
Chronic Inflammation Modulates Opioid Receptor Gene Expression and Triggers Respiratory Burst in a Teleost Model




Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
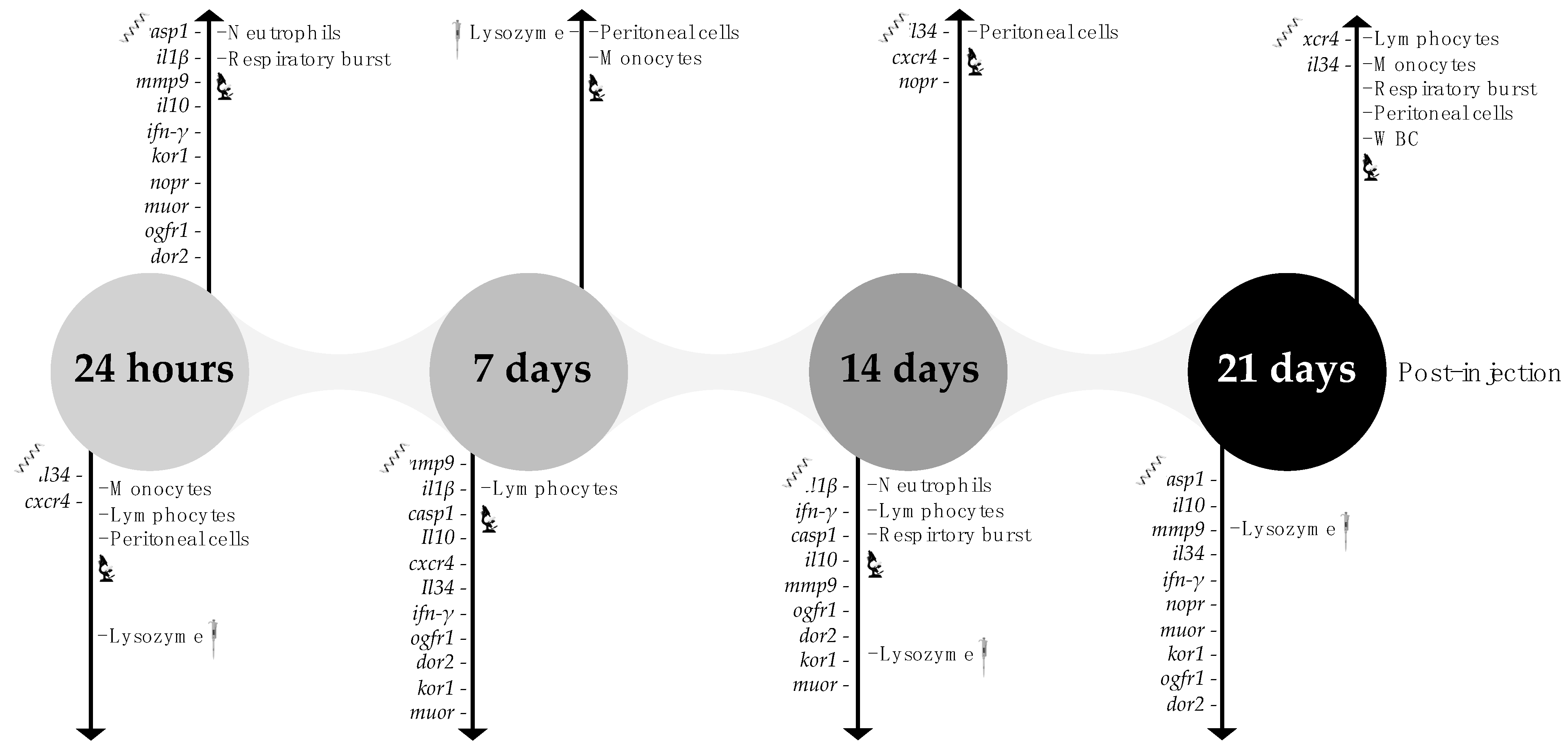
2.1. Experimental Design
2.2. Peritoneal Exudates
2.3. Hematological Parameters
2.4. Respiratory Burst
2.5. Plasma Innate Immune Parameters
2.5.1. Antiprotease Activity
2.5.2. Protease Activity
2.5.3. Lysozyme Concentration
2.5.4. Peroxidase Activity
2.5.5. Immunoglobulin M
2.6. Gene Expression Analysis
2.7. Data Analysis
3. Results
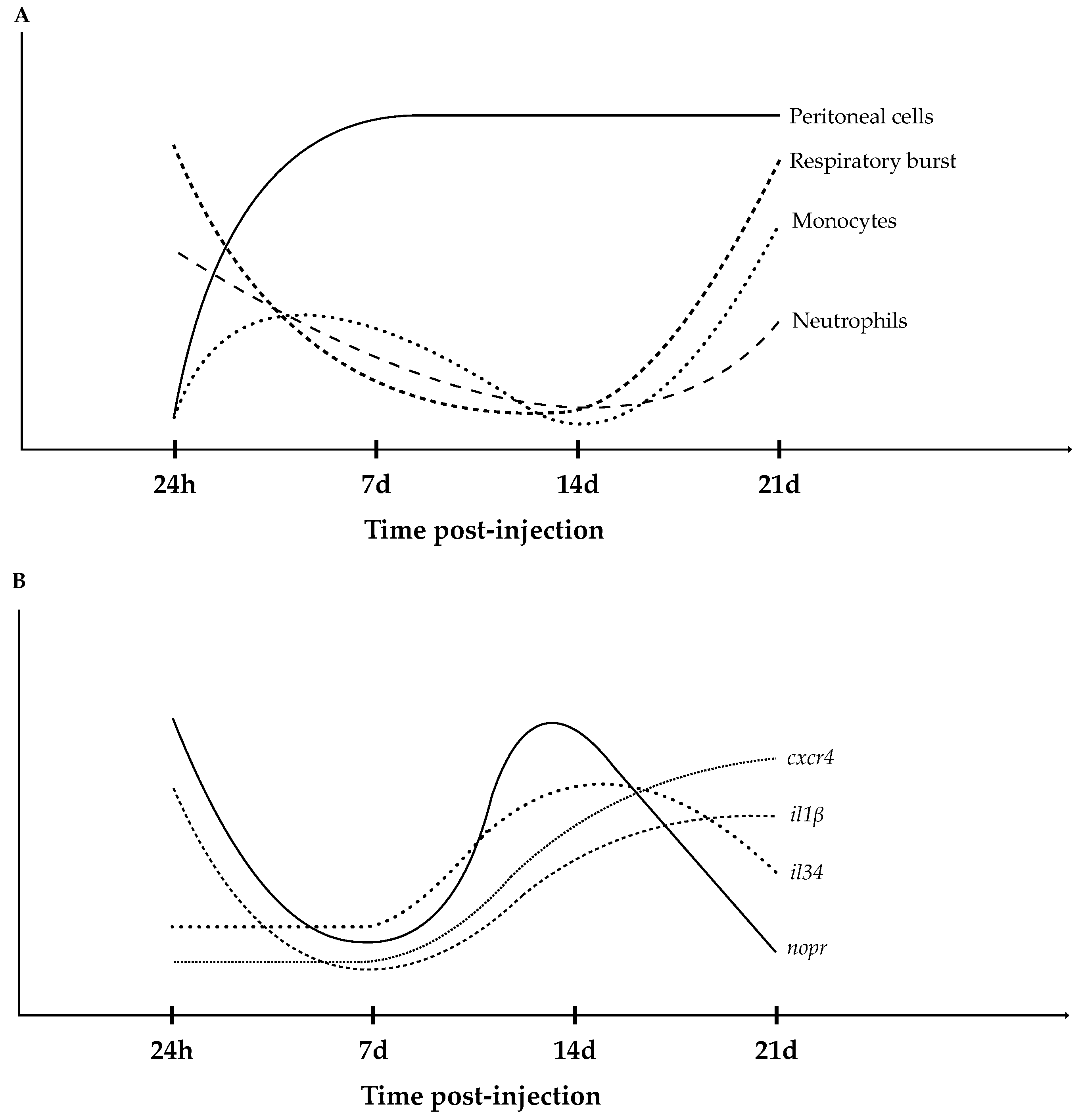
3.1. Hematological Response
3.2. Total Peritoneal Cell Counts and Respiratory Burst of Circulating Leukocytes
3.3. Innate Humoral Parameters
3.4. Head-Kidney Gene Expression
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Matos, E.; Gonçalves, A.; Nunes, M.L.; Dinis, M.T.; Dias, J. Effect of harvesting stress and slaughter conditions on selected flesh quality criteria of gilthead seabream (Sparus aurata). Aquaculture 2010, 305, 66–72. [Google Scholar] [CrossRef]
- Poli, B.M.; Parisi, G.; Scappini, F.; Zampacavallo, G. Fish welfare and quality as affected by pre-slaughter and slaughter management. Aquac. Int. 2005, 13, 29–49. [Google Scholar] [CrossRef]
- Costas, B.; Aragão, C.; Dias, J.; Afonso, A.; Conceição, L.E.C. Interactive effects of a high-quality protein diet and high stocking density on the stress response and some innate immune parameters of Senegalese sole Solea senegalensis. Fish Physiol. Biochem. 2013, 39, 1141–1151. [Google Scholar] [CrossRef] [PubMed]
- Acerete, L.; Reig, L.; Alvarez, D.; Flos, R.; Tort, L. Comparison of two stunning/slaughtering methods on stress response and quality indicators of European sea bass (Dicentrarchus labrax). Aquaculture 2009, 287, 139–144. [Google Scholar] [CrossRef]
- Engelsma, M.; Hougee, S.; Nap, D.; Hofenk, M.; Rombout, J.; van Muiswinkel, W.; Verburg-van Kemenade, B. Multiple acute temperature stress affects leucocyte populations and antibody responses in common carp, Cyprinus carpio L. Fish Shellfish Immunol. 2003, 15, 397–410. [Google Scholar] [CrossRef]
- Ribas, L.; Flos, R.; Reig, L.; MacKenzie, S.; Barton, B.; Tort, L. Comparison of methods for anaesthetizing Senegal sole (Solea senegalensis) before slaughter: Stress responses and final product quality. Aquaculture 2007, 269, 250–258. [Google Scholar] [CrossRef]
- Sadoul, B.; Vijayan, M.M. Stress and Growth. In Biology of Stress in Fish Fish Physiology; Schreck, C., Tort, L., Farrell, A., Brauner, C., Eds.; Fish Physiology; Academic Press: Cambridge, MA, USA, 2016; Volume 35. [Google Scholar]
- Terova, G.; Gornati, R.; Rimoldi, S.; Bernardini, G.; Saroglia, M. Quantification of a glucocorticoid receptor in sea bass (Dicentrarchus labrax, L.) reared at high stocking density. Gene 2005, 357, 144–151. [Google Scholar] [CrossRef] [PubMed]
- Tort, L. Stress and immune modulation in fish. Dev. Comp. Immunol. 2011, 35, 1366–1375. [Google Scholar] [CrossRef]
- Verburg-van Kemenade, B.M.L.; Stolte, E.H.; Metz, J.R.; Chadzinska, M. Neuroendocrine–Immune Interactions in Teleost Fish. In Fish Neuroendocrinology; Bernier, N., Van Der Kraak, G., Farrel, A., Brauner, C., Eds.; Fish Physiology; Academic Press: Cambridge, MA, USA, 2009; Volume 28, pp. 313–364. [Google Scholar]
- Dezfuli, B.; Lui, A.; Boldrini, P.; Pironi, F.; Giari, L. The inflammatory response of fish to helminth parasites. Parasite 2008, 15, 426–433. [Google Scholar] [CrossRef] [Green Version]
- Sharkey, K. Substance P and Calcitonin Gene-Related Peptide (CGRP) in Gastrointestinal Inflammation. Ann. N. Y. Acad. Sci. 1992, 664, 425–442. [Google Scholar] [CrossRef]
- Suzuki, Y.; Iida, T. Fish granulocytes in the process of inflammation. Annu. Rev. Fish Dis. 1992, 2, 149–160. [Google Scholar] [CrossRef]
- Azeredo, R. Amino Acids as Novel Nutraceutics to Modulate Immune Mechanisms and Increase Disease Resistance in Fish. Ph.D. Thesis, Faculty of Sciences of University of Porto, Porto, Portugal, 2017. [Google Scholar]
- Havixbeck, J.J.; Rieger, A.M.; Wong, M.E.; Hodgkinson, J.W.; Barreda, D.R. Neutrophil contributions to the induction and regulation of the acute inflammatory response in teleost fish. J. Leukoc. Biol. 2016, 99, 241–252. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Finn, J.P.; Nielson, N.O. The inflammatory response of rainbow trout. J. Fish Biol. 1971, 3, 463–478. [Google Scholar] [CrossRef]
- Chadzinska, M.; Savelkoul, H.F.; Kemenade, B.L.V.-V. Morphine affects the inflammatory response in carp by impairment of leukocyte migration. Dev. Comp. Immunol. 2009, 33, 88–96. [Google Scholar] [CrossRef] [PubMed]
- Herrero-Turrión, M. Opioids and Opioid Receptors in Fishes. In Reference Module in Life Sciences; Elsevier: Amsterdam, The Netherlands, 2017. [Google Scholar] [CrossRef]
- Chadzinska, M.; Hermsen, T.; Savelkoul, H.F.; Verburg-van Kemenade, B.M. Cloning of opioid receptors in common carp (Cyprinus carpio L.) and their involvement in regulation of stress and immune response. Brain Behav. Immun. 2009, 23, 257–266. [Google Scholar] [CrossRef]
- Bayne, C.J.; Gerwick, L. The acute phase response and innate immunity of fish. Dev. Comp. Immunol. 2001, 25, 725–743. [Google Scholar] [CrossRef]
- Azeredo, R.; Machado, M.; Afonso, A.; Fierro-Castro, C.; Reyes-López, F.E.; Tort, L.; Gesto, M.; Conde-Sieira, M.; Míguez, J.M.; Soengas, J.; et al. Neuroendocrine and Immune Responses Undertake Different Fates following Tryptophan or Methionine Dietary Treatment: Tales from a Teleost Model. Front. Immunol. 2017, 8, 1226. [Google Scholar] [CrossRef] [Green Version]
- Afonso, A.; Ellis, A.E.; Silva, M.T. The leucocyte population of the unstimulated peritoneal cavity of rainbow trout (Oncorhynchus mykiss). Fish Shellfish Immunol. 1997, 7, 335–348. [Google Scholar] [CrossRef]
- Machado, M.; Azeredo, R.; Díaz-Rosales, P.; Afonso, A.; Peres, H.; Oliva-Teles, A.; Costas, B. Dietary tryptophan and methionine as modulators of European seabass (Dicentrarchus labrax) immune status and inflammatory response. Fish Shellfish Immunol. 2015, 42, 353–362. [Google Scholar] [CrossRef] [Green Version]
- Afonso, A.; Lousada, S.; Silva, J.; Ellis, A.E.; Silva, M.T. Neutrophil and macrophage responses to inflammation in the peritoneal cavity of rainbow trout Oncorhynchus mykiss. A light and electron microscopic cytochemical study. Dis. Aquat. Org. 1998, 34, 27–37. [Google Scholar] [CrossRef] [Green Version]
- Nikoskelainen, S.; Ouwehand, A.; Bylund, G.; Salminen, S.; Lilius, E.-M. Immune enhancement in rainbow trout (Oncorhynchus mykiss) by potential probiotic bacteria (Lactobacillus rhamnosus). Fish Shellfish Immunol. 2003, 15, 443–452. [Google Scholar] [CrossRef]
- Ellis, A. Serum antiproteases in fish. In Techiniques in Fish Immunology; Stolen, F.T., Andrerson, D.P., Roberson, B.S., Van Muiswinkel, W.B., Eds.; SOS, Warehouse: Fair Haven, NJ, USA, 1990. [Google Scholar]
- Ellis, A. Lysozyme assays. In Techiniques in Fish Immunology; Stolen, F.T., Andrerson, D.P., Roberson, B.S., Van Muiswinkel, W.B., Eds.; SOS, Warehouse: Fair Haven, NJ, USA, 1990. [Google Scholar]
- Quade, M.J.; Roth, J.A. A rapid, direct assay to measure degranulation of bovine neutrophil primary granules. Vet. Immunol. Immunopathol. 1997, 58, 239–248. [Google Scholar] [CrossRef]
- Cuesta, A.; Meseguer, J.; Esteban, M. Total serum immunoglobulin M levels are affected by immunomodulators in seabream (Sparus aurata L.). Vet. Immunol. Immunopathol. 2004, 101, 203–210. [Google Scholar] [CrossRef] [PubMed]
- Azeredo, R.; Machado, M.; Guardiola, F.A.; Cerezuela, R.; Afonso, A.; Peres, H.; Oliva-Teles, A.; Esteban, M.; Costas, B. Local immune response of two mucosal surfaces of the European seabass, Dicentrarchus labrax, fed tryptophan- or methionine-supplemented diets. Fish Shellfish Immunol. 2017, 70, 76–86. [Google Scholar] [CrossRef]
- Machado, M.; Azeredo, R.; Domingues, A.; Boo, S.F.; Dias, J.; Conceicao, L.; Costas, B. Dietary tryptophan deficiency and its supplementation compromises inflammatory mechanisms and disease resistance in a teleost fish. Sci. Rep. 2019, 9, 7689. [Google Scholar] [CrossRef]
- Rieger, A.M.; Konowalchuk, J.D.; Grayfer, L.; Katzenback, B.; Havixbeck, J.J.; Kiemele, M.D.; Belosevic, M.; Barreda, D.R. Fish and Mammalian Phagocytes Differentially Regulate Pro-Inflammatory and Homeostatic Responses In Vivo. PLoS ONE 2012, 7, e47070. [Google Scholar] [CrossRef]
- Griffin, B. Random and directed migration of trout (Salmo gairdneri) leukocytes: Activation by antibody, complement, and normal serum components. Dev. Comp. Immunol. 1984, 8, 589–597. [Google Scholar] [CrossRef]
- do Vale, A.; Afonso, A.; Silva, M.T. The professional phagocytes of sea bass (Dicentrarchus labrax L.): Cytochemical characterisation of neutrophils and macrophages in the normal and inflamed peritoneal cavity. Fish Shellfish Immunol. 2002, 13, 183–198. [Google Scholar] [CrossRef]
- Rossaint, J.; Margraf, A.; Zarbock, A. Role of Platelets in Leukocyte Recruitment and Resolution of Inflammation. Front. Immunol. 2018, 9, 2712. [Google Scholar] [CrossRef]
- Costas, B.; Rêgo, P.C.N.P.; Simões, I.; Marques, J.; Castro-Cunha, M.; Afonso, A. Cellular and humoral immune responses of Senegalese sole, Solea senegalensis (Kaup), following challenge with two Photobacterium damselae subsp. piscicida strains from different geographical origins. J. Fish Dis. 2013, 36, 543–553. [Google Scholar] [CrossRef]
- Ferdous, F.; Scott, T. A comparative examination of thrombocyte/platelet immunity. Immunol. Lett. 2015, 163, 32–39. [Google Scholar] [CrossRef] [PubMed]
- Nagasawa, T.; Nakayasu, C.; Rieger, A.M.; Barreda, D.; Somamoto, T.; Nakao, M. Phagocytosis by Thrombocytes is a Conserved Innate Immune Mechanism in Lower Vertebrates. Front. Immunol. 2014, 5, 445. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stosik, M.; Tokarz-Deptuła, B.; Deptuła, W. Characterisation of thrombocytes in Osteichthyes. J. Vet. Res. 2019, 63, 123–131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klinger, M.H.; Jelkmann, W. Review: Role of Blood Platelets in Infection and Inflammation. J. Interf. Cytokine Res. 2002, 22, 913–922. [Google Scholar] [CrossRef]
- Semple, J.W.; Italiano, J.E., Jr.; Freedman, J. Platelets and the immune continuum. Nat. Rev. Immunol. 2011, 11, 264–274. [Google Scholar] [CrossRef]
- Secombes, C.J.; Cross, A.R.; Sharp, G.J.; Garcia, R. Nadph oxidase-like activity in rainbow trout Oncorhynchus mykiss (walbaum) macrophages. Dev. Comp. Immunol. 1992, 16, 405–413. [Google Scholar] [CrossRef]
- Plouffe, D.A.; Hanington, P.C.; Walsh, J.G.; Wilson, E.C.; Belosevic, M. Comparison of select innate immune mechanisms of fish and mammals. Xenotransplantation 2005, 12, 266–277. [Google Scholar] [CrossRef]
- Campos-Pérez, J.; Ellis, A.; Secombes, C. Investigation of factors influencing the ability of Renibacterium salmoninarum to stimulate rainbow trout macrophage respiratory burst activity. Fish Shellfish Immunol. 1997, 7, 555–566. [Google Scholar] [CrossRef]
- Lamas, J.; Ellis, A.E. Atlantic salmon (Salmo salar) neutrophil responses to Aeromonas salmonicida. Fish Shellfish Immunol. 1994, 4, 201–219. [Google Scholar] [CrossRef]
- Saurabh, S.; Sahoo, P.K. Lysozyme: An important defence molecule of fish innate immune system. Aquac. Res. 2008, 39, 223–239. [Google Scholar] [CrossRef]
- Murphy, A.E.; Stokesbury, M.J.W.; Easy, R.H. Exploring epidermal mucus protease activity as an indicator of stress in Atlantic sturgeon (Acipenser oxyrinchus oxyrhinchus). J. Fish Biol. 2020, 97, 1354–1362. [Google Scholar] [CrossRef] [PubMed]
- Esteban, M.Á. An Overview of the Immunological Defenses in Fish Skin. ISRN Immunol. 2012, 2012, 853470. [Google Scholar] [CrossRef] [Green Version]
- Ahmed, A.U.; Williams, B.R.G.; Hannigan, G.E. Transcriptional Activation of Inflammatory Genes: Mechanistic Insight into Selectivity and Diversity. Biomolecules 2015, 5, 3087–3111. [Google Scholar] [CrossRef]
- Reis, M.I.R.; Vale, A.D.; Pereira, P.J.B.; Azevedo, J.E.; dos Santos, N.M.S. Caspase-1 and IL-1β Processing in a Teleost Fish. PLoS ONE 2012, 7, e50450. [Google Scholar] [CrossRef] [Green Version]
- Weber, A.; Wasiliew, P.; Kracht, M. Interleukin-1 (IL-1) Pathway. Sci. Signal. 2010, 3, cm1. [Google Scholar] [CrossRef] [PubMed]
- Zou, J.; Secombes, C.J. The Function of Fish Cytokines. Biology 2016, 5, 23. [Google Scholar] [CrossRef]
- Machado, M.; Azeredo, R.; Fontinha, F.; Fernández-Boo, S.; Conceicao, L.; Dias, J.; Costas, B. Dietary Methionine Improves the European Seabass (Dicentrarchus labrax) Immune Status, Inflammatory Response, and Disease Resistance. Front. Immunol. 2018, 9, 2672. [Google Scholar] [CrossRef] [Green Version]
- Kubiczkova, L.; Sedlarikova, L.; Hajek, R.; Sevcikova, S. TGF-β—An excellent servant but a bad master. J. Transl. Med. 2012, 10, 183. [Google Scholar] [CrossRef] [Green Version]
- Reyes-Cerpa, S.; Maisey, K.; Reyes-Lpez, F.; Toro-Ascuy, D.; Mara, A.; Imarai, M. Fish Cytokines and Immune Response. In New Advances and Contributions to Fish Biology; Türker, H., Ed.; IntechOpen: London, UK, 2012. [Google Scholar] [CrossRef]
- Pinto, R.D.; Nascimento, D.S.; Reis, M.I.; do Vale, A.D.; dos Santos, N.M. Molecular characterization, 3D modelling and expression analysis of sea bass (Dicentrarchus labrax L.) interleukin-10. Mol. Immunol. 2007, 44, 2056–2065. [Google Scholar] [CrossRef]
- Tafalla, C.; Coll, J.; Secombes, C. Expression of genes related to the early immune response in rainbow trout (Oncorhynchus mykiss) after viral haemorrhagic septicemia virus (VHSV) infection. Dev. Comp. Immunol. 2005, 29, 615–626. [Google Scholar] [CrossRef]
- Wang, T.; Kono, T.; Monte, M.M.; Kuse, H.; Costa, M.M.; Korenaga, H.; Maehr, T.; Husain, M.; Sakai, M.; Secombes, C.J. Identification of IL-34 in teleost fish: Differential expression of rainbow trout IL-34, MCSF1 and MCSF2, ligands of the MCSF receptor. Mol. Immunol. 2013, 53, 398–409. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grayfer, L.; Kerimoglu, B.; Yaparla, A.; Hodgkinson, J.W.; Xie, J.; Belosevic, M. Mechanisms of Fish Macrophage Antimicrobial Immunity. Front. Immunol. 2018, 9, 1105. [Google Scholar] [CrossRef] [PubMed]
- Masteller, E.L.; Wong, B.R. Targeting IL-34 in chronic inflammation. Drug Discov. Today 2014, 19, 1212–1216. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, X.; Kang, L.; Liu, W.; Lou, B.; Wu, C.; Jiang, L. Molecular characterization and expression analysis of the large yellow croaker (Larimichthys crocea) chemokine receptors CXCR2, CXCR3, and CXCR4 after bacterial and poly I:C challenge. Fish Shellfish Immunol. 2017, 70, 228–239. [Google Scholar] [CrossRef] [PubMed]
- Fu, Q.; Yang, Y.; Li, C.; Zeng, Q.; Zhou, T.; Li, N.; Liu, Y.; Liu, S.; Liu, Z. The CC and CXC chemokine receptors in channel catfish (Ictalurus punctatus) and their involvement in disease and hypoxia responses. Dev. Comp. Immunol. 2017, 77, 241–251. [Google Scholar] [CrossRef] [PubMed]
- Isles, H.M.; Herman, K.D.; Robertson, A.L.; Loynes, C.A.; Prince, L.R.; Elks, P.M.; Renshaw, S.A. The CXCL12/CXCR4 Signaling Axis Retains Neutrophils at Inflammatory Sites in Zebrafish. Front. Immunol. 2019, 10, 1784. [Google Scholar] [CrossRef] [Green Version]
- van der Sar, A.M.; Spaink, H.P.; Zakrzewska, A.; Bitter, W.; Meijer, A.H. Specificity of the zebrafish host transcriptome response to acute and chronic mycobacterial infection and the role of innate and adaptive immune components. Mol. Immunol. 2009, 46, 2317–2332. [Google Scholar] [CrossRef]
- Machado, M. Mediators of Inflammation: Unravelling the Role of Methionine and Tryptophan during Infection. Ph.D. Thesis, Univerisity of Porto, Porto, Portugal, 2020. [Google Scholar]
- Sneddon, L.U. Comparative Physiology of Nociception and Pain. Physiology 2018, 33, 63–73. [Google Scholar] [CrossRef]
- Enyedi, B.; Kala, S.; Nikolich-Zugich, T.; Niethammer, P. Tissue damage detection by osmotic surveillance. Nat. Cell Biol. 2013, 15, 1123–1130. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Acronym | GenBank ID | Eff (%) 1 | Ta (°C) | Primer Sequence (5′–3′) |
|---|---|---|---|---|---|
| Elongation factor 1-α | ef1α | AJ866727.1 | 96.45 | 57 | F: AACTTCAACGCCCAGGTCAT R: CTTCTTGCCAGAACGACGGT |
| 40s ribosomal protein | 40s | HE978789.1 | 92.96 | 55 | F: TGATTGTGACAGACCCTCGTG R: CACAGAGCAATGGTGGGGAT |
| ribosomal protein L13 mRNA | L13α | DQ836931.1 | 127.31 | 55 | F: AGTCCGGTGTCCCACTATCA R: GAGTTCCTCCAGGGTGAAGC |
| Interleukin 34 | il34 | DLAgn_00164750 2 | 120.94 | 60 | F: GGAAATACGCTTCAGGGATG R: GGCACTCTGTCGGGTTCTT |
| C-Reactive protein | ptx | EU660933.1 | 114.76 | 60 | F: TGAAGTTTTTGCTGCTGGTG R: GGTTTCTTGTGGGAAGGTGA |
| Chemokine CXC receptor 4 | cxcr4 | FN687464.1 | 93.43 | 60 | F: ACCAGACCTTGTGTTTGCCA R: ATGAAGCCCACCAGGATGTG |
| Interleukin 1 β | il1β | AJ269472.1 | 96.7 | 57 | F: AGCGACATGGTGCGATTTCT R: CTCCTCTGCTGTGCTGATGT |
| Interleukin 10 | il10 | AM268529.1 | 116 | 55 | F: ACCCCGTTCGCTTGCCA R: CATCTGGTGACATCACTC |
| Transforming growth factor-β | tgf β | AM421619.1 | 105.56 | 55 | F: ACCTACATCTGGAACGCTGA R: TGTTGCCTGCCCACATAGTAG |
| Caspase 1 | casp1 | DQ198377.1 | 124 | 55 | F: GTGTTTCAGATGCGGGAGGA R: ATTTAAGTTAACTCACCGGGGG |
| Interferon γ | ifn-γ | FQ310507.3 | 118.38 | 55 | F: GTACAGACAGGCGTCCAAAGCATCA R: CAAACAGGGCAGCCGTCTCATCAA |
| Inmunoglobulin M | IgM | FN908858 | 91.64 | 60 | F: AGGACAGGACTGCTGCTGTT R: CACCTGCTGTCTGCTGTTGT |
| Matrix-metalloproteinase 9 | mmp9 | FN908863.1 | 98.44 | 57 | F: TGTGCCACCACAGACAACTT R: TTCCATCTCCACGTCCCTCA |
| μ opioid receptor | muor | DLAgn_00015310 2 | 99.81 | 60 | F: GTCACCAGCACCCTACCATT R: CGAGGAGAGAATCCAGTTGC |
| κ-type opioid receptor-like 1 | kor1 | DLAgn_00007470 2 | 89.71 | 60 | F: TCTGGTGCTTGTGGTAGTCG R: TGGCAGTCTCTGTGTCCTTG |
| Nociceptin opioid receptor | nopr | DLAgn_00125610 2 | 97.57 | 60 | F: CTCCTTTCTCATCCCTGTGG R: GTTGCGGTCCTTTTCCTTG |
| δ-opioid receptor | dor2 | DLAgn_00062690 2 | 108.06 | 60 | F: CGCTTCTCGGTCTCCATAACT R: GGTCTCATTACTACTTGAAG |
| Opioid growth factor receptor-like protein 1-like | ogfr1 | DLAgn_00128530 2 | 96.79 | 60 | F: GTTGGGAATGGAGATGGAAA R: GCTTCAGATTTTGGCTCAGG |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Peixoto, D.; Machado, M.; Azeredo, R.; Costas, B. Chronic Inflammation Modulates Opioid Receptor Gene Expression and Triggers Respiratory Burst in a Teleost Model. Biology 2022, 11, 764. https://doi.org/10.3390/biology11050764
Peixoto D, Machado M, Azeredo R, Costas B. Chronic Inflammation Modulates Opioid Receptor Gene Expression and Triggers Respiratory Burst in a Teleost Model. Biology. 2022; 11(5):764. https://doi.org/10.3390/biology11050764
Chicago/Turabian StylePeixoto, Diogo, Marina Machado, Rita Azeredo, and Benjamín Costas. 2022. "Chronic Inflammation Modulates Opioid Receptor Gene Expression and Triggers Respiratory Burst in a Teleost Model" Biology 11, no. 5: 764. https://doi.org/10.3390/biology11050764
APA StylePeixoto, D., Machado, M., Azeredo, R., & Costas, B. (2022). Chronic Inflammation Modulates Opioid Receptor Gene Expression and Triggers Respiratory Burst in a Teleost Model. Biology, 11(5), 764. https://doi.org/10.3390/biology11050764