
Image Classification of Amazon Parrots by Deep Learning: A Potentially Useful Tool for Wildlife Conservation


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Collection of Images
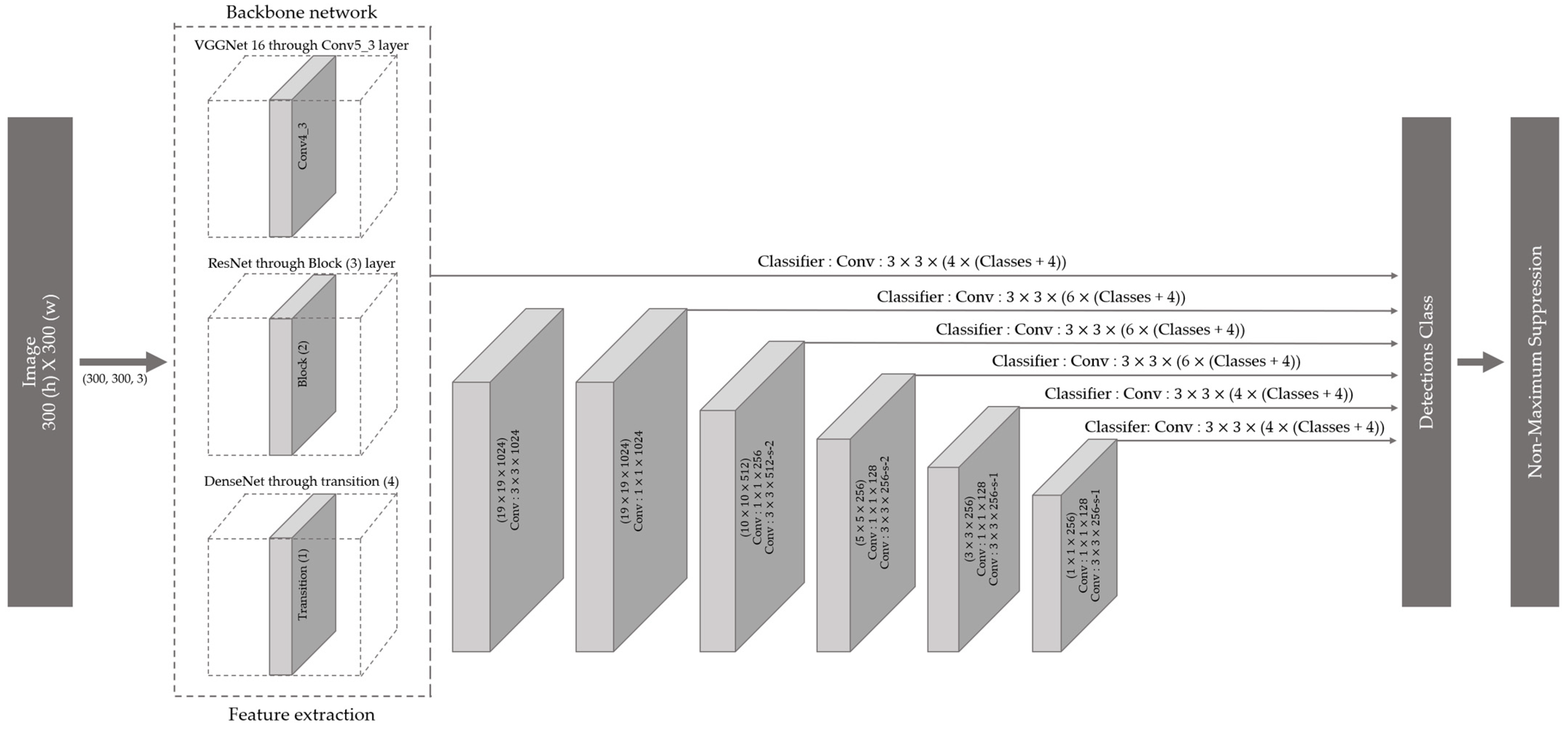
2.2. Training of Deep Learning Models
2.3. Evaluation of Model Performances
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Blanco, G.; Hiraldo, F.; Tella, J.L. Ecological functions of parrots: An integrative perspective from plant life cycle to ecosystem functioning. Emu 2018, 118, 36–49. [Google Scholar] [CrossRef]
- Symes, C.T.; Perrin, M.R. Feeding biology of the greyheaded parrot, Poicephalus fuscicollis suahelicus (Reichenow), in Northern Province, South Africa. Emu 2003, 103, 49–58. [Google Scholar] [CrossRef]
- Olah, G.; Butchart, S.H.; Symes, A.; Guzmán, I.M.; Cunningham, R.; Brightsmith, D.J.; Heinsohn, R. Ecological and socio-economic factors affecting extinction risk in parrots. Biodivers. Conserv. 2016, 25, 205–223. [Google Scholar] [CrossRef]
- Bush, E.R.; Baker, S.E.; Macdonald, D.W. Global trade in exotic pets 2006–2012. Conserv. Biol. 2014, 28, 663–676. [Google Scholar] [CrossRef] [PubMed]
- Scheffers, B.R.; Oliveira, B.F.; Lamb, I.; Edwards, D.P. Global wildlife trade across the tree of life. Science 2019, 366, 71–76. Available online: https://www.science.or (accessed on 27 June 2022). [CrossRef]
- Forshaw, J.M. Parrots of the World; Princeton University Press: Princeton, NJ, USA, 2010; pp. 278–308. [Google Scholar]
- Parr, M.; Juniper, T. Parrots: A Guide to Parrots of the World; Bloomsbury Publishing: London, UK, 2010; pp. 523–554. [Google Scholar]
- Del Hoyo, J. All the Birds of the World; Lynx Edicions: Barcelona, Spain, 2020; pp. 364–366. [Google Scholar]
- BirdLife International Data Zone. Available online: http://datazone.birdlife.org/species/search (accessed on 27 June 2022).
- Checklist of CITES Species. Available online: https://checklist.cites.org/#/en (accessed on 20 June 2022).
- Chan, D.T.C.; Poon, E.S.K.; Wong, A.T.C.; Sin, S.Y.W. Global trade in parrots—Influential factors of trade and implications for conservation. Glob. Ecol. Conserv. 2021, 30, e01784. [Google Scholar] [CrossRef]
- Sánchez-Mercado, A.; Ferrer-Paris, J.R.; Rodríguez, J.P.; Tella, J.L. A literature synthesis of actions to tackle illegal parrot trade. Diversity 2021, 13, 191. [Google Scholar] [CrossRef]
- Sinovas, P.; Price, B.; King, E.; Hinsley, A.; Pavitt, A. Wildlife Trade in the Amazon Countries: An Analysis of Trade in CITES Listed Species; UN Environment—World Conservation Monitoring Centre: Cambridge, UK, 2017; pp. 31–34. [Google Scholar]
- Cardador, L.; Lattuada, M.; Strubbe, D.; Tella, J.L.; Reino, L.; Figueira, R.; Carrete, M. Regional bans on wild-bird trade modify invasion risks at a global scale. Conserv. Lett. 2017, 10, 717–725. [Google Scholar] [CrossRef]
- Seo, M.H.; Kim, H.J. The Guideline for Import and Export Review of CITES Species; National Institute of Biological Resources: Incheon, Korea, 2016; pp. 76–132. [Google Scholar]
- Schneider, S.; Taylor, G.W.; Kremer, S. Deep learning object detection methods for ecological camera trap data. In Proceedings of the 2018 15th Conference on Computer and Robot Vision (CRV), Toronto, ON, Canada, 9–11 May 2018; pp. 321–328. [Google Scholar]
- Rich, L.N.; Davis, C.L.; Farris, Z.J.; Miller, D.A.; Tucker, J.M.; Hamel, S.; Farhadinia, M.S.; Steenweg, R.; Di Bitetti, M.S.; Thapa, K. Assessing global patterns in mammalian carnivore occupancy and richness by integrating local camera trap surveys. Global Ecol. Biogeogr. 2017, 26, 918–929. [Google Scholar] [CrossRef]
- Tempier, L. Building worldwide expertise to detect and seize illegally traded wildlife. In UN Chronicle: Illegal Wildlife Trad; United Nations: New York, NY, USA, 2014; Volume LI, pp. 25–28. [Google Scholar]
- Trail, P.W. Morphological analysis: A powerful tool in wildlife forensic biology. Forensic Sci. Int. 2021, 1, 100025. [Google Scholar] [CrossRef]
- Mahendiran, M.; Parthiban, M.; Azeez, P.A.; Nagarajan, R. In situ measurements of animal morphological features: A non-invasive method. Methods Ecol. Evol. 2018, 9, 613–623. [Google Scholar] [CrossRef]
- Esipova, O.; Love, E.; Noakes, A.; Schatz, A.; Swartz, K.; Vallianos, C. Wildlife Trafficking Detection Tools: Best Practices and Application to the Illegal Rhino Horn Trade; Conservation International: Arlington, VA, USA, 2021; pp. 30–33. [Google Scholar]
- Hopkins, G.; Freckleton, R.P. Declines in the numbers of amateur and professional taxonomists: Implications for conservation. Anim. Conserv. 2002, 5, 245–249. [Google Scholar] [CrossRef]
- Eberhard, J.R.; Bermingham, E. Phylogeny and biogeography of the Amazona ochrocephala (Aves: Psittacidae) complex. Auk 2004, 121, 318–332. [Google Scholar] [CrossRef]
- Wenner, T.J.; Russello, M.A.; Wright, T.F. Cryptic species in a Neotropical parrot: Genetic variation within the Amazona farinosa species complex and its conservation implications. Conserv. Genet. 2012, 13, 1427–1432. [Google Scholar] [CrossRef]
- Abe, H.; Hayano, A.; Inoue-Murayama, M. Forensic species identification of large macaws using DNA barcodes and microsatellite profiles. Mol. Biol. Rep. 2012, 39, 693–699. [Google Scholar] [CrossRef]
- Kim, J.I.; Do, T.D.; Lee, D.; Yeo, Y.; Kim, C.B. Application of Cytochrome b Gene Sequences for Identification of Parrots from Korean Zoos. Anim. Syst. Evol. Divers. 2020, 36, 216–221. [Google Scholar] [CrossRef]
- Brandis, K.J.; Meagher, P.J.; Tong, L.J.; Shaw, M.; Mazumder, D.; Gadd, P.; Ramp, D. Novel detection of provenance in the illegal wildlife trade using elemental data. Sci. Rep. 2018, 8, 15380. [Google Scholar] [CrossRef]
- Wäldchen, J.; Mäder, P. Machine learning for image based species identification. Methods Ecol. Evol. 2018, 9, 2216–2225. [Google Scholar] [CrossRef]
- Schneider, S.; Taylor, G.W.; Linquist, S.; Kremer, S.C. Past, present and future approaches using computer vision for animal re-identification from camera trap data. Methods Ecol. Evol. 2019, 10, 461–470. [Google Scholar] [CrossRef]
- Christin, S.; Hervet, É.; Lecomte, N. Applications for deep learning in ecology. Methods Ecol. Evol. 2019, 10, 1632–1644. [Google Scholar] [CrossRef]
- Surkovic, E.; Smith, L.; Edbury, M.; Pilkington, J.; Taylor, L.; Kahunzire, A.; Kett, M. Science: Tackling the Illegal Wildlife Trade; The Royal Society: London, UK, 2018; pp. 23–25. [Google Scholar]
- Krizhevsky, A.; Sutskever, I.; Hinton, G.E. Imagenet classification with deep convolutional neural networks. Adv. Neural Inf. Process Syst. 2012, 25, 1097–1105. [Google Scholar] [CrossRef]
- Ren, S.; He, K.; Girshick, R.; Sun, J. Faster R-CNN: Towards real-time object detection with region proposal networks. Adv. Neural Inf. Process Syst. 2015, 28, 91–99. [Google Scholar] [CrossRef] [PubMed]
- Redmon, J.; Divvala, S.; Girshick, R.; Farhadi, A. You only look once: Unified, real-time object detection. In Proceedings of the IEEE Conference on Computer Vision and Pattern Recognition, Las Vegas, NV, USA, 27–30 June 2016; pp. 779–788. [Google Scholar]
- Liu, W.; Anguelov, D.; Erhan, D.; Szegedy, C.; Reed, S.; Fu, C.Y.; Berg, A.C. Ssd: Single shot multibox detector. In Proceedings of the European Conference on Computer Vision, Amsterdam, The Netherlands, 8–16 October 2016; pp. 21–37. [Google Scholar]
- Hung, P.D.; Kien, N.N. SSD-Mobilenet Implementation for classifying fish species. In Intelligent Computing and Optimization; Vasant, P., Zelinka, I., Weber, G.W., Eds.; Springer International Publishing: Cham, Switzerland, 2020; pp. 399–408. [Google Scholar]
- Jang, W.; Lee, E.C. Multi-class parrot image classification including subspecies with similar appearance. Biology 2021, 10, 1140. [Google Scholar] [CrossRef] [PubMed]
- Akdemir, K.Ü.; Alaybeyoğlu, E. Classification of Red Mullet, Bluefish and Haddock caught in the Black sea by “Single Shot Multibox Detection”. In Proceedings of the IEEE International Conference on INnovations in Intelligent SysTems and Applications (INISTA), Kocaeli, Turkey, 25–27 August 2021; pp. 1–4. [Google Scholar]
- Patel, A.; Cheung, L.; Khatod, N.; Matijosaitiene, I.; Arteaga, A.; Gilkey, J.W., Jr. Revealing the unknown: Real-time recognition of Galápagos snake species using deep learning. Animals 2020, 10, 806. [Google Scholar] [CrossRef] [PubMed]
- DarkLabel. Available online: https://darkpgmr.tistory.com/16 (accessed on 5 March 2022).
- Simonyan, K.; Zisserman, A. Very deep convolutional networks for large-scale image recognition. arXiv 2014, arXiv:1409.1556. [Google Scholar] [CrossRef]
- He, K.; Zhang, X.; Ren, S.; Sun, J. Deep residual learning for image recognition. In Proceedings of the IEEE Conference on Computer Vision and Pattern Recognition, Las Vegas, NV, USA, 27–30 June 2016; pp. 770–778. [Google Scholar]
- Huang, G.; Liu, Z.; Van Der Maaten, L.; Weinberger, K.Q. Densely connected convolutional networks. In Proceedings of the IEEE Conference on Computer Vision and Pattern Recognition, Honolulu, HI, USA, 21–26 July 2017; pp. 4700–4708. [Google Scholar]
- Everingham, M.; Van Gool, L.; Williams, C.K.; Winn, J.; Zisserman, A. The pascal visual object classes (voc) challenge. Int. J. Comput. Vis. 2010, 88, 303–338. [Google Scholar] [CrossRef]
- Russakovsky, O.; Deng, J.; Su, H.; Krause, J.; Satheesh, S.; Ma, S.; Huang, Z.; Karpathy, A.; Khosla, A.; Bernstein, M. Imagenet large scale visual recognition challenge. Int. J. Comput. Vis. 2015, 115, 211–252. [Google Scholar] [CrossRef]
- Hong, S.; Han, Y.; Kim, S.; Lee, A.; Kim, G. Application of deep-learning methods to bird detection using unmanned aerial vehicle imagery. Sensors 2019, 19, 1651. [Google Scholar] [CrossRef]
- Dodge, S.; Karam, L. Understanding how image quality affects deep neural networks. In Proceedings of the Eighth International Conference on Quality of Multimedia Experience (QoMEX), Lisbon, Portugal, 6–8 June 2016; pp. 1–6. [Google Scholar]
- Spiesman, B.J.; Gratton, C.; Hatfield, R.G.; Hsu, W.H.; Jepsen, S.; McCornack, B.; Patel, K.; Wang, G. Assessing the potential for deep learning and computer vision to identify bumble bee species from images. Sci. Rep. 2021, 11, 7580. [Google Scholar] [CrossRef]
- Hou, J.; He, Y.; Yang, H.; Connor, T.; Gao, J.; Wang, Y.; Zeng, Y.; Zhang, J.; Huang, J.; Zheng, B. Identification of animal individuals using deep learning: A case study of giant panda. Biol. Conserv. 2020, 242, 108414. [Google Scholar] [CrossRef]
- Lin, T.Y.; Roy Chowdhury, A.; Maji, S. Bilinear CNN models for fine-grained visual recognition. In Proceeding of the IEEE International Conference on Computer Vision, Santiago, Chile, 7–13 December 2015; pp. 1449–1457. [Google Scholar]
- Wah, C.; Branson, S.; Welinder, P.; Perona, P.; Belongie, S. The Caltech-UCSD Birds-200–2011 Dataset; Computation & Neural Systems Technical Report, CNS-TR; California Institute of Technology: Pasadena, CA, USA, 2011. [Google Scholar]
- Van Horn, G.; Branson, S.; Farrell, R.; Haber, S.; Barry, J.; Ipeirotis, P.; Perona, P.; Belongie, S. Building a bird recognition app and large scale dataset with citizen scientists: The fine print in fine-grained dataset collection. In Proceedings of the 2015 IEEE Conference on Computer Vision and Pattern Recognition (CVPR), Boston, MA, USA, 7–12 June 2015; pp. 595–604. [Google Scholar]
- Zink, R.M. The role of subspecies in obscuring avian biological diversity and misleading conservation policy. Proc. R. Soc. B Biol. Sci. 2004, 271, 561–564. [Google Scholar] [CrossRef]
- Johnson, R.N. Conservation Genetics and Wildlife Forensics of Birds. In Wildlife Forensics: Methods and Applications; Jane, E.H., John, R.W., Eds.; Wiley-Blackwell: Hoboken, NJ, USA, 2011; pp. 293–325. [Google Scholar]
- Miao, Z.; Liu, Z.; Gaynor, K.M.; Palmer, M.S.; Yu, S.X.; Getz, W.M. Iterative human and automated identification of wildlife images. Nat. Mach. Intell. 2021, 3, 885–895. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Species | Training Set | Validation Set | Test Set |
|---|---|---|---|---|
| 1 | Amazona aestiva | 219 | 46 | 48 |
| 2 | Amazona albifrons | 217 | 46 | 47 |
| 3 | Amazona amazonica | 289 | 62 | 63 |
| 4 | Amazona auropalliata | 215 | 46 | 47 |
| 5 | Amazona autumnalis | 202 | 43 | 44 |
| 6 | Amazona barbadensis | 164 | 35 | 36 |
| 7 | Amazona brasiliensis | 165 | 35 | 36 |
| 8 | Amazona collaria | 78 | 16 | 18 |
| 9 | Amazona dufresniana | 83 | 17 | 19 |
| 10 | Amazona festiva | 95 | 20 | 21 |
| 11 | Amazona finschi | 228 | 48 | 50 |
| 12 | Amazona guatemalae | 84 | 18 | 19 |
| 13 | Amazona guildingii | 95 | 24 | 26 |
| 14 | Amazona leucocephala | 280 | 60 | 61 |
| 15 | Amazona lilacina | 78 | 16 | 18 |
| 16 | Amazona mercenarius | 79 | 16 | 18 |
| 17 | Amazona ochrocephala | 198 | 42 | 44 |
| 18 | Amazona oratrix | 255 | 54 | 56 |
| 19 | Amazona pretrei | 131 | 19 | 21 |
| 20 | Amazona rhodocorytha | 126 | 27 | 28 |
| 21 | Amazona tucumana | 105 | 22 | 23 |
| 22 | Amazona ventralis | 145 | 31 | 32 |
| 23 | Amazona versicolor | 107 | 22 | 24 |
| 24 | Amazona vinacea | 191 | 41 | 42 |
| 25 | Amazona viridigenalis | 180 | 38 | 40 |
| 26 | Amazona vittata | 108 | 23 | 24 |
| Total | 4096 | 867 | 905 | |
| Model | mAP (%) | Inference Time (ms) |
|---|---|---|
| VGGNet16 | 85.9 | 27 |
| ResNet18 | 87.8 | 22 |
| ResNet34 | 87.5 | 25 |
| ResNet50 | 87.2 | 31 |
| DenseNet18 | 87.6 | 31 |
| DenseNet30 | 86.8 | 34 |
| DenseNet50 | 88.6 | 45 |
| DenseNet121 | 88.9 | 48 |
| Predicted Results | |||||||||||||||||||||||||||
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | 13 | 14 | 15 | 16 | 17 | 18 | 19 | 20 | 21 | 22 | 23 | 24 | 25 | 26 | ||
| True Results | 1 | 91.7 | 0.0 | 2.1 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 2.1 | 0.0 | 0.0 | 2.1 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 2.1 |
| 2 | 0.0 | 91.5 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 8.5 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | |
| 3 | 0.0 | 1.6 | 96.8 | 1.6 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | |
| 4 | 2.1 | 0.0 | 0.0 | 83.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 2.1 | 0.0 | 0.0 | 0.0 | 0.0 | 10.6 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 2.1 | 0.0 | 0.0 | |
| 5 | 0.0 | 0.0 | 0.0 | 0.0 | 84.1 | 0.0 | 2.3 | 0.0 | 0.0 | 0.0 | 2.3 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 4.5 | 4.5 | 0.0 | 0.0 | 0.0 | 0.0 | 2.3 | 0.0 | |
| 6 | 2.8 | 0.0 | 0.0 | 0.0 | 0.0 | 80.6 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 2.8 | 13.9 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | |
| 7 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 94.4 | 2.8 | 0.0 | 2.8 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | |
| 8 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 94.4 | 5.6 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | |
| 9 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 100.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | |
| 10 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 100.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | |
| 11 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 2.0 | 0.0 | 0.0 | 2.0 | 82.0 | 0.0 | 0.0 | 0.0 | 4.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 10.0 | 0.0 | |
| 12 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 100.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | |
| 13 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 96.2 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 3.8 | 0.0 | 0.0 | 0.0 | |
| 14 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 1.6 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 93.4 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 3.3 | 0.0 | 0.0 | 1.6 | 0.0 | 0.0 | |
| 15 | 0.0 | 0.0 | 0.0 | 0.0 | 5.6 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 88.9 | 0.0 | 0.0 | 0.0 | 0.0 | 5.6 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | |
| 16 | 0.0 | 0.0 | 0.0 | 11.1 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 5.6 | 0.0 | 0.0 | 0.0 | 83.3 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | |
| 17 | 0.0 | 0.0 | 0.0 | 4.5 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 4.5 | 0.0 | 0.0 | 0.0 | 0.0 | 88.6 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 2.3 | 0.0 | 0.0 | 0.0 | |
| 18 | 0.0 | 1.8 | 0.0 | 0.0 | 0.0 | 3.6 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 92.9 | 0.0 | 1.8 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | |
| 19 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 100.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | |
| 20 | 0.0 | 0.0 | 0.0 | 0.0 | 3.6 | 0.0 | 3.6 | 0.0 | 3.6 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 89.3 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | |
| 21 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 4.3 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 91.3 | 0.0 | 0.0 | 0.0 | 0.0 | 4.3 | |
| 22 | 0.0 | 0.0 | 3.1 | 0.0 | 0.0 | 0.0 | 0.0 | 3.1 | 3.1 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 87.5 | 3.1 | 0.0 | 0.0 | 0.0 | |
| 23 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 4.2 | 0.0 | 0.0 | 0.0 | 0.0 | 4.2 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 87.5 | 0.0 | 4.2 | 0.0 | |
| 24 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 2.4 | 0.0 | 0.0 | 95.2 | 2.4 | 0.0 | |
| 25 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 5.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 2.5 | 2.5 | 0.0 | 0.0 | 0.0 | 90.0 | 0.0 | |
| 26 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 16.7 | 8.3 | 0.0 | 0.0 | 0.0 | 75.0 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, J.-I.; Baek, J.-W.; Kim, C.-B. Image Classification of Amazon Parrots by Deep Learning: A Potentially Useful Tool for Wildlife Conservation. Biology 2022, 11, 1303. https://doi.org/10.3390/biology11091303
Kim J-I, Baek J-W, Kim C-B. Image Classification of Amazon Parrots by Deep Learning: A Potentially Useful Tool for Wildlife Conservation. Biology. 2022; 11(9):1303. https://doi.org/10.3390/biology11091303
Chicago/Turabian StyleKim, Jung-Il, Jong-Won Baek, and Chang-Bae Kim. 2022. "Image Classification of Amazon Parrots by Deep Learning: A Potentially Useful Tool for Wildlife Conservation" Biology 11, no. 9: 1303. https://doi.org/10.3390/biology11091303
APA StyleKim, J. -I., Baek, J. -W., & Kim, C. -B. (2022). Image Classification of Amazon Parrots by Deep Learning: A Potentially Useful Tool for Wildlife Conservation. Biology, 11(9), 1303. https://doi.org/10.3390/biology11091303