
Maternal Adipocyte Connexin43 Gap Junctions Affect Breastmilk Lactose Levels and Neonate Growth in Mice

 ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
3. Results
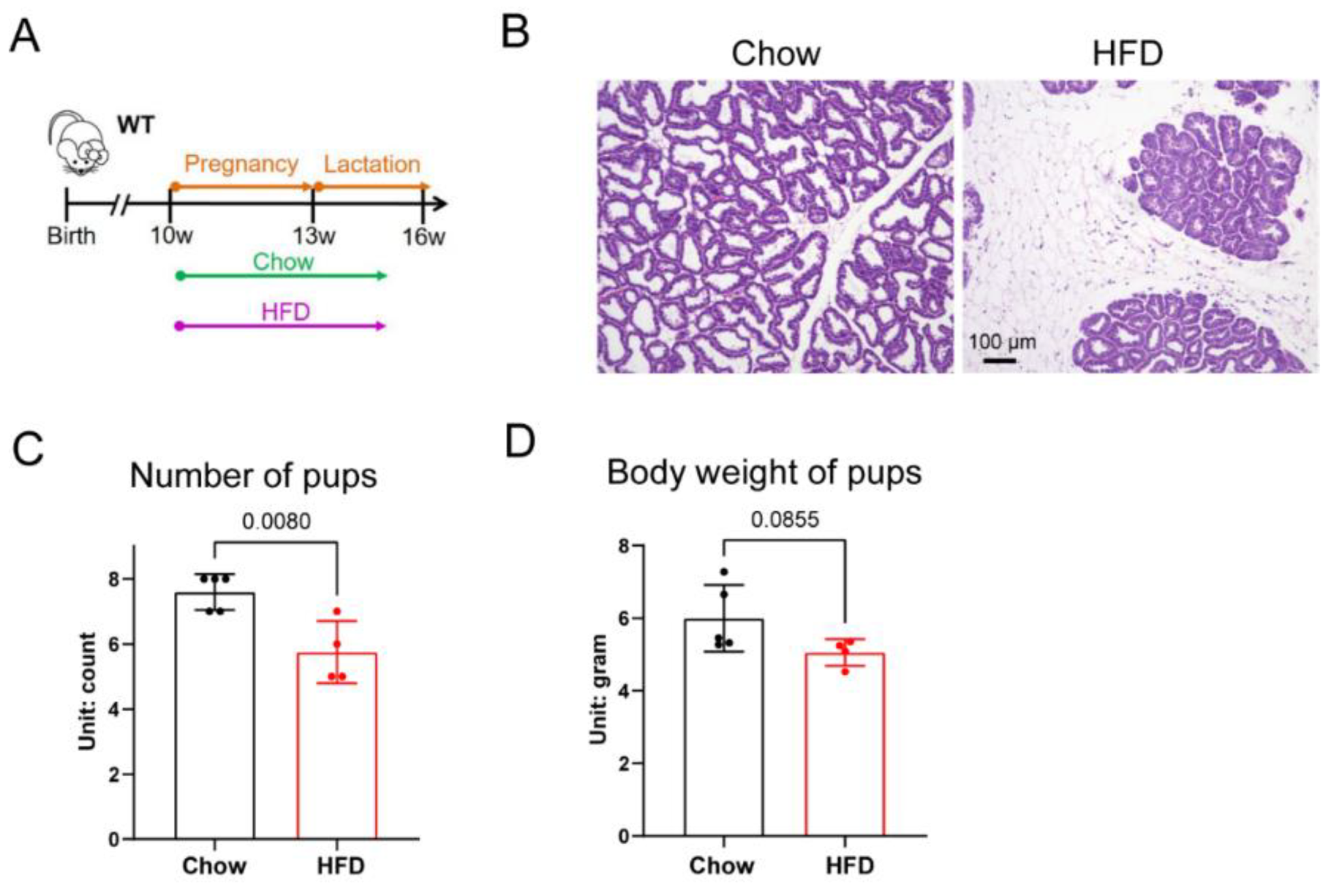
3.1. High-Fat Diet Feeding Impaired Adipocyte Remodeling and Difficulty in Supporting Neonate Growth during Lactation
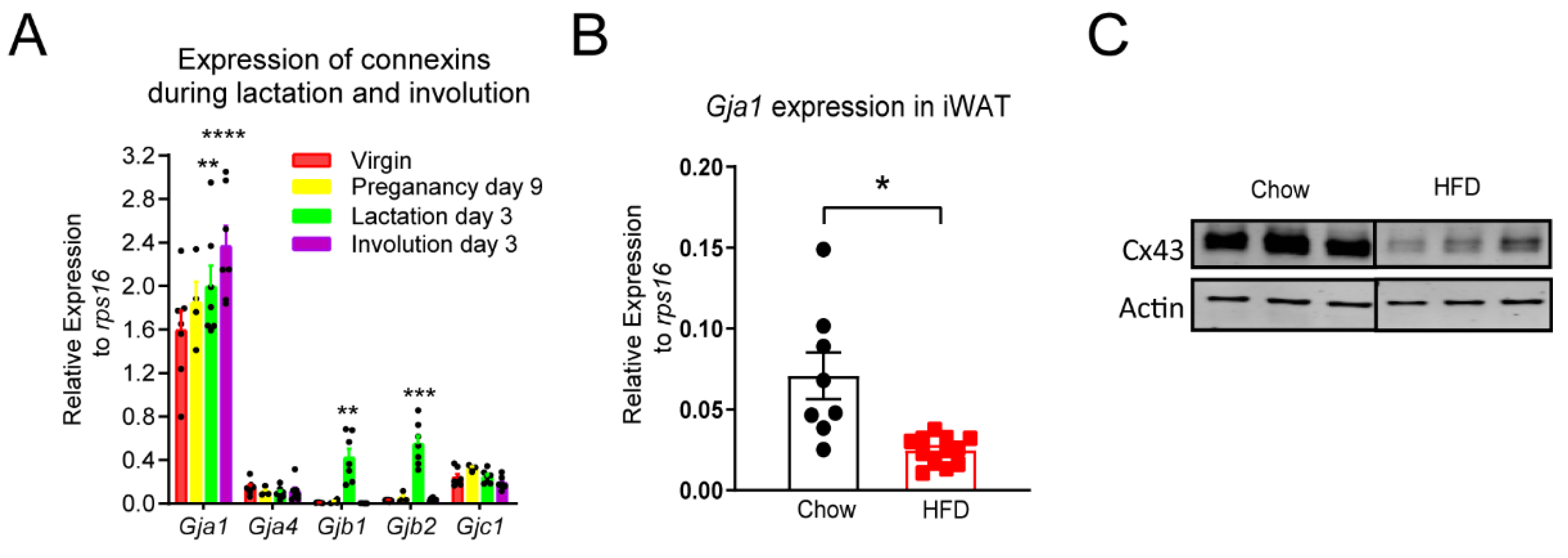
3.2. Gja1 Is the Most Abundant Connexin Isoform in the Mammary Fat Pad and Obese Female Mice Have Reduced Adipose Tissue Gja1 Expression and Cx43 Protein Levels
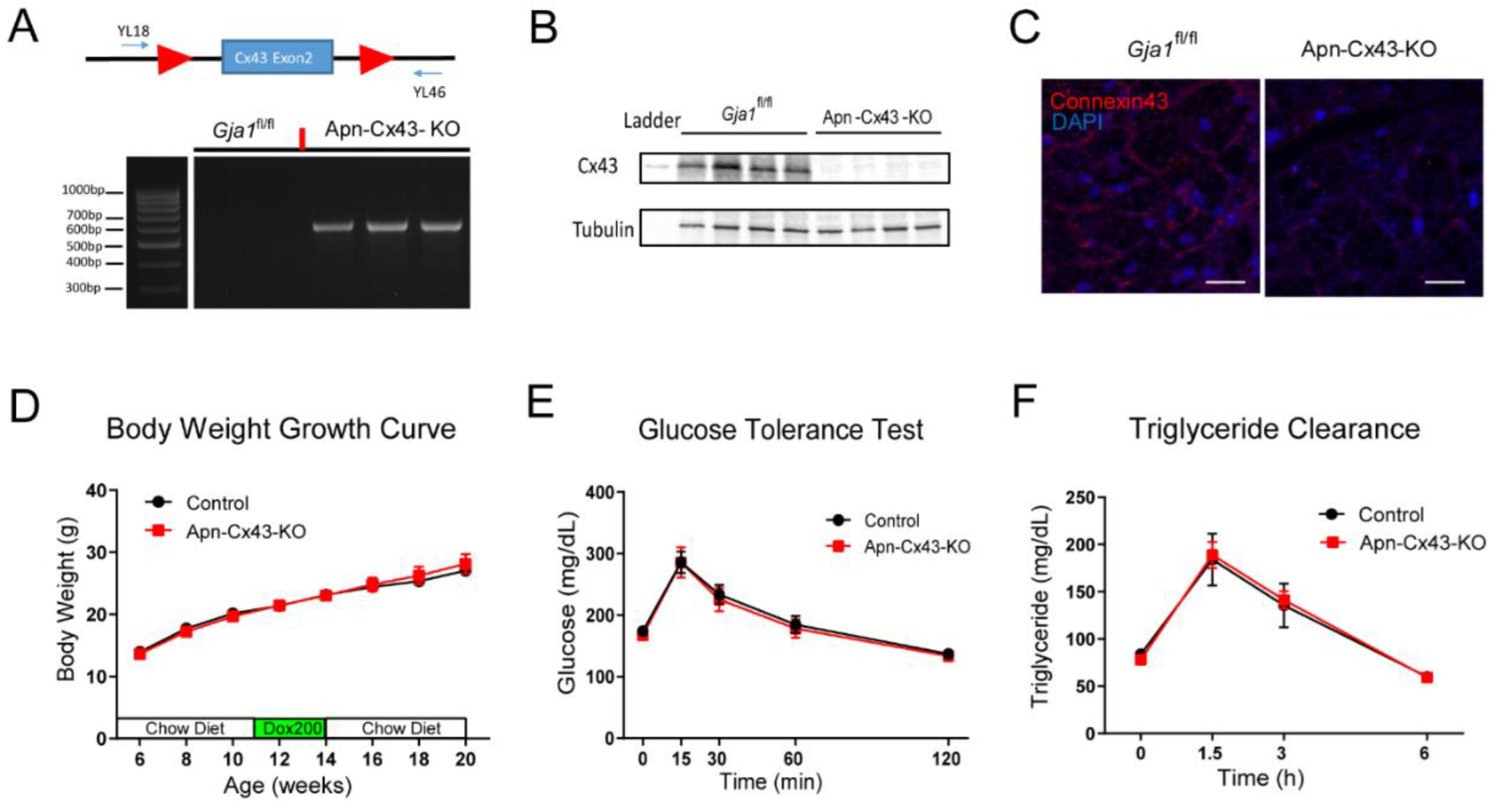
3.3. Adipocyte-Specific Cx43 Deletion Does Not Affect Glucose and Lipid Metabolism in Female Mice
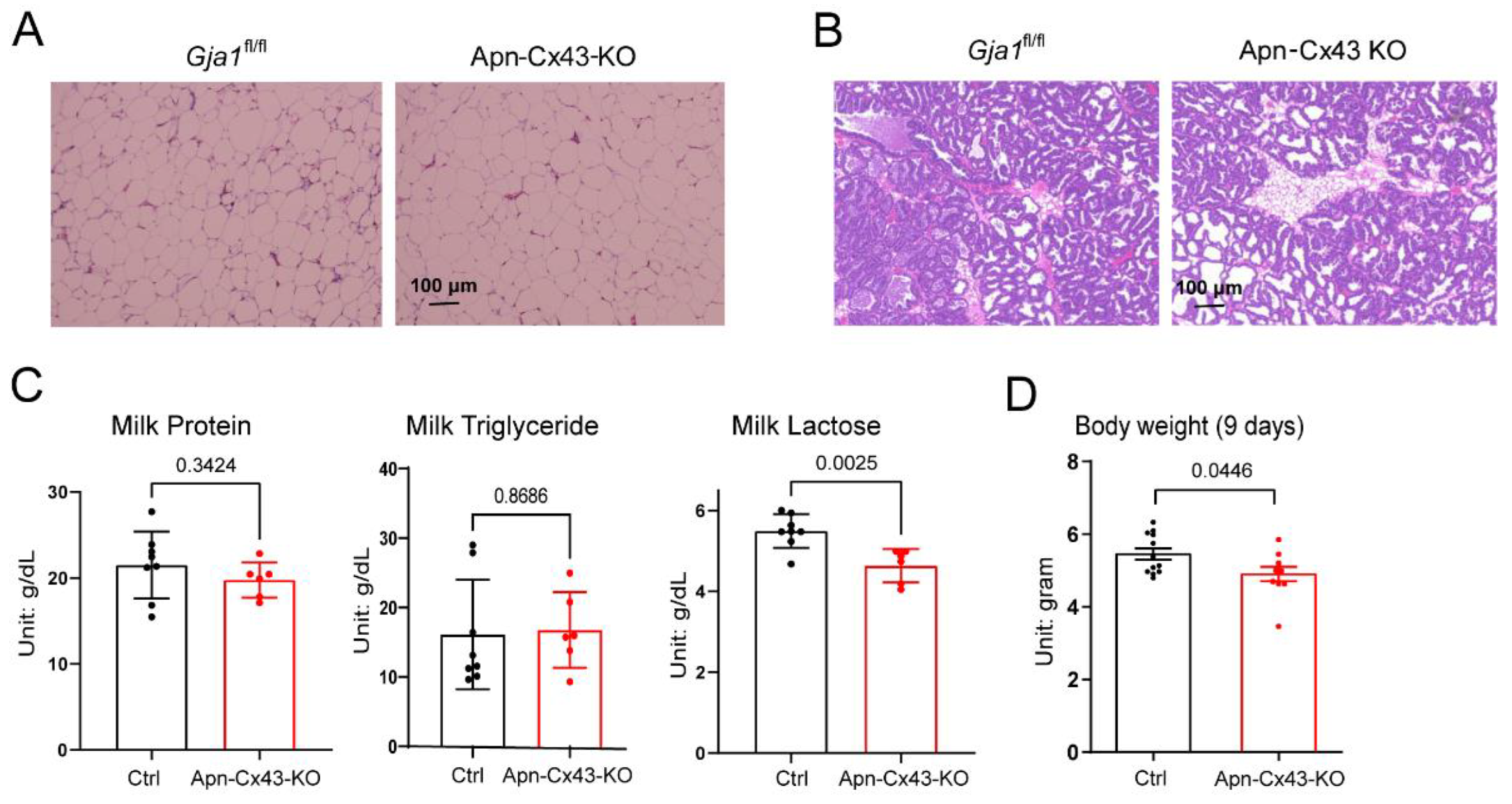
3.4. Adipose Tissue Cx43 Deletion Impairs Adipose Tissue Remodeling and Reduces Breastmilk Lactose Levels during Lactation
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Smith, C.; Patel, R.; Vannabouathong, C.; Sales, B.; Rabinovich, A.; McCormack, R.; Belzile, E.L.; Bhandari, M. Combined intra-articular injection of corticosteroid and hyaluronic acid reduces pain compared to hyaluronic acid alone in the treatment of knee osteoarthritis. Knee Surg. Sports Traumatol. Arthrosc. 2019, 27, 1974–1983. [Google Scholar] [CrossRef] [PubMed]
- Poston, L.; Caleyachetty, R.; Cnattingius, S.; Corvalan, C.; Uauy, R.; Herring, S.; Gillman, M.W. Preconceptional and maternal obesity: Epidemiology and health consequences. Lancet Diabetes Endocrinol. 2016, 4, 1025–1036. [Google Scholar] [CrossRef]
- Li, R.; Jewell, S.; Grummer-Strawn, L. Maternal obesity and breast-feeding practices. Am. J. Clin. Nutr. 2003, 77, 931–936. [Google Scholar] [CrossRef] [Green Version]
- Rutishauser, I.H.; Carlin, J.B. Body mass index and duration of breast feeding: A survival analysis during the first six months of life. J. Epidemiol. Community Health. 1992, 46, 559–565. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hilson, J.A.; Rasmussen, K.M.; Kjolhede, C.L. Maternal obesity and breast-feeding success in a rural population of white women. Am. J. Clin. Nutr. 1997, 66, 1371–1378. [Google Scholar] [CrossRef] [Green Version]
- Scott, J.A.; Binns, C.W.; Oddy, W.H.; Graham, K.I. Predictors of breastfeeding duration: Evidence from a cohort study. Pediatrics 2006, 117, e646–e655. [Google Scholar] [CrossRef] [Green Version]
- Panagos, P.G.; Vishwanathan, R.; Penfield-Cyr, A.; Matthan, N.R.; Shivappa, N.; Wirth, M.D.; Hebert, J.R.; Sen, S. Breastmilk from obese mothers has pro-inflammatory properties and decreased neuroprotective factors. J. Perinatol. 2016, 36, 284–290. [Google Scholar] [CrossRef] [Green Version]
- Mese, G.; Richard, G.; White, T.W. Gap junctions: Basic structure and function. J. Investig. Dermatol. 2007, 127, 2516–2524. [Google Scholar] [CrossRef] [Green Version]
- Goodenough, D.A.; Paul, D.L. Gap junctions. Cold Spring Harb. Perspect. Biol. 2009, 1, a002576. [Google Scholar] [CrossRef]
- Zhu, Y. Gap Junction-Dependent and -Independent Functions of Connexin43 in Biology. Biology 2022, 11, 283. [Google Scholar] [CrossRef]
- Burke, S.; Nagajyothi, F.; Thi, M.M.; Hanani, M.; Scherer, P.E.; Tanowitz, H.B.; Spray, D.C. Adipocytes in both brown and white adipose tissue of adult mice are functionally connected via gap junctions: Implications for Chagas disease. Microbes Infect. 2014, 16, 893–901. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, Y.; Gao, Y.; Tao, C.; Shao, M.; Zhao, S.; Huang, W.; Tao, T.; Johnson, J.A.; Liu, T.; Cypess, A.M.; et al. Connexin 43 Mediates White Adipose Tissue Beiging by Facilitating the Propagation of Sympathetic Neuronal Signals. Cell Metab. 2016, 24, 420–433. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gonzalez-Casanova, J.E.; Duran-Aguero, S.; Caro-Fuentes, N.J.; Gamboa-Arancibia, M.E.; Bruna, T.; Bermudez, V.; Rojas-Gómez, D.M. New Insights on the Role of Connexins and Gap Junctions Channels in Adipose Tissue and Obesity. Int. J. Mol. Sci. 2021, 22, 12145. [Google Scholar] [CrossRef] [PubMed]
- Yanagiya, T.; Tanabe, A.; Hotta, K. Gap-junctional communication is required for mitotic clonal expansion during adipogenesis. Obesity 2007, 15, 572–582. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, Y.; Li, N.; Huang, M.; Chen, X.; An, Y.A.; Li, J.; Zhao, S.; Funcke, J.-B.; Cao, J.; He, Z.; et al. Activating Connexin43 gap junctions primes adipose tissue for therapeutic intervention. Acta Pharm. Sin. B, 2022; in press. [Google Scholar]
- Schneider-Picard, G.; Carpentier, J.L.; Orci, L. Quantitative evaluation of gap junctions during development of the brown adipose tissue. J. Lipid Res. 1980, 21, 600–607. [Google Scholar] [CrossRef]
- Liao, Y.; Day, K.H.; Damon, D.N.; Duling, B.R. Endothelial cell-specific knockout of connexin 43 causes hypotension and bradycardia in mice. Proc. Natl. Acad. Sci. USA 2001, 98, 9989–9994. [Google Scholar] [CrossRef] [Green Version]
- Perl, A.K.; Wert, S.E.; Nagy, A.; Lobe, C.G.; Whitsett, J.A. Early restriction of peripheral and proximal cell lineages during formation of the lung. Proc. Natl. Acad. Sci. USA 2002, 99, 10482–10487. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, K.; Wernstedt Asterholm, I.; Kusminski, C.M.; Bueno, A.C.; Wang, Z.V.; Pollard, J.W.; Brekken, R.A.; Schrer, P.E. Dichotomous effects of VEGF-A on adipose tissue dysfunction. Proc. Natl. Acad. Sci. USA 2012, 109, 5874–5879. [Google Scholar] [CrossRef] [Green Version]
- Gorewit, R.C.; Wachs, E.A.; Sagi, R.; Merrill, W.G. Current concepts on the role of oxytocin in milk ejection. J. Dairy Sci. 1983, 66, 2236–2250. [Google Scholar] [CrossRef]
- DePeters, E.J.; Hovey, R.C. Methods for collecting milk from mice. J. Mammary Gland. Biol. Neoplasia. 2009, 14, 397–400. [Google Scholar] [CrossRef] [Green Version]
- Zhu, Y.; Li, N.; Huang, M.; Bartels, M.; Dogne, S.; Zhao, S.; Chen, X.; Crewe, C.; Straub, L.; Vishanath, L.; et al. Adipose tissue hyaluronan production improves systemic glucose homeostasis and primes adipocytes for CL 316,243-stimulated lipolysis. Nat. Commun. 2021, 12, 4829. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Spandidos, A.; Wang, H.; Seed, B. PrimerBank: A PCR primer database for quantitative gene expression analysis, 2012 update. Nucleic Acids Res. 2012, 40, D1144–D1149. [Google Scholar] [CrossRef] [Green Version]
- Huang, M.; Mathew, N.; Zhu, Y. Assessing Whole-Body Lipid-Handling Capacity in Mice. J. Vis. Exp. 2020. [Google Scholar] [CrossRef] [PubMed]
- Bartick, M.; Reinhold, A. The burden of suboptimal breastfeeding in the United States: A pediatric cost analysis. Pediatrics 2010, 125, e1048–e1056. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Azarnia, R.; Russell, T.R. Cyclic AMP effects on cell-to-cell junctional membrane permeability during adipocyte differentiation of 3T3-L1 fibroblasts. J. Cell Biol. 1985, 100, 265–269. [Google Scholar] [CrossRef] [Green Version]
- Yeganeh, A.; Stelmack, G.L.; Fandrich, R.R.; Halayko, A.J.; Kardami, E.; Zahradka, P. Connexin 43 phosphorylation and degradation are required for adipogenesis. Biochim. Biophys. Acta. 2012, 1823, 1731–1744. [Google Scholar] [CrossRef] [Green Version]
- Yamanouchi, K.; Yada, E.; Ishiguro, N.; Nishihara, M. 18alpha-glycyrrhetinic acid induces phenotypic changes of skeletal muscle cells to enter adipogenesis. Cell Physiol Biochem. 2007, 20, 781–790. [Google Scholar] [CrossRef]
- Schiller, P.C.; D’Ippolito, G.; Brambilla, R.; Roos, B.A.; Howard, G.A. Inhibition of gap-junctional communication induces the trans-differentiation of osteoblasts to an adipocytic phenotype in vitro. J. Biol. Chem. 2001, 276, 14133–14138. [Google Scholar] [CrossRef] [Green Version]
- Zappitelli, T.; Chen, F.; Aubin, J.E. Up-regulation of BMP2/4 signaling increases both osteoblast-specific marker expression and bone marrow adipogenesis in Gja1Jrt/+ stromal cell cultures. Mol. Biol. Cell 2015, 26, 832–842. [Google Scholar] [CrossRef]
- Noyan-Ashraf, M.H.; Shikatani, E.A.; Schuiki, I.; Mukovozov, I.; Wu, J.; Li, R.K.; Volchuk, A.; Robinson, L.A.; Billia, F.; Drucker, D.J.; et al. A glucagon-like peptide-1 analog reverses the molecular pathology and cardiac dysfunction of a mouse model of obesity. Circulation 2013, 127, 74–85. [Google Scholar] [CrossRef] [Green Version]
- Young, B.E.; Patinkin, Z.W.; Pyle, L.; de la Houssaye, B.; Davidson, B.S.; Geraghty, S.; Morrow, A.L.; Krebs, N. Markers of Oxidative Stress in Human Milk do not Differ by Maternal BMI But are Related to Infant Growth Trajectories. Matern Child. Health J. 2017, 21, 1367–1376. [Google Scholar] [CrossRef] [PubMed]
- Leghi, G.E.; Netting, M.J.; Middleton, P.F.; Wlodek, M.E.; Geddes, D.T.; Muhlhausler, A.B.S. The impact of maternal obesity on human milk macronutrient composition: A systematic review and meta-analysis. Nutrients 2020, 12, 934. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, N.; Wang, X.; Li, Q.; Han, B.; Chen, Y.; Zhu, C.; Chen, Y.; Lin, D.; Wang, B.; Jensen, M.D.; et al. The famine exposure in early life and metabolic syndrome in adulthood. Clin. Nutr. 2017, 36, 253–259. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schwarzenberg, S.J.; Georgieff, M.K.; Committee, O.N. Advocacy for Improving Nutrition in the First 1000 Days to Support Childhood Development and Adult Health. Pediatrics 2018, 141, e20173716. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Conklin, C.M.; Bechberger, J.F.; MacFabe, D.; Guthrie, N.; Kurowska, E.M.; Naus, C.C. Genistein and quercetin increase connexin43 and suppress growth of breast cancer cells. Carcinogenesis 2007, 28, 93–100. [Google Scholar] [CrossRef]
- Shen, F.; Huang, W.L.; Xing, B.P.; Fang, X.; Feng, M.; Jiang, C.M. Genistein Improves the Major Depression through Suppressing the Expression of miR-221/222 by Targeting Connexin 43. Psychiatry Investig. 2018, 15, 919–925. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.; Li, L.; Li, Y.; Liang, X.; Sun, Q.; Yu, H.; Zhong, J.; Ni, Y.; Chen, J.; Zhao, Z.; et al. Activation of TRPV1 channel by dietary capsaicin improves visceral fat remodeling through connexin43-mediated Ca2+ Influx. Cardiovasc. Diabetol. 2015, 14, 22. [Google Scholar] [CrossRef] [Green Version]
- Jeong, G.; Park, S.W.; Lee, Y.K.; Ko, S.Y.; Shin, S.M. Maternal food restrictions during breastfeeding. Korean J. Pediatr. 2017, 60, 70–76. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name of the Gene | PCR Product Length | Name of the Primer | Sequence (5′ to 3′) |
|---|---|---|---|
| Gja1 | 158 bp | forward | GTCACTGGTGACAGAAACAA |
| reverse | CTGTCGTCAGGGAAATCAAA | ||
| Gja4 | 220 bp | forward | CCCACATCCGATACTGGGTG |
| reverse | CGAAGACGACCGTCCTCTG | ||
| Gjb1 | 134 bp | forward | GCACGTAGCTCACCAACAG |
| reverse | TGATGACATAGGTCCACCACA | ||
| Gjb2 | 133 bp | forward | ATCCTCGGGGGTGTCAACAA |
| reverse | AGACAAAATCGGCTTGCTCATC | ||
| Gjc1 | 167 bp | forward | CAGAGCCAACCAAAACCTAAGC |
| reverse | CTGCACACATAAAATGGGTGGA | ||
| Rps16 | 127 bp | forward | CACTGCAAACGGGGAAATGG |
| reverse | CACCAGCAAATCGCTCCTTG | ||
| Rps18 | 174 bp | forward | GGAGATATGCTCATGTGGTG |
| reverse | TGTACTTCCCATCCTTCACA |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huang, M.; Song, A.; Chen, X.; Ishtiaq, S.; Wang, C.; Hadsell, D.L.; Wang, Q.A.; Zhu, Y. Maternal Adipocyte Connexin43 Gap Junctions Affect Breastmilk Lactose Levels and Neonate Growth in Mice. Biology 2022, 11, 1023. https://doi.org/10.3390/biology11071023
Huang M, Song A, Chen X, Ishtiaq S, Wang C, Hadsell DL, Wang QA, Zhu Y. Maternal Adipocyte Connexin43 Gap Junctions Affect Breastmilk Lactose Levels and Neonate Growth in Mice. Biology. 2022; 11(7):1023. https://doi.org/10.3390/biology11071023
Chicago/Turabian StyleHuang, Mingyang, Anying Song, Xi Chen, Sarah Ishtiaq, Chunmei Wang, Darryl L. Hadsell, Qiong A. Wang, and Yi Zhu. 2022. "Maternal Adipocyte Connexin43 Gap Junctions Affect Breastmilk Lactose Levels and Neonate Growth in Mice" Biology 11, no. 7: 1023. https://doi.org/10.3390/biology11071023
APA StyleHuang, M., Song, A., Chen, X., Ishtiaq, S., Wang, C., Hadsell, D. L., Wang, Q. A., & Zhu, Y. (2022). Maternal Adipocyte Connexin43 Gap Junctions Affect Breastmilk Lactose Levels and Neonate Growth in Mice. Biology, 11(7), 1023. https://doi.org/10.3390/biology11071023