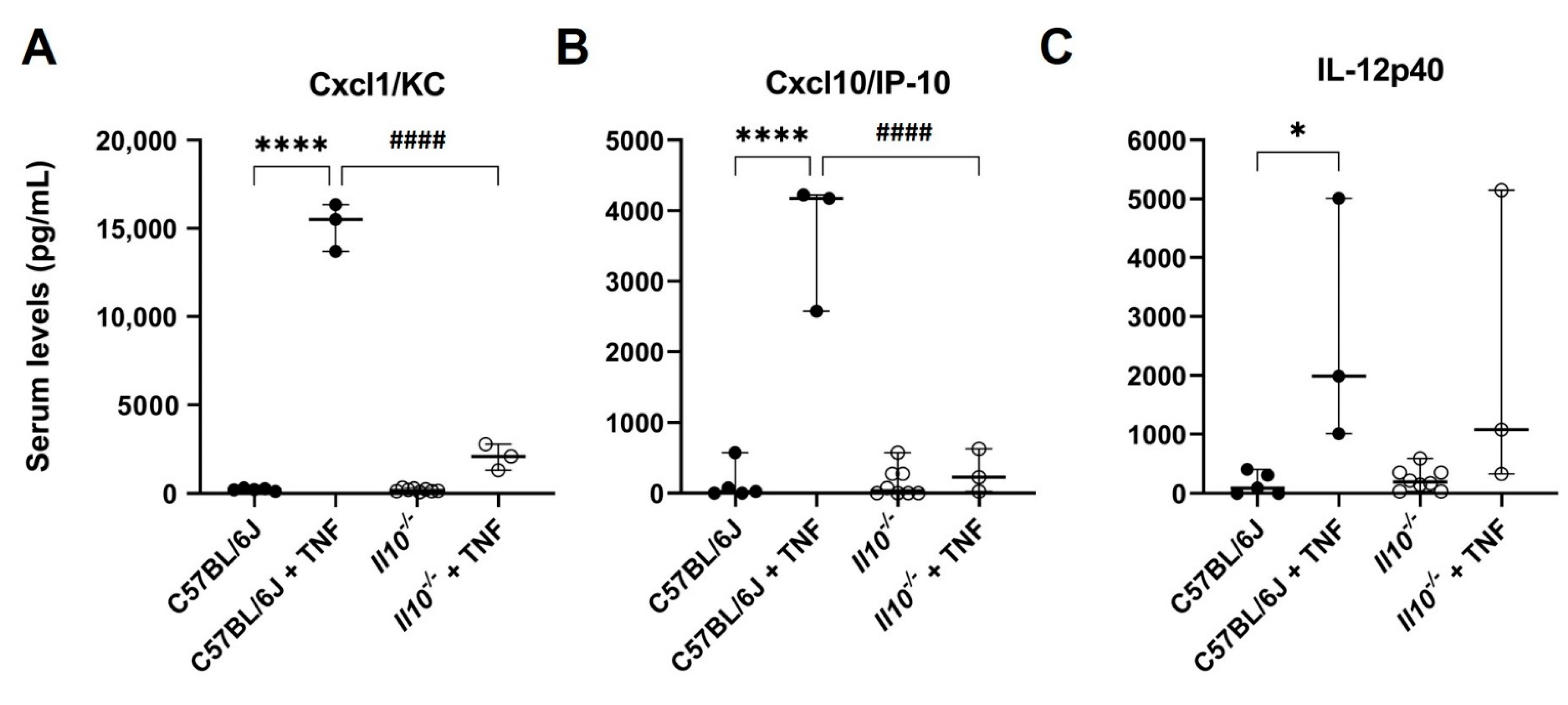
3.1. TNF-Induced NFκB-Dependent Serum Cytokine/Chemokine Responses Were Attenuated in IL-10-Deficient Mice
3.2. Dysregulated Expression of TNF in the Resting Small Intestine of IL-10-Deficient Mice
3.3. Impaired NFκB Activation and Inducible TNF Expression in the Small Intestine of IL-10-Deficient Mice
3.4. Altered A20 and IκBα Synthesis Observed in the Small Intestine of Il10−/− Mice
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wei, H.X.; Wang, B.; Li, B. IL-10 and IL-22 in Mucosal Immunity: Driving Protection and Pathology. Front. Immunol. 2020, 11, 1315. [Google Scholar] [CrossRef] [PubMed]
- Kamanaka, M.; Kim, S.T.; Wan, Y.Y.; Sutterwala, F.S.; Lara-Tejero, M.; Galan, J.E.; Harhaj, E.; Flavell, R.A. Expression of interleukin-10 in intestinal lymphocytes detected by an interleukin-10 reporter knockin tiger mouse. Immunity 2006, 25, 941–952. [Google Scholar] [CrossRef]
- Bando, J.K.; Gilfillan, S.; Di Luccia, B.; Fachi, J.L.; Secca, C.; Cella, M.; Colonna, M. ILC2s are the predominant source of intestinal ILC-derived IL-10. J. Exp. Med. 2020, 217, e20191520. [Google Scholar] [CrossRef]
- Morhardt, T.L.; Hayashi, A.; Ochi, T.; Quiros, M.; Kitamoto, S.; Nagao-Kitamoto, H.; Kuffa, P.; Atarashi, K.; Honda, K.; Kao, J.Y.; et al. IL-10 produced by macrophages regulates epithelial integrity in the small intestine. Sci. Rep. 2019, 9, 1223. [Google Scholar] [CrossRef] [PubMed]
- Krause, P.; Morris, V.; Greenbaum, J.A.; Park, Y.; Bjoerheden, U.; Mikulski, Z.; Muffley, T.; Shui, J.W.; Kim, G.; Cheroutre, H.; et al. IL-10-producing intestinal macrophages prevent excessive antibacterial innate immunity by limiting IL-23 synthesis. Nat. Commun. 2015, 6, 7055. [Google Scholar] [CrossRef]
- Nguyen, H.D.; Aljamaei, H.M.; Stadnyk, A.W. The Production and Function of Endogenous Interleukin-10 in Intestinal Epithelial Cells and Gut Homeostasis. Cell. Mol. Gastroenterol. Hepatol. 2021, 12, 1343–1352. [Google Scholar] [CrossRef] [PubMed]
- Denning, T.L.; Campbell, N.A.; Song, F.; Garofalo, R.P.; Klimpel, G.R.; Reyes, V.E.; Ernst, P.B. Expression of IL-10 receptors on epithelial cells from the murine small and large intestine. Int. Immunol. 2000, 12, 133–139. [Google Scholar] [CrossRef] [PubMed]
- Liu, G.; Wang, B.; Chen, Q.; Li, Y.; Li, B.; Yang, N.; Yang, S.; Geng, S.; Liu, G. Interleukin (IL)-21 Promotes the Differentiation of IgA-Producing Plasma Cells in Porcine Peyer’s Patches via the JAK-STAT Signaling Pathway. Front. Immunol. 2020, 11, 1303. [Google Scholar] [CrossRef]
- Takeuchi, T.; Ohno, H. Reciprocal regulation of IgA and the gut microbiota: A key mutualism in the intestine. Int. Immunol. 2021, 33, 781–786. [Google Scholar] [CrossRef]
- Suzuki, K.; Fagarasan, S. Diverse regulatory pathways for IgA synthesis in the gut. Mucosal Immunol. 2009, 2, 468–471. [Google Scholar] [CrossRef] [Green Version]
- Gomes-Santos, A.C.; Moreira, T.G.; Castro-Junior, A.B.; Horta, B.C.; Lemos, L.; Cruz, D.N.; Guimaraes, M.A.; Cara, D.C.; McCafferty, D.M.; Faria, A.M. New insights into the immunological changes in IL-10-deficient mice during the course of spontaneous inflammation in the gut mucosa. Clin. Dev. Immunol. 2012, 2012, 560817. [Google Scholar] [CrossRef]
- Zigmond, E.; Bernshtein, B.; Friedlander, G.; Walker, C.R.; Yona, S.; Kim, K.W.; Brenner, O.; Krauthgamer, R.; Varol, C.; Muller, W.; et al. Macrophage-restricted interleukin-10 receptor deficiency, but not IL-10 deficiency, causes severe spontaneous colitis. Immunity 2014, 40, 720–733. [Google Scholar] [CrossRef]
- Redhu, N.S.; Bakthavatchalu, V.; Conaway, E.A.; Shouval, D.S.; Tsou, A.; Goettel, J.A.; Biswas, A.; Wang, C.; Field, M.; Muller, W.; et al. Macrophage dysfunction initiates colitis during weaning of infant mice lacking the interleukin-10 receptor. Elife 2017, 6, e27652. [Google Scholar] [CrossRef] [PubMed]
- Chaudhry, A.; Samstein, R.M.; Treuting, P.; Liang, Y.; Pils, M.C.; Heinrich, J.M.; Jack, R.S.; Wunderlich, F.T.; Bruning, J.C.; Muller, W.; et al. Interleukin-10 signaling in regulatory T cells is required for suppression of Th17 cell-mediated inflammation. Immunity 2011, 34, 566–578. [Google Scholar] [CrossRef] [PubMed]
- Kuhn, R.; Lohler, J.; Rennick, D.; Rajewsky, K.; Muller, W. Interleukin-10-deficient mice develop chronic enterocolitis. Cell 1993, 75, 263–274. [Google Scholar] [CrossRef]
- Spencer, S.D.; Di Marco, F.; Hooley, J.; Pitts-Meek, S.; Bauer, M.; Ryan, A.M.; Sordat, B.; Gibbs, V.C.; Aguet, M. The orphan receptor CRF2-4 is an essential subunit of the interleukin 10 receptor. J. Exp. Med. 1998, 187, 571–578. [Google Scholar] [CrossRef]
- Glocker, E.O.; Kotlarz, D.; Klein, C.; Shah, N.; Grimbacher, B. IL-10 and IL-10 receptor defects in humans. Ann. N. Y. Acad. Sci. 2011, 1246, 102–107. [Google Scholar] [CrossRef] [PubMed]
- Nunberg, M.Y.; Werner, L.; Kopylov, U.; Haberman, Y.; Lahad, A.; Weiss, B.; Shouval, D.S. Impaired IL-10 Receptor-mediated Suppression in Monocyte From Patients With Crohn Disease. J. Pediatr. Gastroenterol. Nutr. 2018, 66, 779–784. [Google Scholar] [CrossRef] [PubMed]
- Paul, G.; Khare, V.; Gasche, C. Inflamed gut mucosa: Downstream of interleukin-10. Eur. J. Clin. Investig. 2012, 42, 95–109. [Google Scholar] [CrossRef] [PubMed]
- Engelhardt, K.R.; Grimbacher, B. IL-10 in humans: Lessons from the gut, IL-10/IL-10 receptor deficiencies, and IL-10 polymorphisms. Curr. Top. Microbiol. Immunol. 2014, 380, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Hutchins, A.P.; Diez, D.; Miranda-Saavedra, D. The IL-10/STAT3-mediated anti-inflammatory response: Recent developments and future challenges. Brief. Funct. Genom. 2013, 12, 489–498. [Google Scholar] [CrossRef] [PubMed]
- Liu, T.; Zhang, L.; Joo, D.; Sun, S.C. NF-kappaB signaling in inflammation. Signal Transduct. Target. Ther. 2017, 2, 17023. [Google Scholar] [CrossRef]
- Kondylis, V.; Kumari, S.; Vlantis, K.; Pasparakis, M. The interplay of IKK, NF-kappaB and RIPK1 signaling in the regulation of cell death, tissue homeostasis and inflammation. Immunol. Rev. 2017, 277, 113–127. [Google Scholar] [CrossRef] [PubMed]
- Papoutsopoulou, S.; Pollock, L.; Walker, C.; Tench, W.; Samad, S.S.; Bergey, F.; Lenzi, L.; Sheibani-Tezerji, R.; Rosenstiel, P.; Alam, M.T.; et al. Impact of Interleukin 10 Deficiency on Intestinal Epithelium Responses to Inflammatory Signals. Front. Immunol. 2021, 12, 690817. [Google Scholar] [CrossRef] [PubMed]
- Williams, J.M.; Duckworth, C.A.; Watson, A.J.; Frey, M.R.; Miguel, J.C.; Burkitt, M.D.; Sutton, R.; Hughes, K.R.; Hall, L.J.; Caamano, J.H.; et al. A mouse model of pathological small intestinal epithelial cell apoptosis and shedding induced by systemic administration of lipopolysaccharide. Dis. Model. Mech. 2013, 6, 1388–1399. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Jones, L.G.; Vaida, A.; Thompson, L.M.; Ikuomola, F.I.; Caamano, J.H.; Burkitt, M.D.; Miyajima, F.; Williams, J.M.; Campbell, B.J.; Pritchard, D.M.; et al. NF-kappaB2 signalling in enteroids modulates enterocyte responses to secreted factors from bone marrow-derived dendritic cells. Cell Death Dis. 2019, 10, 896. [Google Scholar] [CrossRef] [PubMed]
- Verstrepen, L.; Carpentier, I.; Beyaert, R. The biology of A20-binding inhibitors of NF-kappaB activation (ABINs). Adv. Exp. Med. Biol. 2014, 809, 13–31. [Google Scholar] [CrossRef] [PubMed]
- Saraiva, M.; Vieira, P.; O’Garra, A. Biology and therapeutic potential of interleukin-10. J. Exp. Med. 2020, 217, e20190418. [Google Scholar] [CrossRef] [PubMed]
- Van Hauwermeiren, F.; Vandenbroucke, R.E.; Grine, L.; Lodens, S.; Van Wonterghem, E.; De Rycke, R.; De Geest, N.; Hassan, B.; Libert, C. TNFR1-induced lethal inflammation is mediated by goblet and Paneth cell dysfunction. Mucosal Immunol. 2015, 8, 828–840. [Google Scholar] [CrossRef] [Green Version]
- Roulis, M.; Armaka, M.; Manoloukos, M.; Apostolaki, M.; Kollias, G. Intestinal epithelial cells as producers but not targets of chronic TNF suffice to cause murine Crohn-like pathology. Proc. Natl. Acad. Sci. USA 2011, 108, 5396–5401. [Google Scholar] [CrossRef] [PubMed]
- Lefrancois, L.; Lycke, N. Isolation of mouse small intestinal intraepithelial lymphocytes, Peyer’s patch, and lamina propria cells. Curr. Protoc. Immunol. 1996, 17, 3.19.1–3.19.16. [Google Scholar] [CrossRef]
- Lindsay, J.O.; Hodgson, H.J. Review article: The immunoregulatory cytokine interleukin-10--a therapy for Crohn’s disease? Aliment. Pharmacol. Ther. 2001, 15, 1709–1716. [Google Scholar] [CrossRef] [PubMed]
- Kullberg, M.C.; Ward, J.M.; Gorelick, P.L.; Caspar, P.; Hieny, S.; Cheever, A.; Jankovic, D.; Sher, A. Helicobacter hepaticus triggers colitis in specific-pathogen-free interleukin-10 (IL-10)-deficient mice through an IL-12- and gamma interferon-dependent mechanism. Infect. Immun. 1998, 66, 5157–5166. [Google Scholar] [CrossRef]
- Sellon, R.K.; Tonkonogy, S.; Schultz, M.; Dieleman, L.A.; Grenther, W.; Balish, E.; Rennick, D.M.; Sartor, R.B. Resident enteric bacteria are necessary for development of spontaneous colitis and immune system activation in interleukin-10-deficient mice. Infect. Immun. 1998, 66, 5224–5231. [Google Scholar] [CrossRef]
- Harris, D.P.; Bandyopadhyay, S.; Maxwell, T.J.; Willard, B.; DiCorleto, P.E. Tumor necrosis factor (TNF)-alpha induction of CXCL10 in endothelial cells requires protein arginine methyltransferase 5 (PRMT5)-mediated nuclear factor (NF)-kappaB p65 methylation. J. Biol. Chem. 2014, 289, 15328–15339. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Ao, X.; Shen, Z.; Ao, L.; Wu, X.; Pu, C.; Guo, W.; Xing, W.; He, M.; Yuan, H.; et al. TNF-alpha augments CXCL10/CXCR3 axis activity to induce Epithelial-Mesenchymal Transition in colon cancer cell. Int. J. Biol. Sci. 2021, 17, 2683–2702. [Google Scholar] [CrossRef]
- Rahim, S.S.; Khan, N.; Boddupalli, C.S.; Hasnain, S.E.; Mukhopadhyay, S. Interleukin-10 (IL-10) mediated suppression of IL-12 production in RAW 264.7 cells also involves c-rel transcription factor. Immunology 2005, 114, 313–321. [Google Scholar] [CrossRef]
- Moore, K.W.; de Waal Malefyt, R.; Coffman, R.L.; O’Garra, A. Interleukin-10 and the interleukin-10 receptor. Annu. Rev. Immunol. 2001, 19, 683–765. [Google Scholar] [CrossRef]
- Choe, J.; Choi, Y.S. IL-10 interrupts memory B cell expansion in the germinal center by inducing differentiation into plasma cells. Eur J. Immunol. 1998, 28, 508–515. [Google Scholar] [CrossRef]
- Shparago, N.; Zelazowski, P.; Jin, L.; McIntyre, T.M.; Stuber, E.; Pecanha, L.M.; Kehry, M.R.; Mond, J.J.; Max, E.E.; Snapper, C.M. IL-10 selectively regulates murine Ig isotype switching. Int. Immunol. 1996, 8, 781–790. [Google Scholar] [CrossRef] [PubMed]
- Briere, F.; Servet-Delprat, C.; Bridon, J.M.; Saint-Remy, J.M.; Banchereau, J. Human interleukin 10 induces naive surface immunoglobulin D+ (sIgD+) B cells to secrete IgG1 and IgG3. J. Exp. Med. 1994, 179, 757–762. [Google Scholar] [CrossRef] [PubMed]
- Malisan, F.; Briere, F.; Bridon, J.M.; Harindranath, N.; Mills, F.C.; Max, E.E.; Banchereau, J.; Martinez-Valdez, H. Interleukin-10 induces immunoglobulin G isotype switch recombination in human CD40-activated naive B lymphocytes. J. Exp. Med. 1996, 183, 937–947. [Google Scholar] [CrossRef] [PubMed]
- Bagheri, Y.; Babaha, F.; Falak, R.; Yazdani, R.; Azizi, G.; Sadri, M.; Abolhassani, H.; Shekarabi, M.; Aghamohammadi, A. IL-10 induces TGF-beta secretion, TGF-beta receptor II upregulation, and IgA secretion in B cells. Eur. Cytokine Netw. 2019, 30, 107–113. [Google Scholar] [CrossRef]
- Pers, J.O.; Jamin, C.; Youinou, P.; Charreire, J. Role of IL-10 in the distribution of B cell subsets in the mouse B-1 cell population. Eur. Cytokine Netw. 2003, 14, 178–185. [Google Scholar]
- Stavnezer, J.; Kang, J. The surprising discovery that TGF beta specifically induces the IgA class switch. J. Immunol. 2009, 182, 5–7. [Google Scholar] [CrossRef]
- Lycke, N.Y.; Bemark, M. The regulation of gut mucosal IgA B-cell responses: Recent developments. Mucosal Immunol. 2017, 10, 1361–1374. [Google Scholar] [CrossRef]
- Ruder, B.; Atreya, R.; Becker, C. Tumour Necrosis Factor Alpha in Intestinal Homeostasis and Gut Related Diseases. Int. J. Mol. Sci. 2019, 20, 1887. [Google Scholar] [CrossRef]
- Papoutsopoulou, S.; Campbell, B.J. Epigenetic Modifications of the Nuclear Factor Kappa B Signalling Pathway and its Impact on Inflammatory Bowel Disease. Curr. Pharm. Des. 2021, 27, 3702–3713. [Google Scholar] [CrossRef]
- Gilmore, T.D.; Wolenski, F.S. NF-kappaB: Where did it come from and why? Immunol. Rev. 2012, 246, 14–35. [Google Scholar] [CrossRef]
- O’Shea, J.M.; Perkins, N.D. Regulation of the RelA (p65) transactivation domain. Biochem. Soc. Trans. 2008, 36, 603–608. [Google Scholar] [CrossRef] [PubMed]
- Kattah, M.G.; Shao, L.; Rosli, Y.Y.; Shimizu, H.; Whang, M.I.; Advincula, R.; Achacoso, P.; Shah, S.; Duong, B.H.; Onizawa, M.; et al. A20 and ABIN-1 synergistically preserve intestinal epithelial cell survival. J. Exp. Med. 2018, 215, 1839–1852. [Google Scholar] [CrossRef] [PubMed]
- Shouval, D.S.; Ouahed, J.; Biswas, A.; Goettel, J.A.; Horwitz, B.H.; Klein, C.; Muise, A.M.; Snapper, S.B. Interleukin 10 receptor signaling: Master regulator of intestinal mucosal homeostasis in mice and humans. Adv. Immunol. 2014, 122, 177–210. [Google Scholar] [CrossRef] [PubMed]
- Minshawi, F.; Lanvermann, S.; McKenzie, E.; Jeffery, R.; Couper, K.; Papoutsopoulou, S.; Roers, A.; Muller, W. The Generation of an Engineered Interleukin-10 Protein With Improved Stability and Biological Function. Front. Immunol. 2020, 11, 1794. [Google Scholar] [CrossRef]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Papoutsopoulou, S.; Pollock, L.; Williams, J.M.; Abdul-Mahdi, M.M.L.F.; Dobbash, R.; Duckworth, C.A.; Campbell, B.J. Interleukin-10 Deficiency Impacts on TNF-Induced NFκB Regulated Responses In Vivo. Biology 2022, 11, 1377. https://doi.org/10.3390/biology11101377
Papoutsopoulou S, Pollock L, Williams JM, Abdul-Mahdi MMLF, Dobbash R, Duckworth CA, Campbell BJ. Interleukin-10 Deficiency Impacts on TNF-Induced NFκB Regulated Responses In Vivo. Biology. 2022; 11(10):1377. https://doi.org/10.3390/biology11101377
Chicago/Turabian StylePapoutsopoulou, Stamatia, Liam Pollock, Jonathan M. Williams, Maya M. L. F. Abdul-Mahdi, Reyhaneh Dobbash, Carrie A. Duckworth, and Barry J. Campbell. 2022. "Interleukin-10 Deficiency Impacts on TNF-Induced NFκB Regulated Responses In Vivo" Biology 11, no. 10: 1377. https://doi.org/10.3390/biology11101377
APA StylePapoutsopoulou, S., Pollock, L., Williams, J. M., Abdul-Mahdi, M. M. L. F., Dobbash, R., Duckworth, C. A., & Campbell, B. J. (2022). Interleukin-10 Deficiency Impacts on TNF-Induced NFκB Regulated Responses In Vivo. Biology, 11(10), 1377. https://doi.org/10.3390/biology11101377