

A Review of Pseudomonas aeruginosa Metallophores: Pyoverdine, Pyochelin and Pseudopaline


Abstract
:Simple Summary
Abstract
1. Introduction
2. Pyoverdine
2.1. Biosynthesis and Secretion of Pyoverdines
2.2. PVD-Fe Complex Uptake
2.3. Storage of PVDI in the Periplasm
2.4. Bacterial Virulence and PVDs
2.5. Iron Uptake Regulation
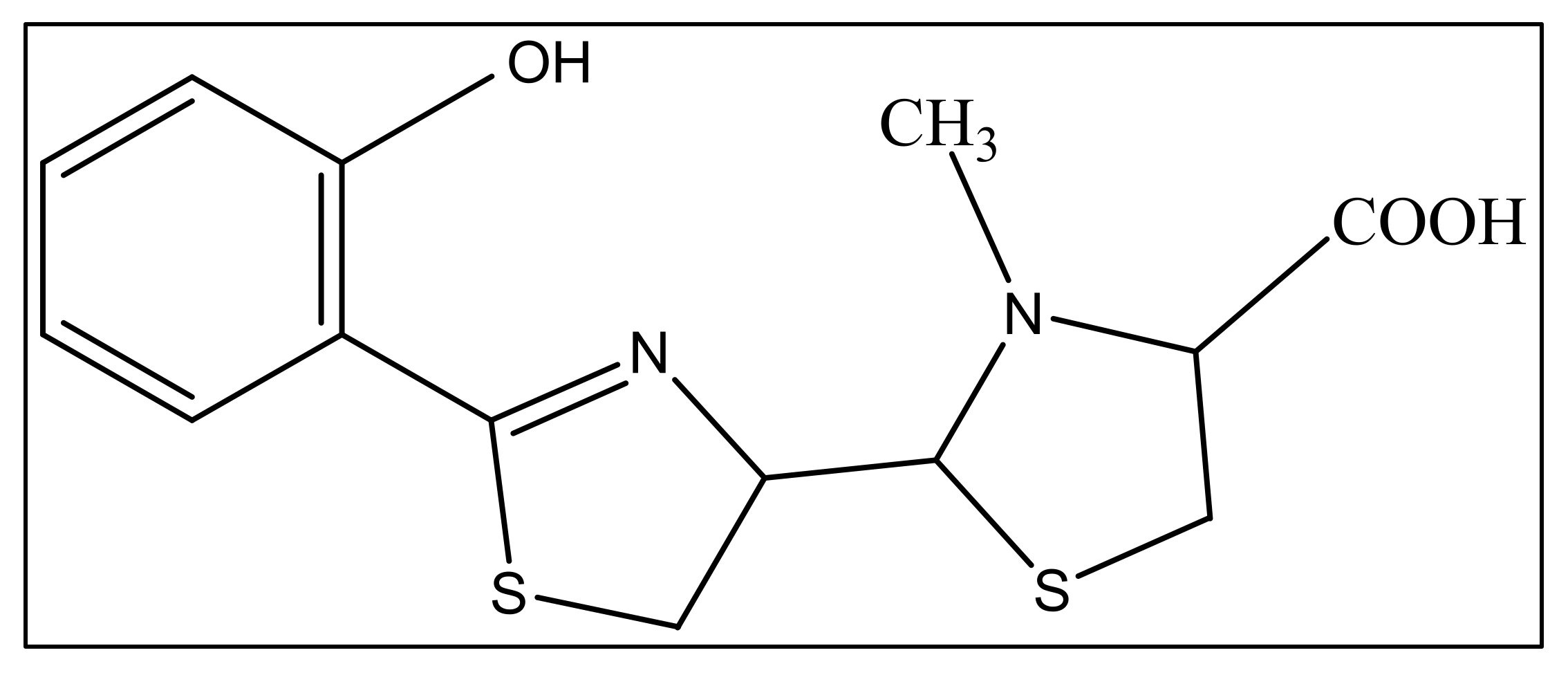
3. Pyochelin
3.1. Pyochelin Biosynthesis
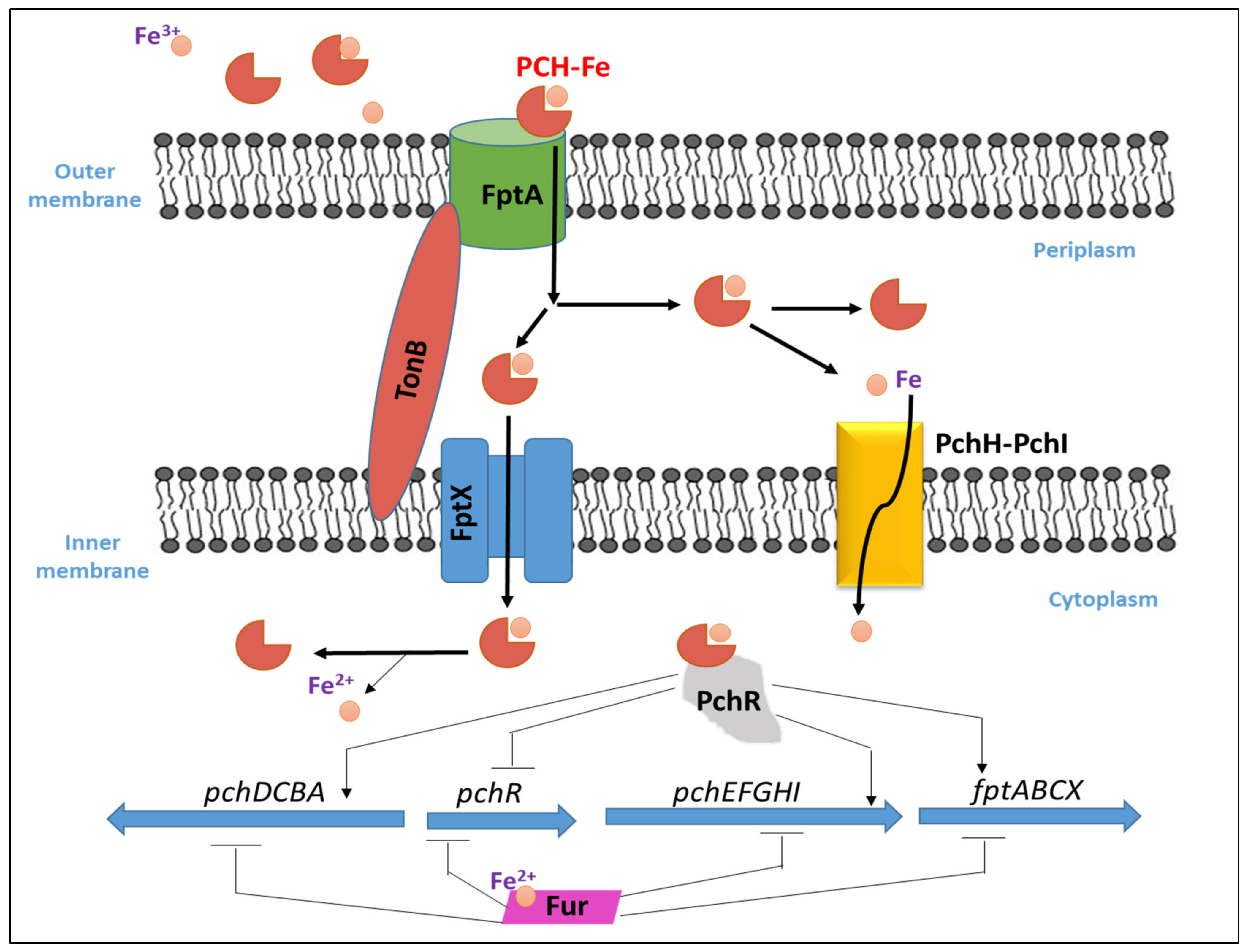
3.2. Pyochelin–Fe Uptake and Regulation
3.3. PCH Metal Chelation
4. Uptake of Siderophores Produced by Other Microorganisms
5. Pseudopaline
Pseudopaline Biosynthesis, Excretion, Import and Regulation
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Peix, A.; Ramírez-Bahena, M.-H.; Velázquez, E. Historical evolution and current status of the taxonomy of genus Pseudomonas. Infect. Genet. Evol. 2009, 9, 1132–1147. [Google Scholar] [CrossRef] [PubMed]
- Anzai, Y.; Kim, H.; Park, J.-Y.; Wakabayashi, H.; Oyaizu, H. Phylogenetic affiliation of the pseudomonads based on 16S rRNA sequence. Int. J. Syst. Evol. Microbiol. 2000, 50, 1563–1589. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wolfgang, M.C.; Kulasekara, B.R.; Liang, X.; Boyd, D.; Wu, K.; Yang, Q.; Miyada, C.G.; Lory, S. Conservation of genome content and virulence determinants among clinical and environmental isolates of Pseudomonas aeruginosa. Proc. Natl. Acad. Sci. USA 2003, 100, 8484–8489. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Silhavy, T.J.; Kahne, D.; Walker, S. The Bacterial Cell Envelope. Cold Spring Harb. Perspect. Biol. 2010, 2, a000414. [Google Scholar] [CrossRef] [PubMed]
- Livermore, D.M. Current Epidemiology and Growing Resistance of Gram-Negative Pathogens. Korean J. Intern. Med. 2012, 27, 128–142. [Google Scholar] [CrossRef]
- Mahajan-Miklos, S.; Rahme, L.G.; Ausubel, F.M. Elucidating the molecular mechanisms of bacterial virulence using non-mammalian hosts. Mol. Microbiol. 2000, 37, 981–988. [Google Scholar] [CrossRef]
- Winstanley, C.; O’Brien, S.; Brockhurst, M.A. Pseudomonas aeruginosa Evolutionary Adaptation and Diversification in Cystic Fibrosis Chronic Lung Infections. Trends Microbiol. 2016, 24, 327–337. [Google Scholar] [CrossRef] [Green Version]
- Moradali, M.F.; Ghods, S.; Rehm, B.H.A. Pseudomonas aeruginosa Lifestyle: A Paradigm for Adaptation, Survival, and Persistence. Front. Cell. Infect. Microbiol. 2017, 7, 39. [Google Scholar] [CrossRef] [Green Version]
- Neves, P.R.; McCulloch, J.A.; Mamizuka, E.M.; Lincopan, N. Aeruginosa. In Encyclopedia of Food Microbiology; Batt, C.A., Tortorello, M.-L., Eds.; Elsevier: Amsterdam, The Netherlands, 2014; Volume 3, pp. 253–260. ISBN 9780123847331. [Google Scholar]
- CDC. Antibiotic Resistance Threats in the United States; Centers for Disease Control and Prevention: Atlanta, GA, USA, 2013; 114p. [Google Scholar]
- Aloush, V.; Navon-Venezia, S.; Seigman-Igra, Y.; Cabili, S.; Carmeli, Y. Multidrug-Resistant Pseudomonas aeruginosa: Risk Factors and Clinical Impact. Antimicrob. Agents Chemother. 2006, 50, 43–48. [Google Scholar] [CrossRef] [Green Version]
- Schechner, V.; Gottesman, T.; Schwartz, O.; Korem, M.; Maor, Y.; Rahav, G.; Karplus, R.; Lazarovitch, T.; Braun, E.; Finkelstein, R.; et al. Pseudomonas aeruginosa bacteremia upon hospital admission: Risk factors for mortality and influence of inadequate empirical antimicrobial therapy. Diagn. Microbiol. Infect. Dis. 2011, 71, 38–45. [Google Scholar] [CrossRef]
- Breidenstein, E.B.; de la Fuente-Núñez, C.; Hancock, R.E. Pseudomonas aeruginosa: All roads lead to resistance. Trends Microbiol. 2011, 19, 419–426. [Google Scholar] [CrossRef] [PubMed]
- Lambert, P.A. Mechanisms of antibiotic resistance in Pseudomonas aeruginosa. J. R. Soc. Med. 2002, 95, 22–26. [Google Scholar] [CrossRef] [PubMed]
- Wu, M.; Guina, T.; Brittnacher, M.; Nguyen, H.; Eng, J.; Miller, S.I. The Pseudomonas aeruginosa Proteome during Anaerobic Growth. J. Bacteriol. 2005, 187, 8185–8190. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hall-Stoodley, L.; Costerton, J.W.; Stoodley, P. Bacterial biofilms: From the Natural environment to infectious diseases. Nat. Rev. Genet. 2004, 2, 95–108. [Google Scholar] [CrossRef]
- Lazdunski, A. Regulation of virulence factors in Pseudomonas aeruginosa. Med. Mal. Infect. 1998, 28, 109–118. [Google Scholar]
- Hahn, H.P. The type-4 pilus is the major virulence-associated adhesin of Pseudomonas aeruginosa—A review. Gene 1997, 192, 99–108. [Google Scholar] [CrossRef]
- Wick, M.; Frank, D.; Storey, D.; Iglewski, B. Structure, Function, and Regulation of Pseudomonas aeruginosa Exotoxin A. Annu. Rev. Microbiol. 1990, 44, 335–363. [Google Scholar] [CrossRef]
- Kipnis, E.; Sawa, T.; Wiener-Kronish, J. Targeting mechanisms of Pseudomonas aeruginosa pathogenesis. Med. Mal. Infect. 2006, 36, 78–91. [Google Scholar] [CrossRef]
- Guillon, L.; El Mecherki, M.; Altenburger, S.; Graumann, P.L.; Schalk, I.J. High cellular organization of pyoverdine biosynthesis in Pseudomonas aeruginosa: Clustering of PvdA at the old cell pole. Environ. Microbiol. 2012, 14, 1982–1994. [Google Scholar] [CrossRef]
- Guerra, A.J.; Giedroc, D.P. Metal site occupancy and allosteric switching in bacterial metal sensor proteins. Arch. Biochem. Biophys. 2012, 519, 210–222. [Google Scholar] [CrossRef] [Green Version]
- Foster, A.W.; Osman, D.; Robinson, N.J. Metal Preferences and Metallation. J. Biol. Chem. 2014, 289, 28095–28103. [Google Scholar] [CrossRef] [PubMed]
- Cornelis, P. Iron uptake and metabolism in pseudomonads. Appl. Microbiol. Biotechnol. 2010, 86, 1637–1645. [Google Scholar] [CrossRef] [PubMed]
- Hider, R.C.; Kong, X. Chemistry and biology of siderophores. Nat. Prod. Rep. 2010, 27, 637–657. [Google Scholar] [CrossRef]
- Meyer, J.M.; Neely, A.; Stintzi, A.; Georges, C.; Holder, A.I. Pyoverdin is essential for virulence of Pseudomonas aeruginosa. Infect. Immun. 1996, 64, 518–523. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiong, Y.; Vasil, M.L.; Johnson, Z.; Ochsner, U.A.; Bayer, A.S. The Oxygen-and Iron-Dependent Sigma Factor pvdS of Pseudomonas aeruginosa Is an Important Virulence Factor in Experimental Infective Endocarditis. J. Infect. Dis. 2000, 181, 1020–1026. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Minandri, F.; Imperi, F.; Frangipani, E.; Bonchi, C.; Visaggio, D.; Facchini, M.; Pasquali, P.; Bragonzi, A.; Visca, P. Role of Iron Uptake Systems in Pseudomonas aeruginosa Virulence and Airway Infection. Infect. Immun. 2016, 84, 2324–2335. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lamont, I.L.; Beare, P.A.; Ochsner, U.; Vasil, A.I.; Vasil, M.L. Siderophore-mediated signaling regulates virulence factor production in Pseudomonas aeruginosa. Proc. Natl. Acad. Sci. USA 2002, 99, 7072–7077. [Google Scholar] [CrossRef] [Green Version]
- Wilderman, P.J.; Vasil, A.I.; Johnson, Z.; Wilson, M.J.; Cunliffe, H.E.; Lamont, I.L.; Vasil, M.L. Characterization of an Endoprotease (PrpL) Encoded by a PvdS-Regulated Gene in Pseudomonas aeruginosa. Infect. Immun. 2001, 69, 5385–5394. [Google Scholar] [CrossRef] [Green Version]
- Brandel, J.; Humbert, N.; Elhabiri, M.; Schalk, I.J.; Mislin, G.L.A.; Albrecht-Gary, A.-M. Pyochelin, a siderophore of Pseudomonas aeruginosa: Physicochemical characterization of the iron(iii), copper(ii) and zinc(ii) complexes. Dalton Trans. 2012, 41, 2820–2834. [Google Scholar] [CrossRef]
- Serino, L.; Reimmann, C.; Visca, P.; Beyeler, M.; Chiesa, V.D.; Haas, D. Biosynthesis of pyochelin and dihydroaeruginoic acid requires the iron-regulated pchDCBA operon in Pseudomonas aeruginosa. J. Bacteriol. 1997, 179, 248–257. [Google Scholar] [CrossRef] [Green Version]
- Dumas, Z.; Ross-Gillespie, A.; Kümmerli, R. Switching between apparently redundant iron-uptake mechanisms benefits bacteria in changeable environments. Proc. R. Soc. B Biol. Sci. 2013, 280, 20131055. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lyczak, J.B.; Cannon, C.L.; Pier, G.B. Lung Infections Associated with Cystic Fibrosis. Clin. Microbiol. Rev. 2002, 15, 194–222. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lhospice, S.; Gomez, N.O.; Ouerdane, L.; Brutesco, C.; Ghssein, G.; Hajjar, C.; Liratni, A.; Wang, S.; Richaud, P.; Bleves, S.; et al. Pseudomonas aeruginosa zinc uptake in chelating environment is primarily mediated by the metallophore pseudopaline. Sci. Rep. 2017, 7, 17132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ghssein, G.; Brutesco, C.; Ouerdane, L.; Fojcik, C.; Izaute, A.; Wang, S.; Hajjar, C.; Lobinski, R.; Lemaire, D.; Richaud, P.; et al. Biosynthesis of a broad-spectrum nicotianamine-like metallophore in Staphylococcus aureus. Science 2016, 352, 1105–1109. [Google Scholar] [CrossRef]
- Skaar, E.P. The Battle for Iron between Bacterial Pathogens and Their Vertebrate Hosts. PLOS Pathog. 2010, 6, e1000949. [Google Scholar] [CrossRef] [Green Version]
- Ochsner, A.U.; Vasil, I.A.; Vasil, M.L. Role of the ferric uptake regulator of Pseudomonas aeruginosa in the regulation of siderophores and exotoxin A expression: Purification and activity on iron-regulated promoters. J. Bacteriol. 1995, 177, 7194–7201. [Google Scholar] [CrossRef] [Green Version]
- Troxell, B.; Hassan, H.M. Transcriptional regulation by Ferric Uptake Regulator (Fur) in pathogenic bacteria. Front. Cell Infect. Microbiol. 2013, 3, 59. [Google Scholar] [CrossRef] [Green Version]
- Totter, J.R.; Moseley, F.T. Influence of the concentration of iron on the production of fluorescin by Pseudomonas aeruginosa. J Bacteriol. 1952, 6, 45–47. [Google Scholar] [CrossRef] [Green Version]
- Meyer, J.M.; Hornspreger, J. Role of pyoverdinePf the iron binding fluorescent pigment of Pseudomonas fluorescens iron transport. J. Gen. Microbiol. 1978, 107, 329–331. [Google Scholar] [CrossRef] [Green Version]
- Budzikiewicz, H.; Schäfer, M.; Fernández, D.U.; Matthijs, S.; Cornelis, P. Characterization of the chromophores of pyoverdins and related siderophores by electrospray tandem mass spectrometry. BioMetals 2006, 20, 135–144. [Google Scholar] [CrossRef]
- Boukhalfa, H.; Crumbliss, A.L. Chemical aspects of siderophore mediated iron transport. BioMetals 2002, 15, 325–339. [Google Scholar] [CrossRef] [PubMed]
- Demange, P.; Wendenbaum, S.; Linget, C.; Mertz, C.; Cung, M.T.; Dell, A.; Abdallah, M.A. Bacterial siderophores: Structure and NMR assigment of pyoverdins PaA, siderophores of Pseudomonas aeruginosa ATCC 15692. Biol. Met. 1990, 3, 155–170. [Google Scholar] [CrossRef]
- Schalk, I.J.; Guillon, L. Pyoverdine biosynthesis and secretion in Pseudomonas aeruginosa: Implications for metal homeostasis. Environ. Microbiol. 2012, 15, 1661–1673. [Google Scholar] [CrossRef] [PubMed]
- Fuchs, R.; Schafer, M.; Geoffroy, V.; Meyer, J.-M. Siderotyping A Powerful Tool for the Characterization of Pyoverdines. Curr. Top. Med. Chem. 2001, 1, 31–57. [Google Scholar] [CrossRef] [PubMed]
- Meyer, J.-M.; Stintzi, A.; De Vos, D.; Cornelis, P.; Tappe, R.; Taraz, K.; Budzikiewicz, H. Use of Siderophores to Type Pseudomonads: The Three Pseudomonas Aeruginosa Pyoverdine Systems. Microbiology 1997, 143, 35–43. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Briskot, G.; Taraz, K.; Budzikiewicz, H. ChemInform Abstract: Bacterial Constituents. Part 37. Pyoverdin-Type Siderophores from Pseudomonas aeruginosa. ChemInform 1989, 1989, 375–384. [Google Scholar] [CrossRef]
- Tappe, R.; Tara, K.; Budzikiewicz, H.; Meyer, J.M.; Lefèvre, J.F. Structure elucidation of a pyoverdine produced by Pseudomonas aeruginosa ATCC 27853. J. Prakt. Chem. 1993, 335, 83–87. [Google Scholar] [CrossRef]
- Gipp, S.; Hahn, J.; Taraz, K.; Budzikiewicz, H. Zwei Pyoverdine aus Pseudomonas aeruginosa R./Two Pyoverdins from Pseudomonas aeruginosa R. Z. Naturforsch. C 1991, 46, 534–541. [Google Scholar] [CrossRef]
- Ruangviriyachai, C.; Fernández, D.U.; Fuchs, R.; Meyer, J.-M.; Budzikiewicz, H. A New Pyoverdin from Pseudomonas aeruginosa R’. Z. Naturforsch. C J. Biosci. 2001, 56, 933–938. [Google Scholar] [CrossRef]
- Miethke, M.; Marahiel, M.A. Siderophore-Based Iron Acquisition and Pathogen Control. Microbiol. Mol. Biol. Rev. 2007, 71, 413–451. [Google Scholar] [CrossRef] [Green Version]
- Visca, P.; Imperi, F.; Lamont, I.L. Pyoverdine siderophores: From biogenesis to biosignificance. Trends Microbiol. 2007, 15, 22–30. [Google Scholar] [CrossRef] [PubMed]
- Finking, R.; Marahiel, M.A. Biosynthesis of Nonribosomal Peptides. Annu. Rev. Microbiol. 2004, 58, 453–488. [Google Scholar] [CrossRef] [PubMed]
- Ravel, J.; Cornelis, P. Genomics of pyoverdine-mediated iron uptake in pseudomonads. Trends Microbiol. 2003, 11, 195–200. [Google Scholar] [CrossRef] [PubMed]
- Ackerley, D.F.; Caradoc-Davies, T.T.; Lamont, I.L. Substrate Specificity of the Nonribosomal Peptide Synthetase PvdD from Pseudomonas aeruginosa. J. Bacteriol. 2003, 185, 2848–2855. [Google Scholar] [CrossRef] [Green Version]
- Mossialos, D.; Ochsner, U.; Baysse, C.; Chablain, P.; Pirnay, J.-P.; Koedam, N.; Budzikiewicz, H.; Fernández, D.U.; Schäfer, M.; Ravel, J.; et al. Identification of new, conserved, non-ribosomal peptide synthetases from fluorescent pseudomonads involved in the biosynthesis of the siderophore pyoverdine. Mol. Microbiol. 2002, 45, 1673–1685. [Google Scholar] [CrossRef] [Green Version]
- Drake, E.J.; Gulick, A.M. Structural Characterization and High-Throughput Screening of Inhibitors of PvdQ, an NTN Hydrolase Involved in Pyoverdine Synthesis. ACS Chem. Biol. 2011, 6, 1277–1286. [Google Scholar] [CrossRef] [Green Version]
- Vandenende, C.S.; Vlasschaert, M.; Seah, S.Y.K. Functional Characterization of an Aminotransferase Required for Pyoverdine Siderophore Biosynthesis in Pseudomonas aeruginosa PAO1. J. Bacteriol. 2004, 186, 5596–5602. [Google Scholar] [CrossRef] [Green Version]
- McMorran, B.J.; Kumara, H.M.C.S.; Sullivan, K.; Lamont, I.L. Involvement of a transformylase enzyme in siderophore synthesis in Pseudomonas aeruginosa The GenBank accession number for the sequence reported in this paper is U07359. Microbiology 2001, 147, 1517–1524. [Google Scholar] [CrossRef] [Green Version]
- Yeterian, E.; Martin, L.W.; Guillon, L.; Journet, L.; Lamont, I.; Schalk, I.J. Synthesis of the siderophore pyoverdine in Pseudomonas aeruginosa involves a periplasmic maturation. Amino Acids 2009, 38, 1447–1459. [Google Scholar] [CrossRef]
- Hannauer, M.; Schäfer, M.; Hoegy, F.; Gizzi, P.; Wehrung, P.; Mislin, G.L.; Budzikiewicz, H.; Schalk, I.J. Biosynthesis of the pyoverdine siderophore of Pseudomonas aeruginosa involves precursors with a myristic or a myristoleic acid chain. FEBS Lett. 2011, 586, 96–101. [Google Scholar] [CrossRef] [Green Version]
- Lamont, I.L.; Martin, L.W. Identification and characterization of novel pyoverdine synthesis genes in Pseudomonas aeruginosa. Microbiology 2003, 149, 833–842. [Google Scholar] [CrossRef] [PubMed]
- Schalk, I.J.; Hannauer, M.; Braud, A. New roles for bacterial siderophores in metal transport and tolerance. Environ. Microbiol. 2011, 13, 2844–2854. [Google Scholar] [CrossRef] [PubMed]
- Bokhove, M.; Jimenez, P.N.; Quax, W.J.; Dijkstra, B.W. The quorum-quenching N -acyl homoserine lactone acylase PvdQ is an Ntn-hydrolase with an unusual substrate-binding pocket. Proc. Natl. Acad. Sci. USA 2009, 107, 686–691. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dorrestein, P.C.; Poole, A.K.; Begley, T.P. Formation of the Chromophore of the Pyoverdine Siderophores by an Oxidative Cascade. Org. Lett. 2003, 5, 2215–2217. [Google Scholar] [CrossRef]
- Jacques, P.; Ongena, M.; Bernard, F.; Fuchs, R.; Budzikiewicz, H.; Thonart, P. Fluorescent Pseudomonas mainly produce the dihydroform of pyoverdine at low specific growth rate. Lett. Appl. Microbiol. 2003, 36, 259–262. [Google Scholar] [CrossRef]
- Nadal-Jimenez, P.; Koch, G.; Reis, C.R.; Muntendam, R.; Raj, H.; Jeronimus-Stratingh, C.M.; Cool, R.H.; Quax, W.J. PvdP Is a Tyrosinase That Drives Maturation of the Pyoverdine Chromophore in Pseudomonas aeruginosa. J. Bacteriol. 2014, 196, 2681–2690. [Google Scholar] [CrossRef] [Green Version]
- Hannauer, M.; Yeterian, E.; Martin, L.W.; Lamont, I.L.; Schalk, I.J. An efflux pump is involved in secretion of newly synthesized siderophore by Pseudomonas aeruginosa. FEBS Lett. 2010, 584, 4751–4755. [Google Scholar] [CrossRef] [Green Version]
- Nikaido, H.; Takatsuka, Y. Mechanisms of RND multidrug efflux pumps. Biochim. Biophys. Acta 2009, 1794, 769–781. [Google Scholar] [CrossRef] [Green Version]
- Spencer, D.H.; Kas, A.; Smith, E.E.; Raymond, C.K.; Sims, E.H.; Hastings, M.; Burns, J.L.; Kaul, R.; Olson, M.V. Whole-Genome Sequence Variation among Multiple Isolates of Pseudomonas aeruginosa. J. Bacteriol. 2003, 185, 1316–1325. [Google Scholar] [CrossRef] [Green Version]
- Schalk, I.J.; Kyslik, P.; Prome, D.; van Dorsselaer, A.; Poole, K.; Abdallah, A.M.A.; Pattus, F. Copurification of the FpvA Ferric Pyoverdin Receptor of Pseudomonas aeruginosa with Its Iron-Free Ligand: Implications for Siderophore-Mediated Iron Transport. Biochemistry 1999, 38, 9357–9365. [Google Scholar] [CrossRef]
- Brillet, K.; Journet, L.; Célia, H.; Paulus, L.; Stahl, A.; Pattus, F.; Cobessi, D. A β Strand Lock Exchange for Signal Transduction in TonB-Dependent Transducers on the Basis of a Common Structural Motif. Structure 2007, 15, 1383–1391. [Google Scholar] [CrossRef] [PubMed]
- De Chial, M.; Ghysels, B.; Beatson, S.A.; Geoffroy, V.; Meyer, J.M.; Pattery, T.; Baysse, C.; Chablain, P.; Parsons, Y.N.; Winstanley, C.; et al. Identification of type II and type III pyoverdine receptors from Pseudomonas aeruginosa. Microbiology 2003, 149, 821–831. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ghysels, B.; Dieu, B.T.M.; Beatson, S.A.; Pirnay, J.-P.; Ochsner, U.A.; Vasil, M.L.; Cornelis, P. FpvB, an alternative type I ferripyoverdine receptor of Pseudomonas aeruginosa. Microbiology 2004, 150, 1671–1680. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cobessi, D.; Celia, H.; Folschweiller, N.; Schalk, I.J.; Abdallah, M.A.; Pattus, F. The Crystal Structure of the Pyoverdine Outer Membrane Receptor FpvA from Pseudomonas aeruginosa at 3.6Å Resolution. J. Mol. Biol. 2005, 347, 121–134. [Google Scholar] [CrossRef]
- Wirth, C.; Meyer-Klaucke, W.; Pattus, F.; Cobessi, D. From the Periplasmic Signaling Domain to the Extracellular Face of an Outer Membrane Signal Transducer of Pseudomonas aeruginosa: Crystal Structure of the Ferric Pyoverdine Outer Membrane Receptor. J. Mol. Biol. 2007, 368, 398–406. [Google Scholar] [CrossRef]
- Greenwald, J.; Nader, M.; Celia, H.; Gruffaz, C.; Geoffroy, V.; Meyer, J.-M.; Schalk, I.J.; Pattus, F. FpvA bound to non-cognate pyoverdines: Molecular basis of siderophore recognition by an iron transporter. Mol. Microbiol. 2009, 72, 1246–1259. [Google Scholar] [CrossRef]
- Schalk, I.J.; Abdallah, A.M.A.; Pattus, F. Recycling of Pyoverdin on the FpvA Receptor after Ferric Pyoverdin Uptake and Dissociation in Pseudomonas aeruginosa. Biochemistry 2002, 41, 1663–1671. [Google Scholar] [CrossRef]
- Brillet, K.; Ruffenach, F.; Adams, H.; Journet, L.; Gasser, V.; Hoegy, F.; Guillon, L.; Hannauer, M.; Page, A.; Schalk, I.J. An ABC Transporter with Two Periplasmic Binding Proteins Involved in Iron Acquisition in Pseudomonas aeruginosa. ACS Chem. Biol. 2012, 7, 2036–2045. [Google Scholar] [CrossRef]
- Codd, R. Siderophores and Iron Transport, in Reference Module in Chemistry, Molecular Sciences and Chemical Engineering; Elsevier Inc.: Amsterdam, The Netherlands, 2021. [Google Scholar]
- Hannauer, M.; Braud, A.; Hoegy, F.; Ronot, P.; Boos, A.; Schalk, I.J. The PvdRT-OpmQ efflux pump controls the metal selectivity of the iron uptake pathway mediated by the siderophore pyoverdine in Pseudomonas aeruginosa. Environ. Microbiol. 2011, 14, 1696–1708. [Google Scholar] [CrossRef]
- Braud, A.; Hoegy, F.; Jezequel, K.; Lebeau, T.; Schalk, I.J. New insights into the metal specificity of the Pseudomonas aeruginosa pyoverdine-iron uptake pathway. Environ. Microbiol. 2009, 11, 1079–1091. [Google Scholar] [CrossRef]
- Xiao, R.; Kisaalita, W.S. Iron acquisition from transferrin and lactoferrin by Pseudomonas aeruginosa pyoverdin. Microbiology. 1997, 143, 2509–2515. [Google Scholar] [CrossRef]
- Farvacques, N.; Sonnet, P. Chemistry and Biology of Pyoverdines, Pseudomonas Primary Siderophores. Curr. Med. Chem. 2014, 22, 165–186. [Google Scholar] [CrossRef]
- Banin, E.; Vasil, M.L.; Greenberg, E.P. Iron and Pseudomonas aeruginosa biofilm formation. Proc. Natl. Acad. Sci. USA 2005, 102, 11076–11081. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Escolar, L.; Peérez-Martiín, J.; de Lorenzo, V. Opening the Iron Box: Transcriptional Metalloregulation by the Fur Protein. J. Bacteriol. 1999, 181, 6223–6229. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ochsner, U.A.; Wilderman, P.J.; Vasil, A.I.; Vasil, M.L. GeneChip® expression analysis of the iron starvation response in Pseudomonas aeruginosa: Identification of novel pyoverdine biosynthesis genes. Mol. Microbiol. 2002, 45, 1277–1287. [Google Scholar] [CrossRef] [PubMed]
- Palma, M.; Worgall, S.; Quadri, L.E.N. Transcriptome analysis of the Pseudomonas aeruginosa response to iron. Arch. Microbiol. 2003, 180, 374–379. [Google Scholar] [CrossRef] [PubMed]
- Potvin, E.; Sanschagrin, F.; Levesque, R.C. Sigma factors in Pseudomonas aeruginosa. FEMS Microbiol. Rev. 2008, 32, 38–55. [Google Scholar] [CrossRef] [Green Version]
- Llamas, M.A.; Mooij, M.J.; Sparrius, M.; Vandenbroucke-Grauls, C.M.J.E.; Ratledge, C.; Bitter, W. Characterization of five novel Pseudomonas aeruginosa cell-surface signalling systems. Mol. Microbiol. 2007, 67, 458–472. [Google Scholar] [CrossRef]
- Van Oeffelen, L.; Cornelis, P.; Van Delm, W.; De Ridder, F.; De Moor, B.; Moreau, Y. Detecting cis -regulatory binding sites for cooperatively binding proteins. Nucleic Acids Res. 2008, 36, e46. [Google Scholar] [CrossRef]
- Cobessi, D.; Celia, H.; Pattus, F. Crystal Structure at High Resolution of Ferric-pyochelin and its Membrane Receptor FptA from Pseudomonas aeruginosa. J. Mol. Biol. 2005, 352, 893–904. [Google Scholar] [CrossRef]
- Atkinson, R.A.; El Din, A.L.M.S.; Kieffer, B.; Lefèvre, A.J.-F.; Abdallah, M.A. Bacterial Iron Transport: 1H NMR Determination of the Three-Dimensional Structure of the Gallium Complex of Pyoverdin G4R, the Peptidic Siderophore of Pseudomonas putida G4R. Biochemistry 1998, 37, 15965–15973. [Google Scholar] [CrossRef] [PubMed]
- Takase, H.; Nitanai, H.; Hoshino, K.; Otani, T. Impact of Siderophore Production on Pseudomonas aeruginosa Infections in Immunosuppressed Mice. Infect. Immun. 2000, 68, 1834–1839. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ronnebaum, T.; Lamb, A.L. Nonribosomal peptides for iron acquisition: Pyochelin biosynthesis as a case study. Curr. Opin. Struct. Biol. 2018, 53, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Keating, T.A.; Ehmann, D.E.; Kohli, R.M.; Marshall, C.G.; Trauger, J.W.; Walsh, C.T. Chain termination steps in nonribosomal peptide synthetase assembly lines: Directed acyl-S-enzyme breakdown in antibiotic and siderophore biosynthesis. ChemBioChem 2001, 2, 99–107. [Google Scholar] [CrossRef]
- Reimmann, C.; Patel, H.M.; Serino, L.; Barone, M.; Walsh, C.T.; Haas, D. Essential PchG-Dependent Reduction in Pyochelin Biosynthesis of Pseudomonas aeruginosa. J. Bacteriol. 2001, 183, 813–820. [Google Scholar] [CrossRef] [Green Version]
- Serino, L.; Reimmann, C.; Baur, H.; Beyeler, M.; Visca, P.; Haas, D. Structural genes for salicylate biosynthesis from chorismate in Pseudomonas aeruginosa. Mol. Genet. Genom. 1995, 249, 217–228. [Google Scholar] [CrossRef]
- Reimmann, C.; Serino, L.; Beyeler, M.; Haa, D. Dihydroaeruginoic acid synthetase and pyochelin synthetase, products of the pchEF, are induced by extracellular pyochelin in Pseudornonas aeruginosa. Microbiol. 1998, 144, 3135–3148. [Google Scholar] [CrossRef] [Green Version]
- Gaille, C.; Reimmann, C.; Haas, D. Isochorismate Synthase (PchA), the First and Rate-limiting Enzyme in Salicylate Biosynthesis of Pseudomonas aeruginosa. J. Biol. Chem. 2003, 278, 16893–16898. [Google Scholar] [CrossRef] [Green Version]
- Gaille, C.; Kast, P.; Haas, D. Salicylate biosynthesis in Pseudomonas aeruginosa. Purification and characterization of PchB, a novel bifunctional enzyme displaying isochorismate pyruvate-lyase and chorismate mutase activities. J. Biol Chem. 2002, 277, 21768–21775. [Google Scholar] [CrossRef] [Green Version]
- Quadri, L.E.N.; Keating, T.A.; Patel, H.M.; Walsh, C.T. Assembly of the Pseudomonas aeruginosa Nonribosomal Peptide Siderophore Pyochelin: In Vitro Reconstitution of Aryl-4,2-bisthiazoline Synthetase Activity from PchD, PchE, and PchF. Biochemistry 1999, 38, 14941–14954. [Google Scholar] [CrossRef]
- Patel, H.M.; Tao, J.; Walsh, C.T. Epimerization of an l-Cysteinyl to a d-Cysteinyl Residue during Thiazoline Ring Formation in Siderophore Chain Elongation by Pyochelin Synthetase from Pseudomonas aeruginosa. Biochemistry 2003, 42, 10514–10527. [Google Scholar] [CrossRef] [PubMed]
- Cunrath, O.; Gasser, V.; Hoegy, F.; Reimmann, C.; Guillon, L.; Schalk, I.J. A cell biological view of the siderophore pyochelin iron uptake pathway in Pseudomonas aeruginosa. Environ. Microbiol. 2014, 17, 171–185. [Google Scholar] [CrossRef] [PubMed]
- Roche, B.; Garcia-Rivera, M.A.; Normant, V.; Kuhn, L.; Hammann, P.; Brönstrup, M.; Mislin, G.L.A.; Schalk, I.J. A role for PchHI as the ABC transporter in iron acquisition by the siderophore pyochelin in Pseudomonas aeruginosa. Environ. Microbiol. 2021, 24, 866–877. [Google Scholar] [CrossRef] [PubMed]
- Reimmann, C. Inner-membrane transporters for the siderophores pyochelin in Pseudomonas aeruginosa and enantio-pyochelin in Pseudomonas fluorescens display different enantioselectivities. Microbiology 2012, 158, 1317–1324. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Braud, A.; Hannauer, M.; Mislin, G.L.A.; Schalk, I.J. The Pseudomonas aeruginosa Pyochelin-Iron Uptake Pathway and Its Metal Specificity. J. Bacteriol. 2009, 191, 3517–3525. [Google Scholar] [CrossRef] [Green Version]
- Namiranian, S.; Richardson, D.J.; Russell, D.A.; Sodeau, J.R. Excited State Properties of the Siderophore Pyochelin and its Complex with Zinc Ions. Photochem. Photobiol. 1997, 65, 777–782. [Google Scholar] [CrossRef]
- Yang, B.; Hoegy, F.; Mislin, G.L.; Mesini, P.J.; Schalk, I.J. Terbium, a fluorescent probe for investigation of siderophore pyochelin interactions with its outer membrane transporter FptA. J. Inorg. Biochem. 2011, 105, 1293–1298. [Google Scholar] [CrossRef]
- Braud, A.; Geoffroy, V.; Hoegy, F.; Mislin, G.L.A.; Schalk, I.J. Presence of the siderophores pyoverdine and pyochelin in the extracellular medium reduces toxic metal accumulation in Pseudomonas aeruginosa and increases bacterial metal tolerance. Environ. Microbiol. Rep. 2010, 2, 419–425. [Google Scholar] [CrossRef]
- Llamas, M.A.; Imperi, F.; Visca, P.; Lamont, I.L. Cell-surface signaling in Pseudomonas: Stress responses, iron transport, and pathogenicity. FEMS Microbiol. Rev. 2014, 38, 569–597. [Google Scholar] [CrossRef] [Green Version]
- Michel, L.; González, N.; Jagdeep, S.; Nguyen-Ngoc, T.; Reimmann, C. PchR-box recognition by the AraC-type regulator PchR of Pseudomonas aeruginosa requires the siderophore pyochelin as an effector. Mol. Microbiol. 2005, 58, 495–509. [Google Scholar] [CrossRef]
- Michel, L.; Bachelard, A.; Reimmann, C. Ferripyochelin uptake genes are involved in pyochelin-mediated signalling in Pseudomonas aeruginosa. Microbiology 2007, 153, 1508–1518. [Google Scholar] [CrossRef] [PubMed]
- Raymond, K.N.; Dertz, E.A.; Kim, S.S. Enterobactin: An archetype for microbial iron transport. Proc. Natl. Acad. Sci. USA 2003, 100, 3584–3588. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wyckoff, E.E.; Mey, A.R.; Payne, S.M. Iron acquisition in Vibrio cholerae. BioMetals 2007, 20, 405–416. [Google Scholar] [CrossRef] [PubMed]
- Perraud, Q.; Cantero, P.; Roche, B.; Gasser, V.; Normant, V.P.; Kuhn, L.; Hammann, P.; Mislin, G.L.A.; Ehret-Sabatier, L.; Schalk, I.J. Phenotypic Adaption of Pseudomonas aeruginosa by Hacking Siderophores Produced by Other Microorganisms. Mol. Cell. Proteom. 2020, 19, 589–607. [Google Scholar] [CrossRef] [Green Version]
- Schalk, I.J.; Cunrath, O. An overview of the biological metal uptake pathways in Pseudomonas aeruginosa. Environ. Microbiol. 2016, 18, 3227–3246. [Google Scholar] [CrossRef]
- Gonzalez, M.R.; Ducret, V.; Leoni, S.; Perron, K. Pseudomonas aeruginosa zinc homeostasis: Key issues for an opportunistic pathogen. Biochim. Biophys. Acta 2019, 1862, 722–733. [Google Scholar] [CrossRef]
- Fajardo, A.; Hernando-Amado, S.; Oliver, A.; Ball, G.; Filloux, A.; Martinez, J.L. Characterization of a novel Zn2+-dependent intrinsic imipenemase from Pseudomonas aeruginosa. J. Antimicrob. Chemother. 2014, 69, 2972–2978. [Google Scholar] [CrossRef]
- Mastropasqua, M.C.; D’Orazio, M.; Cerasi, M.; Pacello, F.; Gismondi, A.; Canini, A.; Canuti, L.; Consalvo, A.; Ciavardelli, D.; Chirullo, B.; et al. Growth of Pseudomonas aeruginosa in zinc poor environments is promoted by a nicotianamine-related metallophore. Mol. Microbiol. 2017, 106, 543–561. [Google Scholar] [CrossRef] [Green Version]
- McFarlane, J.S.; Lamb, A.L. Biosynthesis of an Opine Metallophore by Pseudomonas aeruginosa. Biochemistry 2017, 56, 5967–5971. [Google Scholar] [CrossRef]
- Laffont, C.; Brutesco, C.; Hajjar, C.; Cullia, G.; Fanelli, R.; Ouerdane, L.; Cavelier, F.; Arnoux, P. Simple rules govern the diversity of bacterial nicotianamine-like metallophores. Biochem. J. 2019, 476, 2221–2233. [Google Scholar] [CrossRef]
- Pederick, V.G.; Eijkelkamp, B.; Begg, S.N.; Ween, M.; McAllister, L.J.; Paton, J.C.; McDevitt, C.A. ZnuA and zinc homeostasis in Pseudomonas aeruginosa. Sci. Rep. 2015, 5, 13139. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gi, M.; Lee, K.-M.; Kim, S.C.; Yoon, J.-H.; Yoon, S.S.; Choi, J.Y. A novel siderophore system is essential for the growth of Pseudomonas aeruginosa in airway mucus. Sci. Rep. 2015, 5, 14644. [Google Scholar] [CrossRef]
- Zhang, Y.; Rodionov, A.D.; Gelfand, M.S.; Gladyshev, V.N. Comparative genomic analyses of nickel, cobalt and vitamin B12 utilization. BMC Genom. 2009, 10, 78. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crespo, A.; Pedraz, L.; Astola, J.; Torrents, E. Pseudomonas aeruginosa Exhibits Deficient Biofilm Formation in the Absence of Class II and III Ribonucleotide Reductases Due to Hindered Anaerobic Growth. Front. Microbiol. 2016, 7, 688. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, J.; Zhao, T.; Yang, R.; Siridechakorn, I.; Wang, S.; Guo, Q.; Bai, Y.; Shen, H.C.; Lei, X. De novo synthesis, structural assignment and biological evaluation of pseudopaline, a metallophore produced by Pseudomonas aeruginosa. Chem. Sci. 2019, 10, 6635–6641. [Google Scholar] [CrossRef] [Green Version]
- Gomez, N.O.; Tetard, A.; Ouerdane, L.; Laffont, C.; Brutesco, C.; Ball, G.; Lobinski, R.; Denis, Y.; Plésiat, P.; Llanes, C.; et al. Involvement of the Pseudomonas aeruginosa MexAB–OprM efflux pump in the secretion of the metallophore pseudopaline. Mol. Microbiol. 2020, 115, 84–98. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Strains | Pyoverdines Type | Peptide Sequences | Ref |
|---|---|---|---|---|
| P. aeruginosa | ATCC15692(PAO1) | PVDI | Ser–Arg–Ser–FoOHOrn–[Lys–FoOHOrn–Thr–Thr] | [48] |
| P. aeruginosa | ATCC27853 | PVDII | Ser–FoOHOrn–Orn–Gly–Thr–Ser–cOHOrn | [49] |
| P. aeruginosa | Pa6 (R) (sv. III-l) | PVDIII-1 | Ser–Dab–FoOHOrn–Gln–Gln–FoOHOrn–Gly | [50] |
| P. aeruginosa | R’ (sv. III-2) | PVDIII-2 | Ser–Dab–FoOHOrn–Gln–FoOHOrn–Gly | [51] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ghssein, G.; Ezzeddine, Z. A Review of Pseudomonas aeruginosa Metallophores: Pyoverdine, Pyochelin and Pseudopaline. Biology 2022, 11, 1711. https://doi.org/10.3390/biology11121711
Ghssein G, Ezzeddine Z. A Review of Pseudomonas aeruginosa Metallophores: Pyoverdine, Pyochelin and Pseudopaline. Biology. 2022; 11(12):1711. https://doi.org/10.3390/biology11121711
Chicago/Turabian StyleGhssein, Ghassan, and Zeinab Ezzeddine. 2022. "A Review of Pseudomonas aeruginosa Metallophores: Pyoverdine, Pyochelin and Pseudopaline" Biology 11, no. 12: 1711. https://doi.org/10.3390/biology11121711
APA StyleGhssein, G., & Ezzeddine, Z. (2022). A Review of Pseudomonas aeruginosa Metallophores: Pyoverdine, Pyochelin and Pseudopaline. Biology, 11(12), 1711. https://doi.org/10.3390/biology11121711