
Metabolic Scaling in Birds and Mammals: How Taxon Divergence Time, Phylogeny, and Metabolic Rate Affect the Relationship between Scaling Exponents and Intercepts

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. BMR Dataset
2.2. Date of Time of Divergence of Taxa
2.3. Statistical Analysis
2.4. Level of BMR and Dimensionless Ratio of BMR
2.5. Phylogenetic Analysis
3. Results
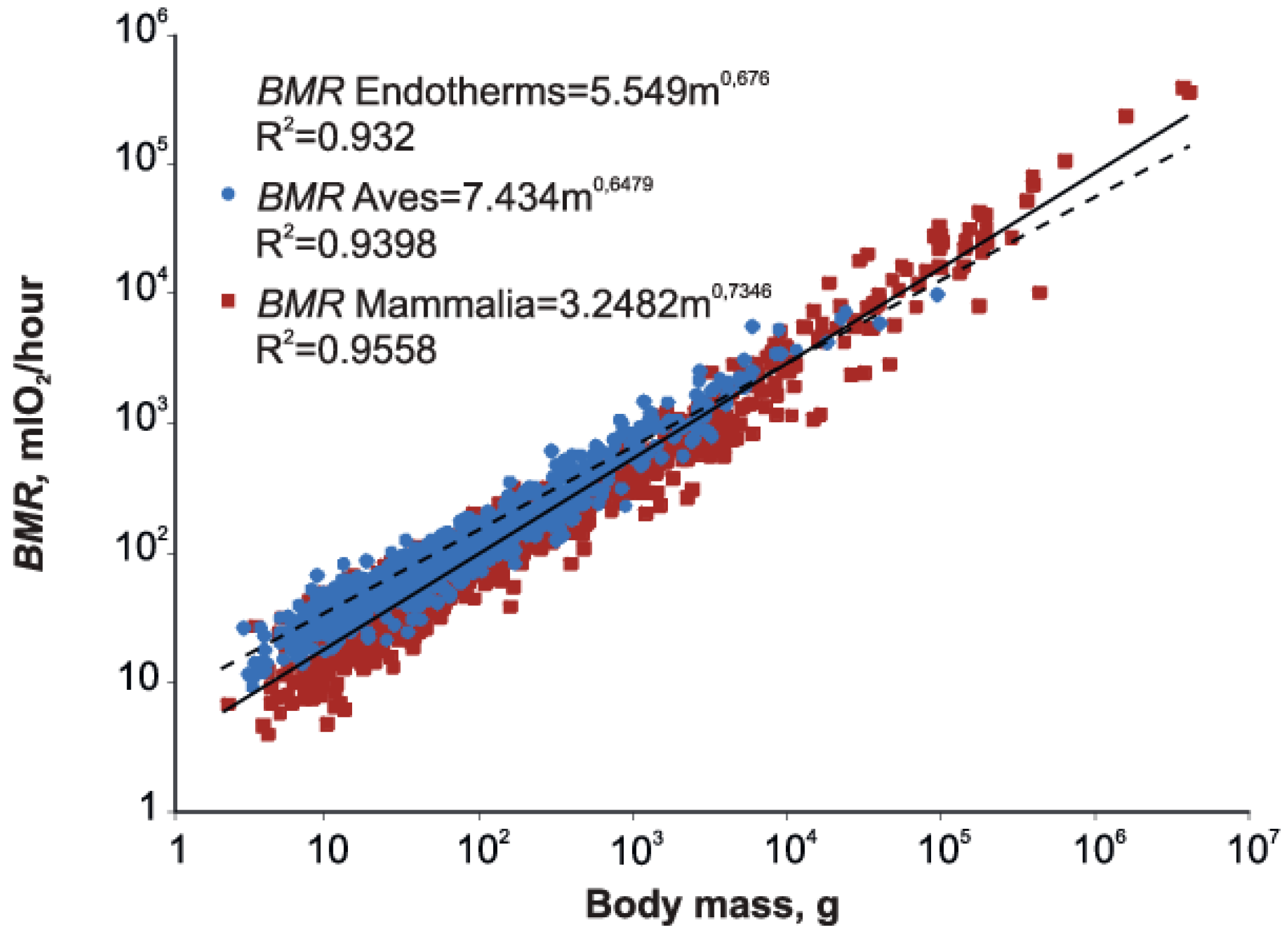
3.1. Allometry of Metabolic Rate in Endotherms
3.2. Mammalia vs. Aves
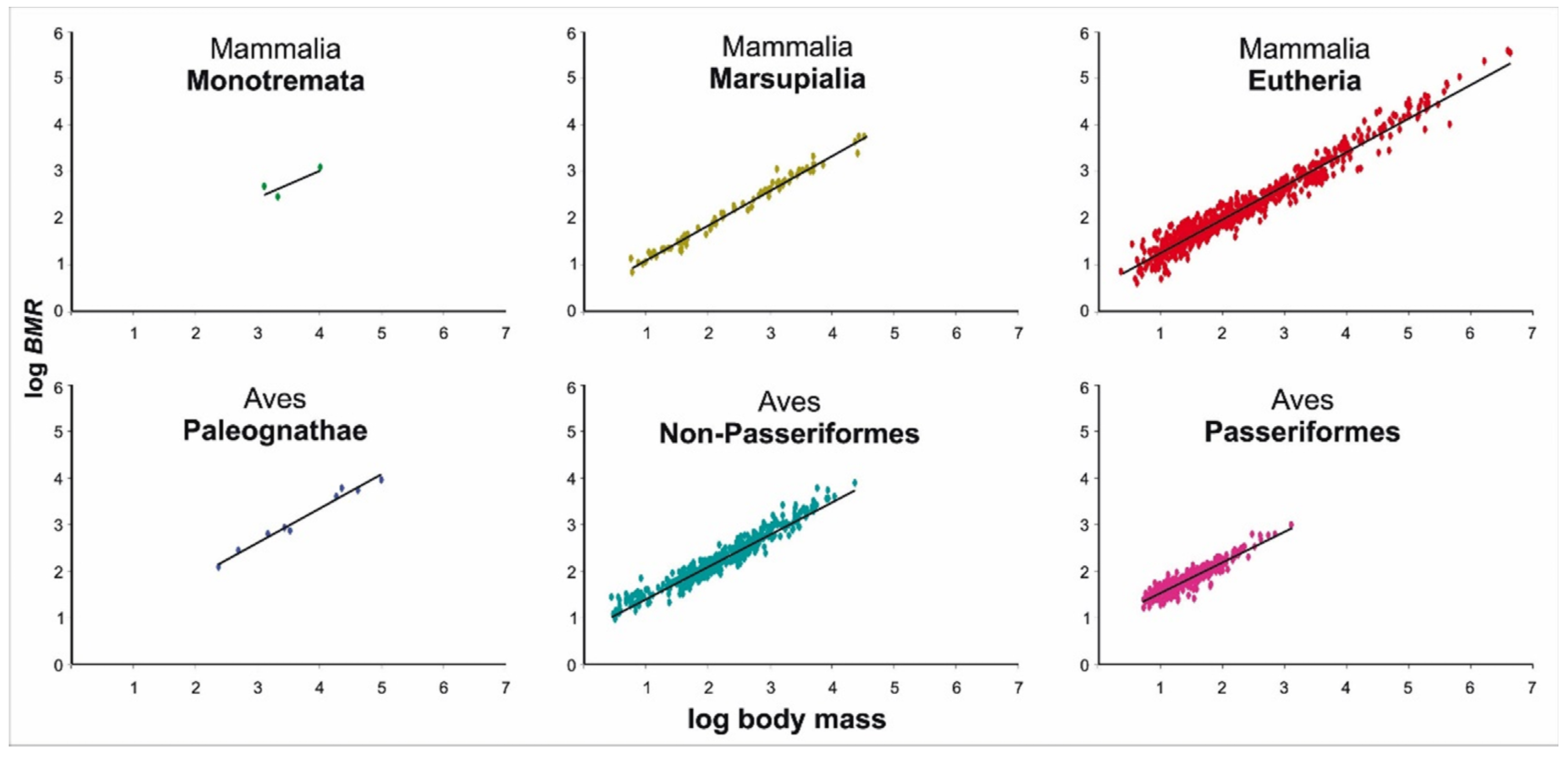
3.3. Allometry of Metabolic Rate in Major Clades of Mammals and Birds
3.4. Allometry of Metabolic Rate in Major Clades of Mammals and Birds
- (1)
- Simple linear regression model: y = a + bx, y = log(BMR), x = log(m)
- (2)
- Model with one slope and separate intercepts for each taxon: y = ai + bx, i = 1,2,…,6
- (3)
- Model with separate slopes and separate intercepts for each taxon: y = ai + bix
- (1)
- Simple linear regressionResidual standard error: 0.1797 on 1815 degrees of freedomMultiple R2: 0.9317, Adjusted R2: 0.9317 AIC = −1076.75, BIC = −1060.24
- (2)
- One slope, separate interceptsResidual standard error: 0.146 on 1810 degrees of freedomMultiple R2: 0.955, Adjusted R2: 0.955, AIC = −1822.26, BIC = −1778.22
- (3)
- Separate slopes, separate interceptsResidual standard error: 0.145 on 1805 degrees of freedomMultiple R2: 0.956, Adjusted R2: 0.956, AIC = −1847.0, BIC = −1775.44
3.5. Metabolic Allometry and Divergence Time of Various Groups of Endotherms
3.6. FMR (Field Metabolic Rate), BMR, and Divergence Time of Various Groups of Endotherms
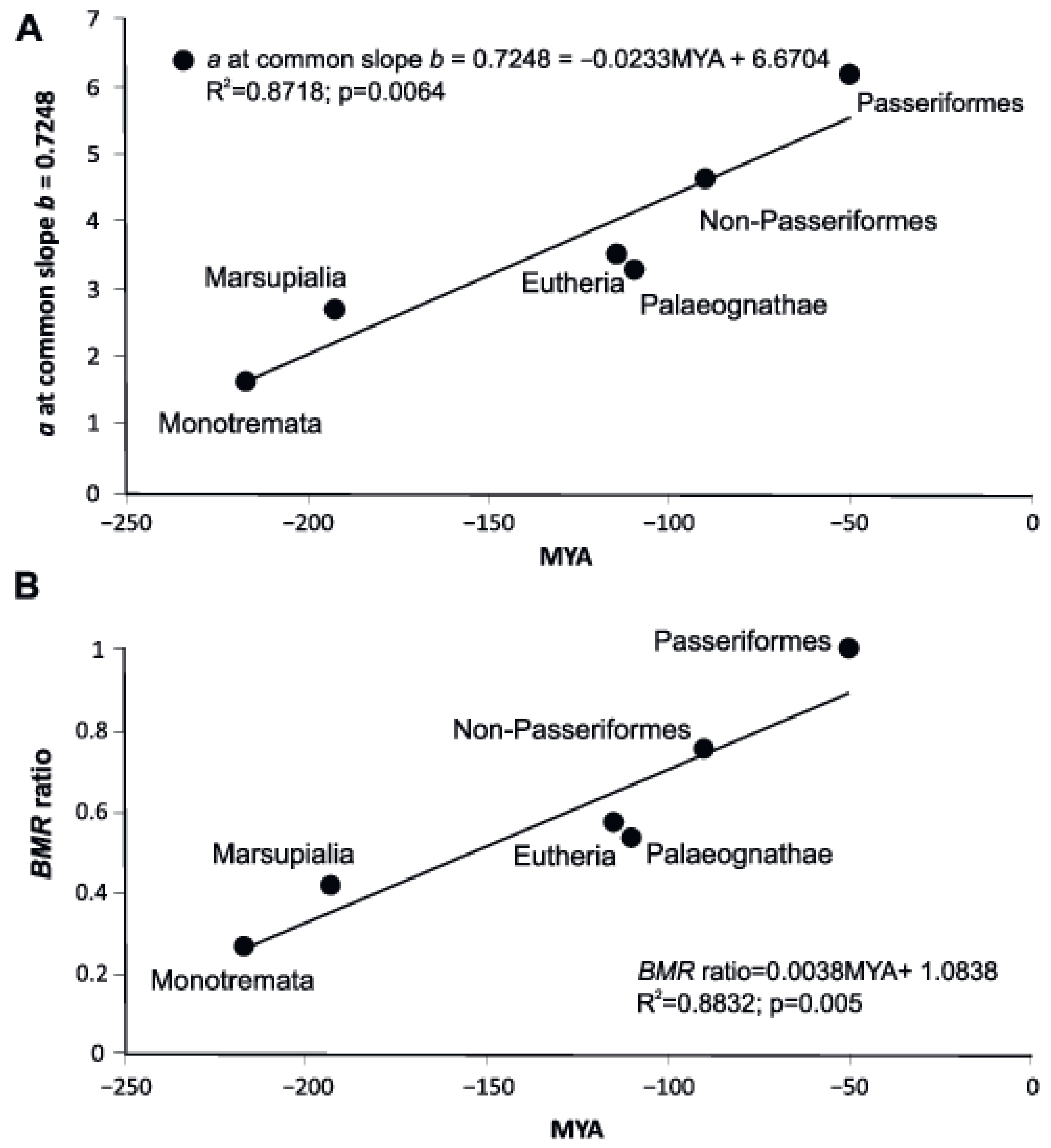
3.7. Relation between the Scaling Exponents and the Allometric Coefficients of Evolutionary Groups
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kleiber, M. Body size and metabolism. Hilgardia 1932, 6, 315–353. [Google Scholar] [CrossRef] [Green Version]
- Glazier, D.S. Beyond the ‘3/4-power law’: Variation in the intra- and interspecific scaling of metabolic rate in animals. Biol. Rev. Camb. Philos. Soc. 2005, 80, 611–662. [Google Scholar] [CrossRef]
- Glazier, D.S. A unifying explanation for diverse metabolic scaling in animals and plants. Biol. Rev. 2010, 85, 111–138. [Google Scholar] [CrossRef] [PubMed]
- Glazier, D.S. Scaling of metabolic scaling within physical limits. Systems 2014, 2, 425–450. [Google Scholar] [CrossRef] [Green Version]
- Glazier, D.S. Rediscovering and reviving old observations and explanations of metabolic scaling in living systems. Systems 2018, 6, 4. [Google Scholar] [CrossRef] [Green Version]
- Glazier, D.S. A Commentary “On the Interpretation of the Normalization Constant in the Scaling Equation” by Niklas, K.J., and S. T. Hammond, 2019. Front. Ecol. Evol. 2020, 6, 212. [Google Scholar] [CrossRef] [Green Version]
- Lovegrove, B.G. The Zoogeography of Mammalian Basal Metabolic Rate. Am. Nat. 2000, 156, 201–219. [Google Scholar] [CrossRef] [PubMed]
- Sibly, R.M.; Brown, J.H.; Kodric-Brown, A. Metabolic Ecology: A Scaling Approach; Wiley-Blackwel: Chichester, UK, 2012. [Google Scholar]
- Savage, V.M.; Deeds, E.J.; Fontana, W. Sizing up allometric scaling theory. PLoS Comput. Biol. 2008, 4, 116–171. [Google Scholar] [CrossRef] [PubMed]
- Savage, V.M.; Gillooly, J.F.; Woodruff, W.H.; West, G.B.; Allen, A.P.; Enquist, B.J.; Brown, J.H. The predominance of quarter-power scaling in biology. Funct. Ecol. 2004, 18, 257–282. [Google Scholar] [CrossRef]
- White, C.R.; Blackburn, T.M.; Seymour, R.S. Phylogenetically informed analysis of the allometry of mammalian basal metabolic rate supports neither geometric nor quarter-power scaling. Evolution 2009, 63, 2658–2667. [Google Scholar] [CrossRef] [PubMed]
- Banavar, J.R.; Maritan, A.; Rinaldo, A. Size and form in efficient transportation networks. Nature 1999, 399, 130–132. [Google Scholar] [CrossRef]
- Banavar, J.R.; Moses, M.E.; Brown, J.H.; Damuth, J.; Rinaldo, A.; Sibly, R.M.; Maritan, A. A general basis for quarter-power scaling in animals. Proc. Natl. Acad. Sci. USA 2010, 107, 15816–15820. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kozlowski, J.; Konarzewski, M. Is West, Brown and Enquist’s model of allometric scaling mathematically correct and biologically relevant? Funct. Ecol. 2004, 18, 283–289. [Google Scholar] [CrossRef]
- Kozlowski, J.; Konarzewski, M. West, Brown and Enquist’s model of allometric scaling again: The same questions remain. Funct. Ecol. 2005, 19, 739–743. [Google Scholar] [CrossRef]
- West, G.B.; Brown, J.H.; Enquist, B.J. A general model for the origin of allometric scaling laws in biology. Science 1997, 276, 122–126. [Google Scholar] [CrossRef] [PubMed]
- Chaui-Berlinck, J.G. A critical understanding of the fractal model of metabolic scaling. J. Exp. Biol. 2006, 209, 3045–3054. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takemoto, K. Heterogeneity of cells may explain allometric scaling of metabolic rate. BioSystems 2015, 130, 11–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ballesteros, F.J.; Martinez, V.J.; Luque, B.; Lacasa, L.; Valor, E.; Moya, A. On the thermodynamic origin of metabolic scaling. Sci. Rep. 2018, 8, 1448. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hulbert, A.J. A sceptics view: “Kleiber’s Law” or the “3/4 Rule” is neither a law nor a rule but rather an empirical approximation. Systems 2014, 2, 186–202. [Google Scholar] [CrossRef] [Green Version]
- Kearney, M.R.; White, C.R. Testing metabolic theories. Am. Nat. 2012, 180, 546–565. [Google Scholar] [CrossRef] [Green Version]
- Kolokotrones, T.; Savage, V.; Deeds, E.J.; Fontana, W. Curvature in metabolic scaling. Nature 2010, 464, 753–756. [Google Scholar] [CrossRef] [PubMed]
- White, C.R.; Kearney, M.R. Metabolic scaling in animals: Methods, empirical results, and theoretical explanations. Compr. Physiol. 2014, 4, 231–256. [Google Scholar] [PubMed]
- Glazier, D.S. Effects of metabolic level on the body size scaling of metabolic rate in birds and mammals. Proc. R. Soc. Lond. B Biol. Sci. 2008, 275, 1405–1410. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zotin, A.I.; Konoplev, V.A. Direction of the evolutionary progress of organisms. In Thermodynamic Bases of Biological Processes. Physiological Reactions and Adaptations; Walter de Gruyter: Berlin, Germany, 1978; pp. 341–347. [Google Scholar]
- Alekseeva, T.A.; Zotin, A.I. Standard metabolism and macrotaxonomy of crustaceans. Biol. Bull. Russ. Acad. Sci. 2001, 28, 157–162. [Google Scholar] [CrossRef]
- Zotin, A.A. Energetic macroevolution of invertebrates. Mosc. Univ. Biol. Sci. Bull. 2018, 45, 1–10. [Google Scholar] [CrossRef]
- Zotin, A.A. Energetic Macroevolution of Vertebrates. Mosc. Univ. Biol. Sci. Bull. 2018, 45, 299–309. [Google Scholar] [CrossRef]
- Atanasov, A.T.; Dimitrov, B.D. Changes of the power coefficient in the ‘metabolism–mass’ relationship in the evolutionary process of animals. BioSystems 2002, 66, 65–71. [Google Scholar] [CrossRef]
- Gould, S.J. Allometry and size in ontogeny and phylogeny. Biol. Rev. Camb. Philos. Soc. 1966, 41, 587–638. [Google Scholar] [CrossRef]
- Peters, R.H. The Ecological Implications of Body Size; Cambridge University Press: New York, NY, USA, 1983. [Google Scholar]
- Glazier, D.S. Metabolic scaling in complex living systems. Systems 2014, 2, 451–540. [Google Scholar] [CrossRef] [Green Version]
- Niklas, K.J.; Hammond, S.T. On the interpretation of the normalization constant in the scaling equation. Front. Ecol. Evol. 2019, 6, 212. [Google Scholar] [CrossRef] [Green Version]
- Gould, S.J. Ontogeny and Phylogeny; Belknap Press of Harvard University Press: Cambridge, MA, USA, 1977. [Google Scholar]
- Cheverud, J.M. Relationships among ontogenetic, static, and evolutionary allometry. Am. J. Phys. Anthropol. 1982, 59, 139–149. [Google Scholar] [CrossRef] [PubMed]
- DeLong, J.P.; Okie, J.G.; Moses, M.E.; Sibly, R.M.; Brown, J.H. Shifts in metabolic scaling, production, and efficiency across major evolutionary transitions of life. Proc. Natl. Acad. Sci. USA 2010, 107, 12941–12945. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hansen, T.F.; Orzack, S.H. Assessing current adaptation and phylogenetic inertia as explanations of trait evolution: The need for controlled comparisons. Evolution 2005, 59, 2063–2072. [Google Scholar] [CrossRef] [PubMed]
- Pélabon, C.; Firmat, C.; Bolstad, G.H.; Voje, K.L.; Houle, D.; Cassara, J.; Rouzic, A.L.; Hansen, T.F. Evolution of morphological allometry. Ann. N. Y. Acad. Sci. 2014, 1320, 58–75. [Google Scholar] [CrossRef] [Green Version]
- Uyeda, J.C.; Pennell, M.W.; Miller, E.T.; Maia, R.; McClain, C.R. The evolution of energetic scaling across the vertebrate tree of life. Am. Nat. 2017, 190, 185–199. [Google Scholar] [CrossRef]
- O’Dwyer, J.P.; Lake, J.K.; Ostling, A.; Savage, V.M.; Green, J.L. An integrative framework for stochastic, size-structured community assembly. Proc. Natl. Acad. Sci. USA 2009, 106, 6170–6175. [Google Scholar] [CrossRef] [Green Version]
- Harte, J. Maximum Entropy and Ecology: A Theory of Abundance, Distribution, and Energetics; Oxford University Press: Oxford, UK, 2011. [Google Scholar]
- Yvon-Durocher, G.; Allen, A.P. Linking community size structure and ecosystem functioning using metabolic theory. Philos. Trans. R. Soc. B 2012, 367, 2998–3007. [Google Scholar] [CrossRef] [Green Version]
- Gilbert, B.; Tunney, T.D.; McCann, K.S.; DeLong, J.P.; Vasseur, D.A.; Savage, V.; Shurin, J.B.; Dell, A.I.; Barton, T.B.; Harley, C.D.G.; et al. A bioenergetic framework for the temperature dependence of trophic interactions. Ecol. Lett. 2014, 17, 902–914. [Google Scholar] [CrossRef]
- Enquist, B.J.; Norberg, J.; Bonser, S.P.; Violle, C.; Webb, C.T.; Henderson, A.; Sloat, L.L.; Savage, V.M. Scaling from traits to ecosystems: Developing a general trait driver theory via integrating trait-based and metabolic scaling theories. Adv. Ecol. Res. 2015, 52, 249–318. [Google Scholar]
- Harte, J.; Rominger, A.; Zhang, W. Integrating macroecological metrics and community taxonomic structure. Ecol. Lett. 2015, 18, 1068–1077. [Google Scholar] [CrossRef]
- Burton, T.; Killen, S.S.; Armstrong, J.D.; Metcalfe, N.B. What causes intraspecific variation in resting metabolic rate, and what are its ecological consequences? Proc. R. Soc. B 2011, 278, 3465–3473. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- White, C.R.; Kearney, M.R. Determinants of inter-specific variation in basal metabolic rate. J. Comp. Physiol. B 2013, 183, 1–26. [Google Scholar] [CrossRef] [PubMed]
- Pettersen, A.K.; Marshall, D.J.; White, C.R. Understanding variation in metabolic rate. J. Exp. Biol. 2018, 221, jeb166876. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- White, C.R.; Marshall, D.J.; Alton, L.A.; Arnold, P.A.; Beaman, J.E.; Bywater, C.L.; Condon, C.; Crispin, T.S.; Janetzki, A.; Pirtle, E.; et al. The origin and maintenance of metabolic allometry in animals. Nat. Ecol. Evol. 2019, 3, 598–603. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Uyeda, J.C.; Bone, N.; McHugh, S.; Rolland, J.; Pennell, M.W. How should functional relationships be evaluated using phylogenetic comparative methods? A case study using metabolic rate and body temperature. Evolution 2021, 75, 1097–1105. [Google Scholar] [CrossRef] [PubMed]
- Arnold, P.A.; Delean, S.; Cassey, P.; White, C.R. Meta-analysis reveals that resting metabolic rate is not consistently related to fitness and performance in animals. J. Comp. Physiol. B 2021, 191, 1097–1110. [Google Scholar] [CrossRef]
- Benedict, F.G. Vital Energetics. A Study in Comparative Basal Metabolism; Carnegie Inst.: Washington, DC, USA, 1938. [Google Scholar]
- King, J.R.; Farner, D.S. Energy metabolism, thermoregulation, and body temperature. In Biology and Comparative Physiology of Birds, 2nd ed.; Marshall, A.J., Ed.; Academic Press: New York, NY, USA, 1961; p. 2. [Google Scholar]
- Hemmingsen, A.M. The relation of standard (basal) energy metabolism to total fresh weight of living organisms. Rep. Steno Mem. Hosp. 1950, 4, 7–58. [Google Scholar]
- Hemmingsen, A.M. Energy metabolism as related to body size and respiratory surface, and its evolution. Rep. Steno Mem. Hosp. 1960, 9, 1–110. [Google Scholar]
- Lasiewski, R.C.; Dawson, W.R. A reexamination of the relation between standard metabolic rate and body weight in birds. Condor 1967, 69, 13–23. [Google Scholar] [CrossRef]
- Dodds, P.S.; Rothman, D.H.; Weitz, J.S. Re-examination of the “3/4-law” of metabolism. J. Theor. Biol. 2001, 209, 9–27. [Google Scholar] [CrossRef] [Green Version]
- McNab, B.K. An analysis of the factors that influence the level and scaling of mammalian BMR. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2008, 151, 5–28. [Google Scholar] [CrossRef] [PubMed]
- McNab, B.K. Ecological factors affect the level and scaling of avian BMR. Mol. Integr. Physiol. A Mol. Integr. Physiol. 2009, 152, 22–45. [Google Scholar] [CrossRef]
- Capellini, I.; Venditti, C.; Barton, R.A. Phylogeny and metabolic scaling in mammals. Ecology 2010, 91, 2783–2793. [Google Scholar] [CrossRef] [PubMed]
- Genoud, M.; Isler, K.; Martin, R.D. Comparative analyses of basal rate of metabolism in mammals: Data selection does matter. Biol. Rev. 2018, 93, 404–438. [Google Scholar] [CrossRef]
- Livezey, B.C.; Zusi, R.L. Higher-order phylogeny of modern birds (Theropoda, Aves: Neornithes) based on comparative anatomy. II. Analysis and discussion. Zool. J. Linn. Soc. 2007, 149, 1–95. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hackett, S.J.; Kimball, R.T.; Reddy, S.; Bowie, R.C.K.; Braun, E.L.; Braun, M.J.; Chojnowski, J.L.; Cox, W.A.; Han, K.-L.; Harshman, J.; et al. A phylogenomic study of birds reveals their evolutionary history. Science 2008, 320, 1763–1768. [Google Scholar] [CrossRef]
- Pacheco, M.A.; Battistuzzi, F.U.; Lentino, M.; Aguilar, R.; Kumar, S.; Escalante, A.A. Evolution of Modern Birds Revealed by Mitogenomics: Timing the Radiation and Origin of Major Orders. Mol. Biol. Evol. 2011, 28, 1927–1942. [Google Scholar] [CrossRef] [Green Version]
- Prum, R.; Berv, J.; Dornburg, A.; Field, D.J.; Townsend, J.P.; Lemmon, E.M.; Lemmon, A.R. A comprehensive phylogeny of birds (Aves) using targeted next-generation DNA sequencing. Nature 2015, 526, 569–573. [Google Scholar] [CrossRef]
- Gill, F.; Donsker, D.; Rasmussen, P. (Eds.) IOC World Bird List. 2020. (v. 10.2). Available online: http://www.worldbirdnames.org/ (accessed on 19 March 2020).
- McNab, B.K. Avian energetics: The passerine/non-passerine dichotomy. Comp. Biochem. Physiol. A 2016, 191, 152–155. [Google Scholar] [CrossRef]
- Gavrilov, V.M.; Golubeva, T.B.; Bushuev, A.V. Evolution of metabolic scaling among the tetrapod: Effect of phylogeny, the geologic time of class formation, and uniformity of species within a class. Integr. Zool. 2021, 1–14. [Google Scholar] [CrossRef]
- Isaac, N.J.; Carbone, C. Why are metabolic scaling exponents so controversial? Quantifying variance and testing hypotheses. Ecol. Lett. 2010, 13, 728–735. [Google Scholar] [CrossRef] [PubMed]
- White, C.R.; Seymour, R.S. Mammalian basal metabolic rate is proportional to body mass 2/3. Proc. Natl. Acad. Sci. USA 2003, 100, 4046–4049. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bokma, F. Evidence against universal metabolic allometry. Funct. Ecol. 2004, 18, 184–187. [Google Scholar] [CrossRef]
- Cyr, H.; Walker, S.C. An illusion of mechanistic understanding. Ecology 2004, 85, 1802–1804. [Google Scholar] [CrossRef]
- Farrell-Gray, C.C.; Gotelli, N.J. Allometric exponents support a 3/4-power scaling law. Ecology 2005, 86, 2083–2087. [Google Scholar] [CrossRef] [Green Version]
- Sieg, A.E.; O’Connor, M.P.; McNair, J.N.; Grant, B.W.; Agosta, S.J.; Dunham, A.E. Mammalian metabolic allometry: Do intraspecific variation, phylogeny, and regression models matter? Am. Nat. 2009, 174, 720–733. [Google Scholar] [CrossRef] [Green Version]
- McClain, C.R.; Allen, A.P.; Tittensor, D.P.; Rex, M.A. Energetics of life on the deep seafloor. Proc. Natl. Acad. Sci. USA 2012, 109, 15366–15371. [Google Scholar] [CrossRef] [Green Version]
- Hudson, L.N.; Isaac, N.J.; Reuman, D.C. The relationship between body mass and field metabolic rate among individual birds and mammals. J. Anim. Ecol. 2013, 82, 1009–1020. [Google Scholar] [CrossRef] [Green Version]
- Kendeigh, S.C.; Dolnik, V.R.; Gavrilov, V.M. Avian energetics. In Granivorous Birds in Ecosystem; Pinowski, J., Kendeigh, S.C., Eds.; Cambridge University Press: Cambridge, UK, 1977; pp. 127–204. [Google Scholar]
- Gavrilov, V.M. Ecological and scaling analysis of the energy expenditure of rest, activity, flight, and evaporative water loss in passeriformes and non-passeriformes in relation to seasonal migrations and to the occupation of boreal stations in high and moderate latitudes. Q. Rev. Biol. 2014, 89, 107–150. [Google Scholar]
- Nagy, K. Field metabolic rate and body size. J. Exp. Biol. 2005, 208, 1621–1625. [Google Scholar] [CrossRef] [Green Version]
- McNab, B.K. On the utility of uniformity in the definition of basal rate of metabolism. Physiol. Zool. 1997, 70, 718–720. [Google Scholar] [CrossRef]
- Phillips, M.J.; Fruciano, C. The soft explosive model of placental mammal evolution. BMC Evol. Biol. 2018, 18, 104. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beck, R.M.D.; Baillie, C. Improvements in the fossil record may largely resolve current conflicts between morphological and molecular estimates of mammal phylogeny. Proc. R. Soc. B 2018, 285, 1632. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brusatte, S.L.; O’Connor, J.K.; Jarvis, E.D. The Origin and Diversification of Birds. Curr. Biol. 2015, 2519, R888–R898. [Google Scholar] [CrossRef] [Green Version]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2019. [Google Scholar]
- Fox, J.; Weisberg, S. An R Companion to Applied Regression, 2nd ed.; SAGE Publications, Inc.: Thousand Oaks, CA, USA, 2014. [Google Scholar]
- Killen, S.S.; Atkinson, D.; Glazier, D.S. The intraspecific scaling of metabolic rate with body mass in fishes depends on lifestyle and temperature. Ecol. Lett. 2010, 13, 184–193. [Google Scholar] [CrossRef] [PubMed]
- Bushuev, A.; Tolstenkov, O.; Zubkova, E.; Solovyeva, E.; Kerimov, A. Basal metabolic rate in free-living tropical birds: The influence of phylogenetic, behavioral, and ecological factors. Curr. Zool. 2018, 64, 33–43. [Google Scholar] [CrossRef] [Green Version]
- Freckleton, R.P.; Harvey, P.H.; Pagel, M. Phylogenetic analysis and comparative data: A test and review of evidence. Am. Nat. 2002, 160, 712–726. [Google Scholar] [CrossRef]
- Grafen, A. The phylogenetic regression. Philos. Trans. R. Soc. B: Biol. Sci. 1989, 326, 119–157. [Google Scholar]
- Orme, D.; Freckleton, R.; Thomas, G.; Petzoldt, T.; Fritz, S.; Isaac, N.; Pearse, W. Caper: Comparative Analyses of Phylogenetics and Evolution in R. R Package Version 1.0.1. 2018. Available online: https://cran.r-project.org/web/packages/caper/index.html (accessed on 1 February 2020).
- Pagel, M. Inferring the historical patterns of biological evolution. Nature 1999, 401, 877–884. [Google Scholar] [CrossRef]
- White, C.R.; Phillips, N.F.; Seymour, R.S. The scaling and temperature dependence of vertebrate metabolism. Biol. Lett. 2006, 2, 125–127. [Google Scholar] [CrossRef] [Green Version]
- West, G.B.; Brown, J.H.; Enquist, B.J. The fourth dimension of life: Fractal geometry and allometric scaling of organisms. Science 1999, 284, 1677–1679. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McMahon, T.A. Allometry and Biomechanics: Limb Bones in Adult Ungulates. Am. Nat. 1975, 109, 547–563. [Google Scholar] [CrossRef]
- Bejan, A. The constructal law of organization in nature: Tree-shaped flows and body size. J. Exp. Biol. 2005, 208, 1677–1686. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bejan, A.; Lorente, S. The constructal law of design and evolution in nature. Philos. Trans. R. Soc. Lond. B, Biol. Sci. 2010, 365, 1335–1347. [Google Scholar] [CrossRef] [Green Version]
- Banavar, J.R.; Maritan, A.; Rinaldo, A. Rivers, blood and transportation networks—Reply. Nature 2000, 408, 160. [Google Scholar] [CrossRef]
- Darveau, C.-A.; Suarez, R.K.; Andrews, R.D.; Hochachka, P.W. Allometric cascade as a unifying principle of body mass effects on metabolism. Nature 2002, 417, 166–170. [Google Scholar] [CrossRef]
- Etienne, R.S.; Apol, M.E.F.; Olff, H.A.N. Demystifying the West, Brown and Enquist model of the allometry of metabolism. Funct. Ecol. 2006, 20, 394–399. [Google Scholar] [CrossRef] [Green Version]
- Sapoval, B.; Filoche, M.; Weibel, E.R. Smaller is better—But not too small: A physical scale for the design of the mammalian pulmonary acinus. Proc. Natl. Acad. Sci. USA 2002, 99, 10411–10416. [Google Scholar] [CrossRef] [Green Version]
- Makarieva, A.M.; Gorshkov, V.G.; Li, B.-L.; Chown, S.L.; Reich, P.B.; Gavrilov, V.M. Mean mass-specific metabolic rates are strikingly similar across life’s major domains: Evidence for life’s metabolic optimum. Proc. Natl. Acad. Sci. USA 2008, 105, 16994–16999. [Google Scholar] [CrossRef] [Green Version]
- Makarieva, A.M.; Gorshkov, V.G.; Li, B.L. Revising the distributive networks models of West, Brown and Enquist 1997 and Banavar, Maritan and Rinaldo 1999: Metabolic inequity of living tissues provides clues for the observed allometric scaling rules. J. Theor. Biol. 2005, 237, 291–301. [Google Scholar] [CrossRef]
- McNab, B.K. What determines the basal rate of metabolism? J. Exp. Biol. 2019, 222, jeb205591. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schmidt-Nielsen, K. Scaling: Why Is Animal Size So Important? Cambridge University Press: New York, NY, USA, 1984. [Google Scholar]
- Kempes, C.P.; Wolpert, D.; Cohen, Z.; Pérez-Mercader, J. The thermodynamic efficiency of computations made in cells across the range of life. Philos. Trans. Royal Soc. A 2017, 375, 20160343. [Google Scholar] [CrossRef] [PubMed]
- Makarieva, A.M.; Nefiodov, A.V.; Li, B.L. Life’s Energy and Information: Contrasting Evolution of Volume- versus Surface-Specific Rates of Energy Consumption. Entropy 2020, 22, 1025. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | Number of Species | Body Mass Range, g | OLS: a ± SE | OLS: b ± SE | OLS: R2 | Pagel’s λ | PGLS: a ± SE | PGLS: b ± SE | PGLS: R2 |
|---|---|---|---|---|---|---|---|---|---|
| Mammalia | 817 | 2.2–4,037,500 | 3.248 ± 0.107 | 0.735 ± 0.006 | 0.956 | 0.870 | 2.357 ± 0.632 | 0.735 ± 0.009 | 0.888 |
| Monotremata | 3 | 1284–10,300 | 5.861± 0.512 | 0.565 ± 0.387 | 0.681 | 0.000 | 5.861 ± NA | 0.565 ± 0.387 | 0.681 |
| Marsupialia | 84 | 5.4–32,490 | 2.300 ± 0.152 | 0.753 ± 0.011 | 0.983 | 0.214 | 2.407 ± 0.222 | 0.746 ± 0.013 | 0.976 |
| Eutheria | 730 | 2.2–4,037,500 | 3.326 ± 0.115 | 0.736 ± 0.006 | 0.956 | 0.813 | 2.910 ± 0.393 | 0.733 ± 0.011 | 0.874 |
| Aves | 1000 | 2.8–92,400 | 7.435 ± 0.167 | 0.648 ± 0.005 | 0.940 | 0.664 | 5.514 ± 0.605 | 0.679 ± 0.010 | 0.830 |
| Paleognathae | 9 | 220.8–92,400 | 3.221 ± 1.147 | 0.727 ± 0.041 | 0.978 | 0.000 | 3.221 ± 0.871 | 0.727 ± 0.041 | 0.978 |
| Non-Passeriformes | 404 | 3.2–23,370 | 5.507 ± 0.262 | 0.691 ± 0.009 | 0.939 | 0.630 | 4.833 ± 0.589 | 0.708 ± 0.014 | 0.865 |
| Group | Number of Species | PGLS: a, mL O2/h at b = 0.698 | R2 for a at b = 0.698 | OLS: a, mL O2/h at b = 0.704 | R2 for Regression at b = 0.704 | OLS:a, mL O2/h at b = 0.7248 | R2 for Regression at b = 0.7248 |
|---|---|---|---|---|---|---|---|
| Monotremata | 3 | 2.02 | 0.6570 | 1.92 | 0.6590 | 1.63 | 0.666 |
| Marsupialia | 84 | 3.14 | 0.9740 | 3.03 | 0.9760 | 2.69 | 0.980 |
| Eutheria | 730 | 4.07 | 0.9460 | 3.94 | 0.9480 | 3.53 | 0.952 |
| Paleognathae | 9 | 4.14 | 0.9740 | 3.93 | 0.9750 | 3.29 | 0.978 |
| Non-Passeriformes | 404 | 5.32 | 0.9360 | 5.16 | 0.9370 | 4.65 | 0.939 |
| Passeriformes | 587 | 6.72 | 0.8620 | 6.59 | 0.8640 | 6.18 | 0.868 |
| Group | Number of Species | PGLS: a/aPass (BMR ratio) | R2 b = 0.698337 | OLS: a/aPass (BMR ratio) | R2 b = 0.70449 | Indicator Variables a/aPass (BMR ratio) | R2 b = 0.7248 |
|---|---|---|---|---|---|---|---|
| Monotremata | 3 | 0.3000 | 0.6570 | 0.2915 | 0.6590 | 0.264 | 0.666 |
| Marsupialia | 84 | 0.4670 | 0.9740 | 0.4600 | 0.9760 | 0.435 | 0.980 |
| Eutheria | 730 | 0.6054 | 0.9460 | 0.5977 | 0.9480 | 0.571 | 0.952 |
| Paleognathae | 9 | 0.6153 | 0.9740 | 0.5960 | 0.9750 | 0.532 | 0.978 |
| Non-Passeriformes | 404 | 0.7924 | 0.9360 | 0.7833 | 0.9370 | 0.752 | 0.939 |
| Passeriformes | 578 | 1.0000 | 0.8620 | 1.0000 | 0.8640 | 1.0000 | 0.868 |
| Group | BMR a at b = 0.7248 | FMR a at b = 0.6851 | BMR/FMR |
|---|---|---|---|
| Monotremata | 1.63 | 7.81 | 0.21 |
| Marsupialia | 2.69 | 11.48 | 0.23 |
| Eutheria | 3.53 | 12.52 | 0.28 |
| Paleognathae | 3.29 | 15.33 | 0.21 |
| Non-Passeriformes | 4.65 | 21.54 | 0.22 |
| Passeriformes | 6.18 | 21.32 | 0.29 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gavrilov, V.M.; Golubeva, T.B.; Warrack, G.; Bushuev, A.V. Metabolic Scaling in Birds and Mammals: How Taxon Divergence Time, Phylogeny, and Metabolic Rate Affect the Relationship between Scaling Exponents and Intercepts. Biology 2022, 11, 1067. https://doi.org/10.3390/biology11071067
Gavrilov VM, Golubeva TB, Warrack G, Bushuev AV. Metabolic Scaling in Birds and Mammals: How Taxon Divergence Time, Phylogeny, and Metabolic Rate Affect the Relationship between Scaling Exponents and Intercepts. Biology. 2022; 11(7):1067. https://doi.org/10.3390/biology11071067
Chicago/Turabian StyleGavrilov, Valery M., Tatiana B. Golubeva, Giles Warrack, and Andrey V. Bushuev. 2022. "Metabolic Scaling in Birds and Mammals: How Taxon Divergence Time, Phylogeny, and Metabolic Rate Affect the Relationship between Scaling Exponents and Intercepts" Biology 11, no. 7: 1067. https://doi.org/10.3390/biology11071067
APA StyleGavrilov, V. M., Golubeva, T. B., Warrack, G., & Bushuev, A. V. (2022). Metabolic Scaling in Birds and Mammals: How Taxon Divergence Time, Phylogeny, and Metabolic Rate Affect the Relationship between Scaling Exponents and Intercepts. Biology, 11(7), 1067. https://doi.org/10.3390/biology11071067