
Survival and Interplay of γ-Aminobutyric Acid-Producing Psychobiotic Candidates with the Gut Microbiota in a Continuous Model of the Human Colon




Abstract
:Simple Summary
Abstract
1. Introduction
2. Material and Methods
2.1. Bacterial Strains, Media, and Culture Conditions
2.2. Bacterial Enumeration by Plate Counts
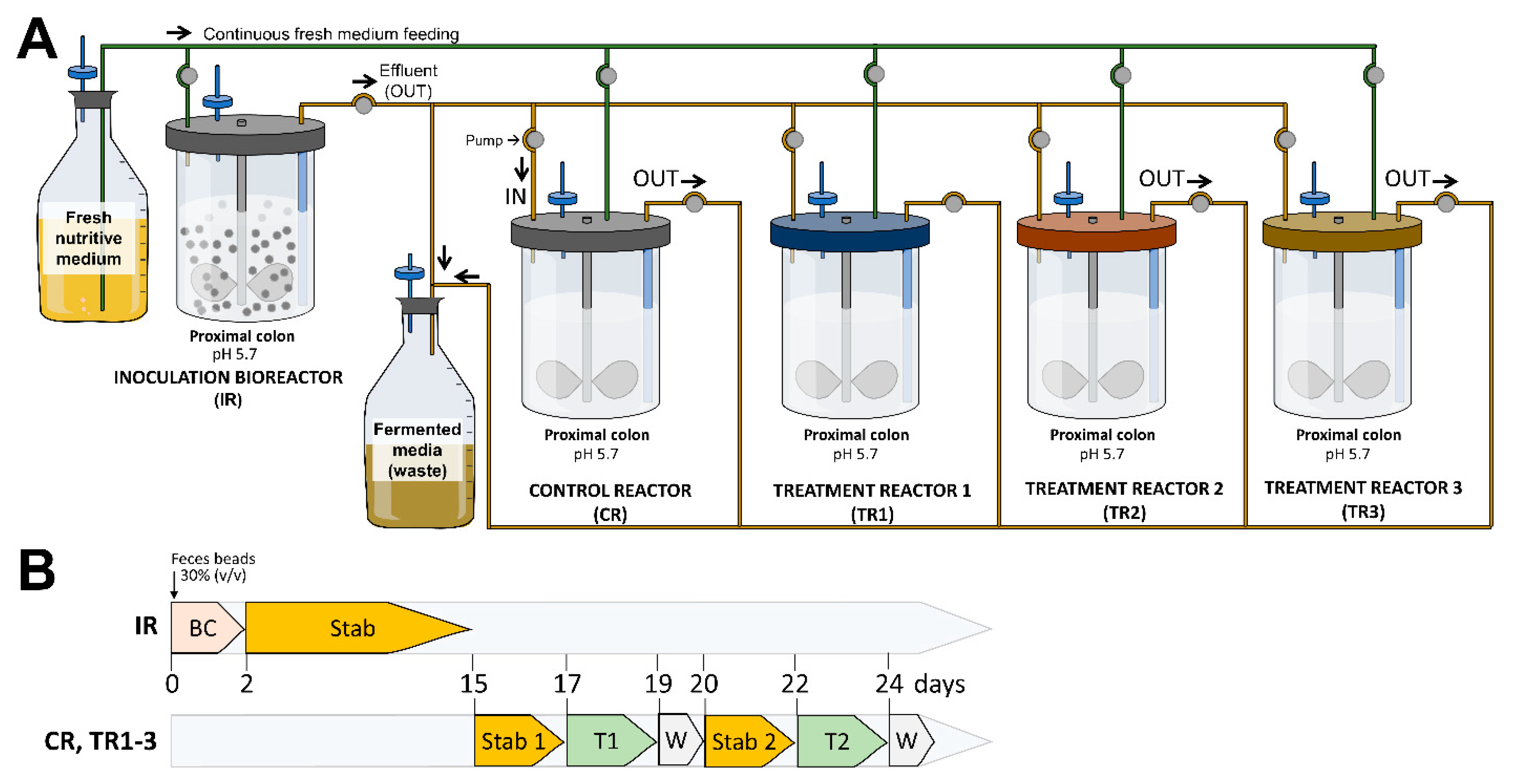
2.3. Human Colonic Fermentation Model
2.3.1. Nutritive Culture Medium
2.3.2. Fecal Sample Collection and Cell Immobilization in Gel Beads
2.3.3. Experimental Setup and Fermentation Procedure
2.4. Microbiota Diversity Analyses
2.4.1. Genomic DNA Extraction
2.4.2. High-Throughput 16S DNA Sequencing
2.4.3. qPCR Analysis
2.5. Determination of Production of SCFAs Using GC
2.6. Statistical Analyses
3. Results
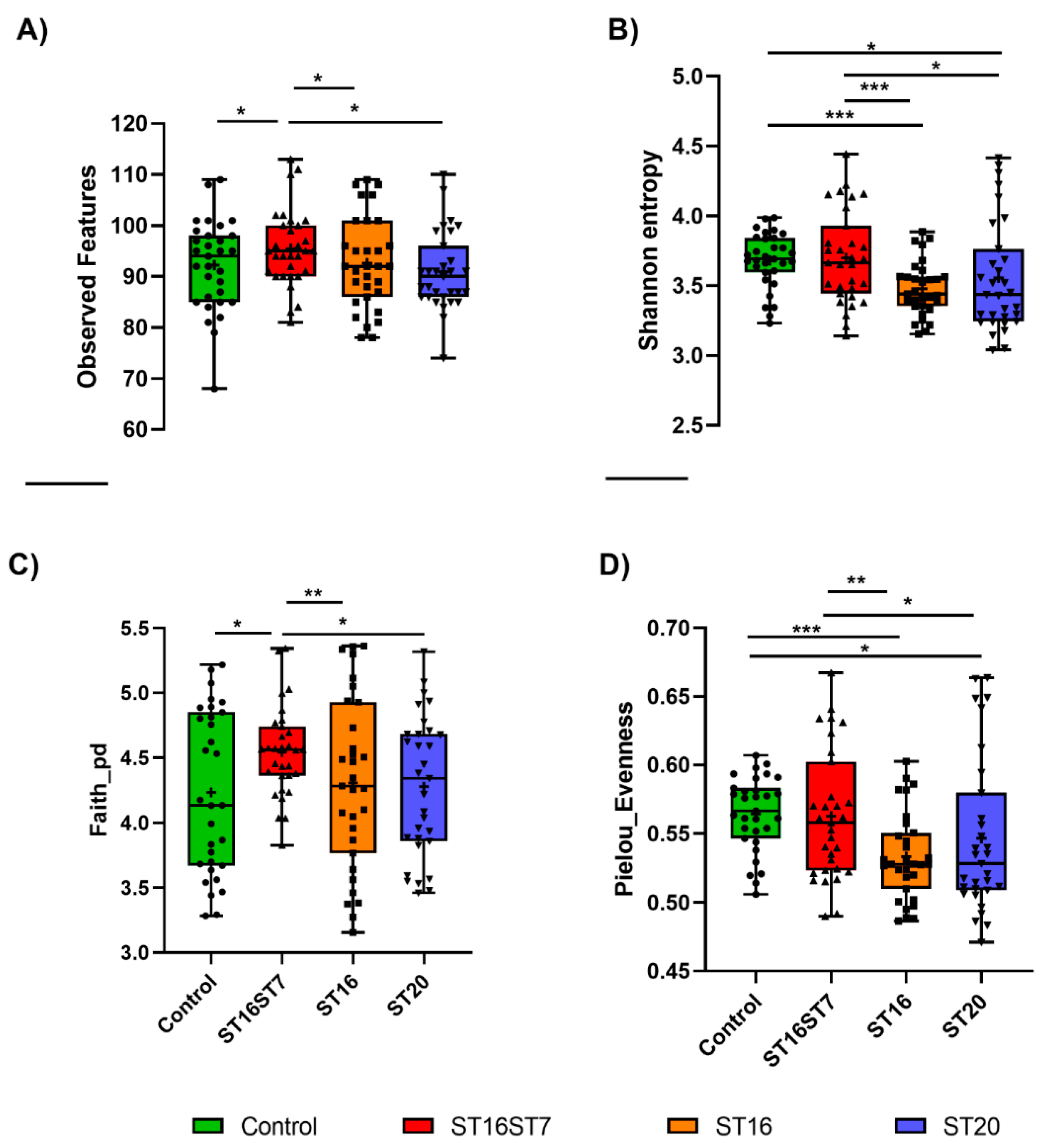
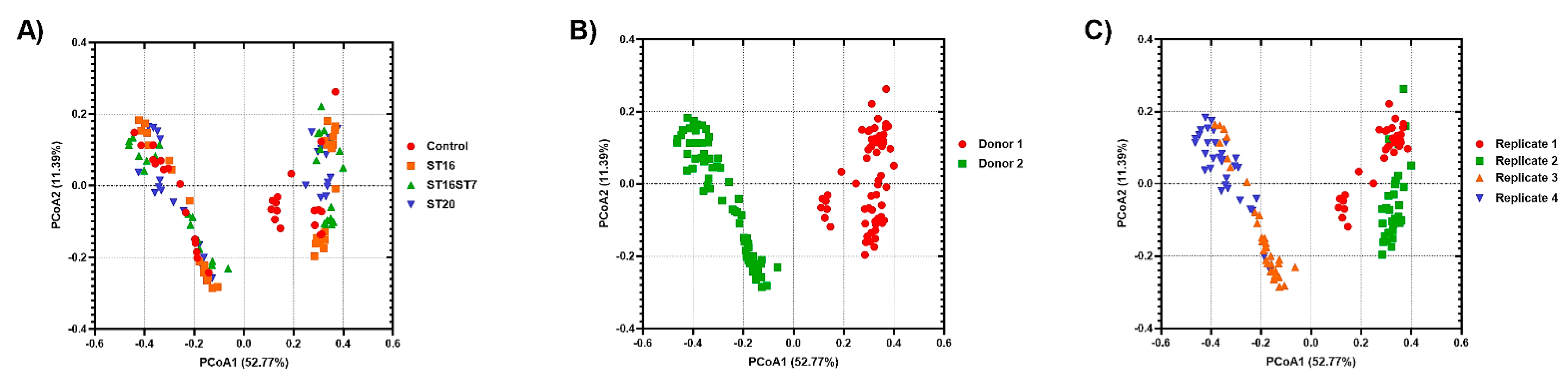
3.1. Gut Microbiota Diversity
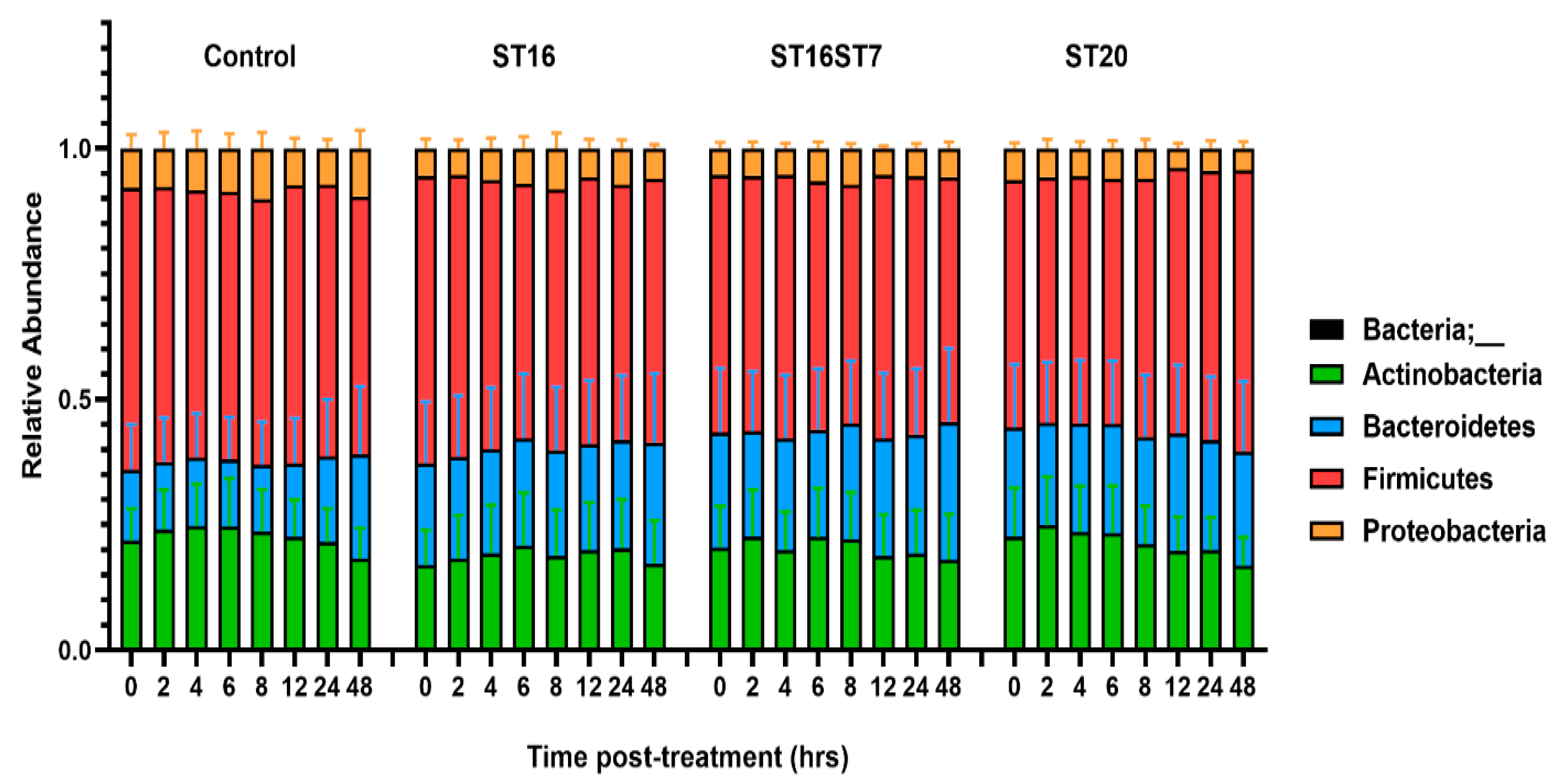
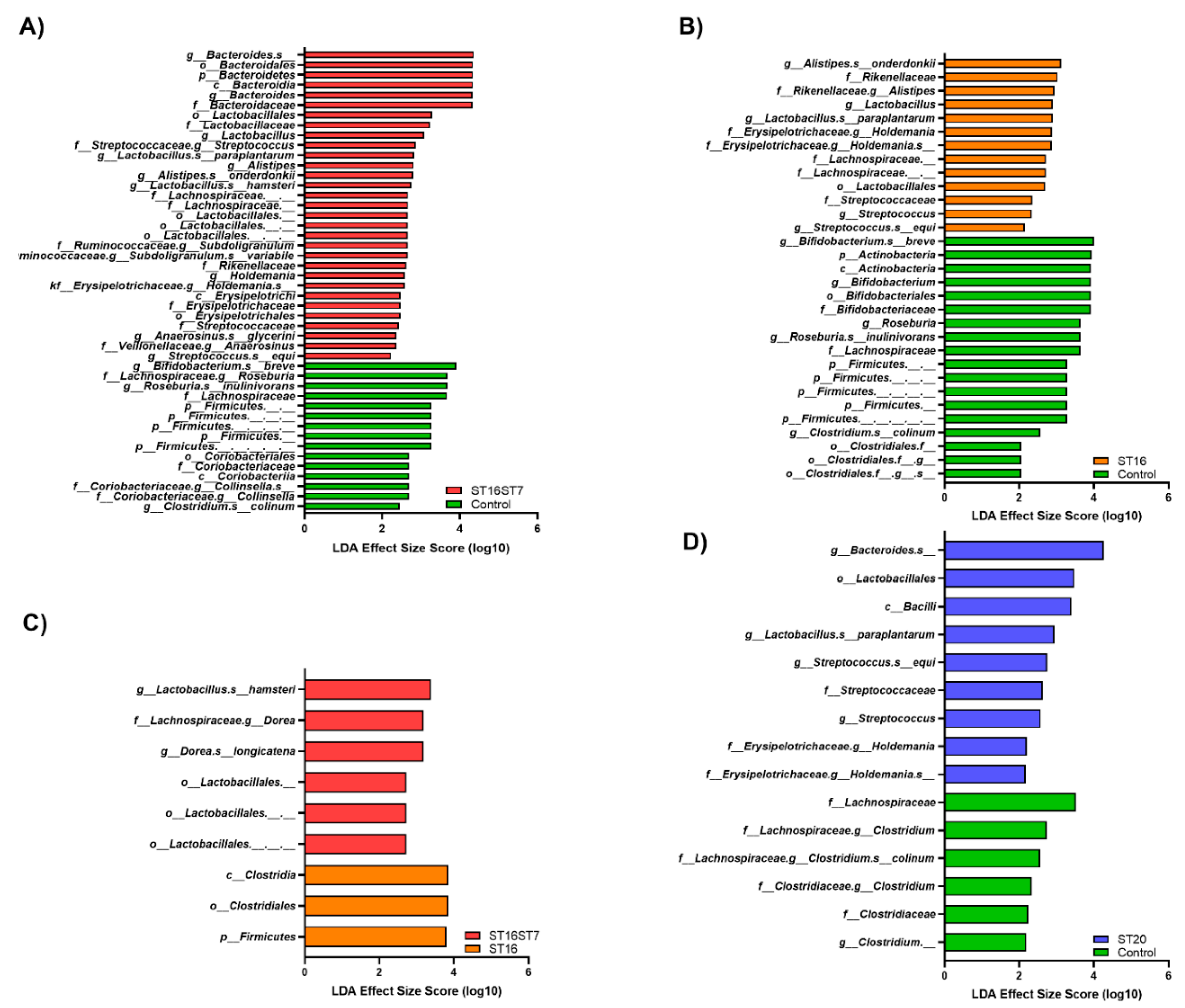
3.2. Effect of GABA-Producing Probiotic Candidates on Gut Microbiota Composition
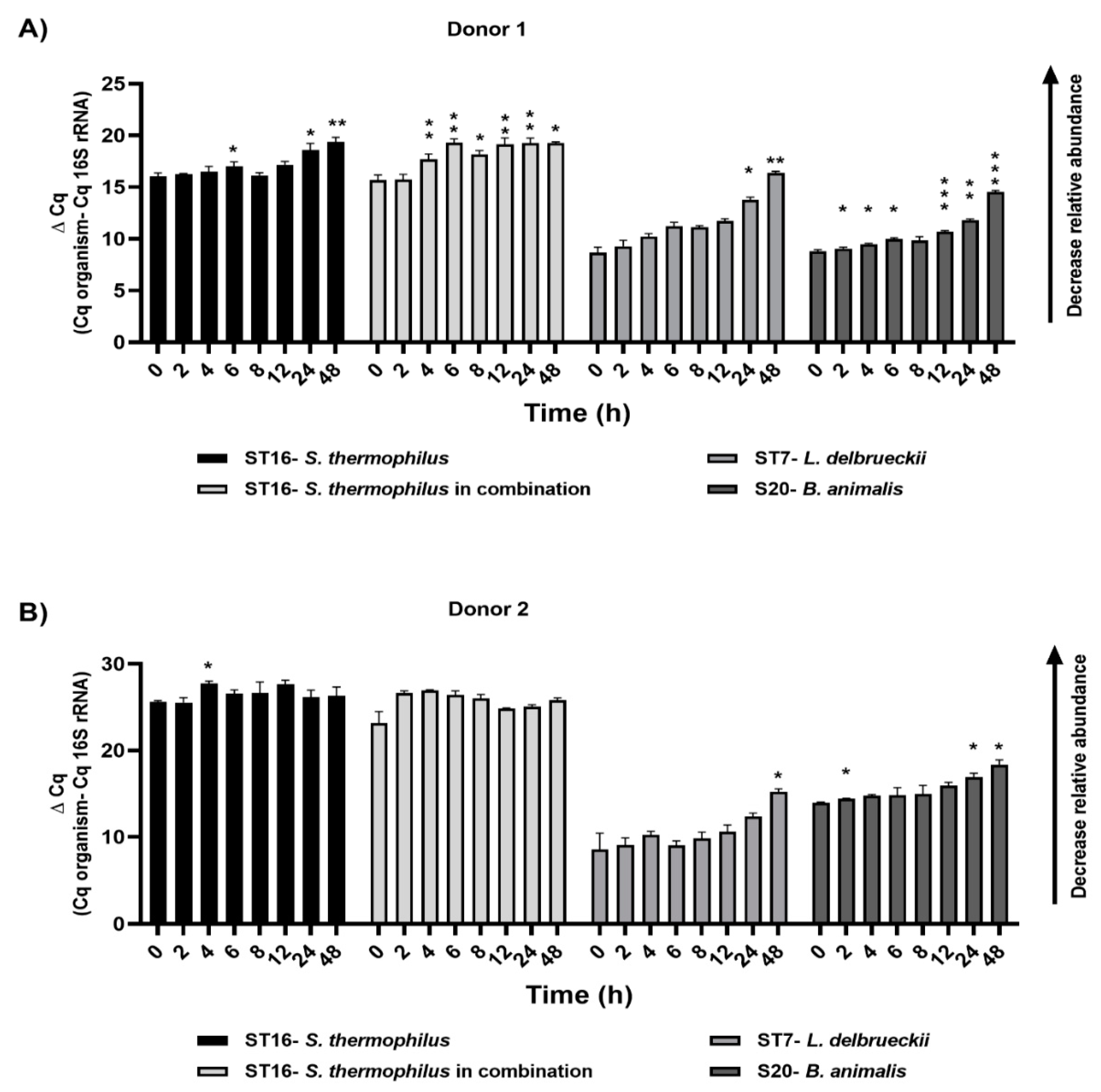
3.3. Microbial Survival Analysis by qPCR
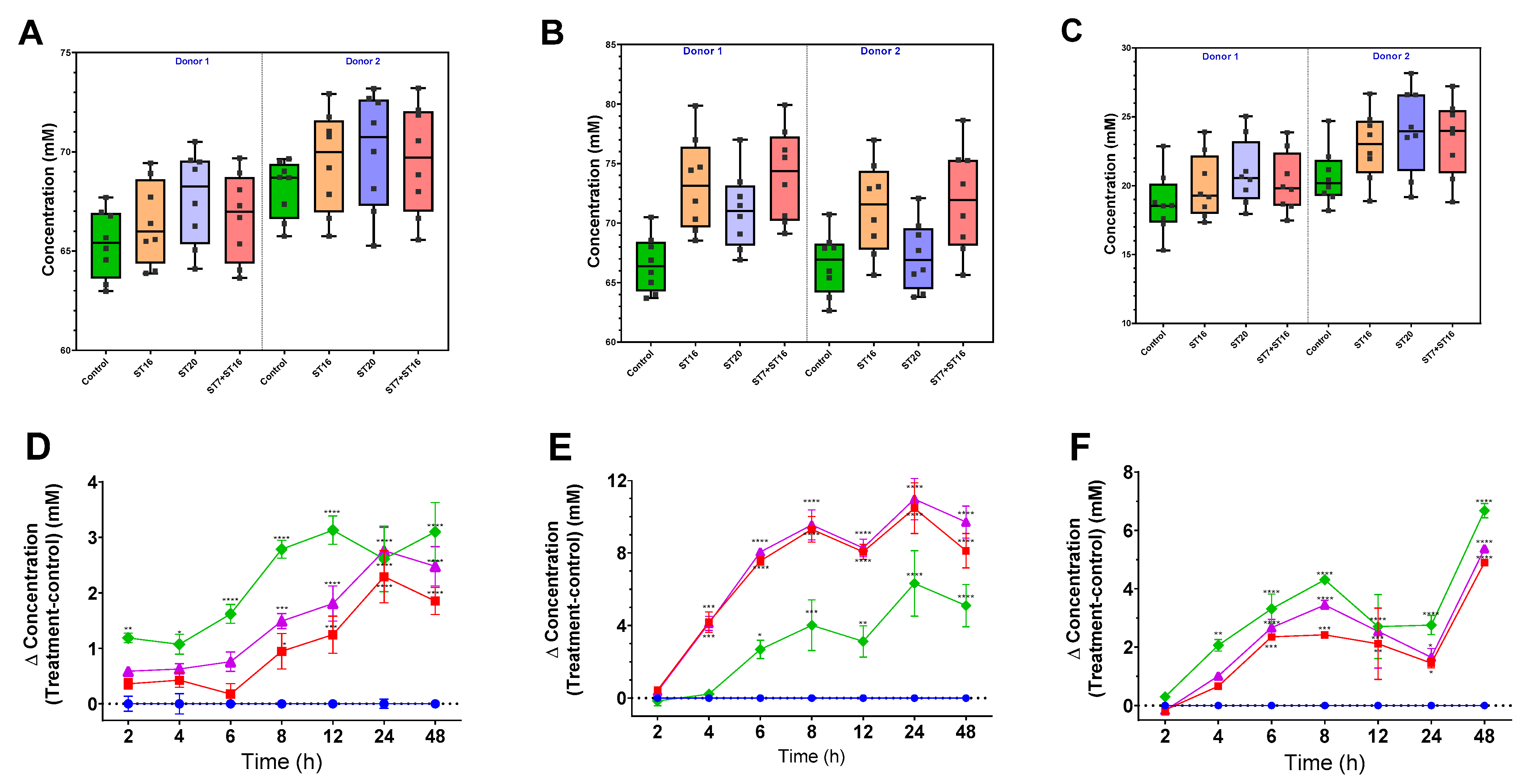
3.4. Effect of GABA-Producing Probiotic Candidates on Microbiota Generation of SCFAs
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Carabotti, M.; Scirocco, A.; Maselli, M.A.; Severi, C. The Gut-Brain Axis: Interactions between Enteric Microbiota, Central and Enteric Nervous Systems. Ann. Gastroenterol. 2015, 28, 203–209. [Google Scholar] [PubMed]
- Ma, Q.; Xing, C.; Long, W.; Wang, H.Y.; Liu, Q.; Wang, R.-F. Impact of Microbiota on Central Nervous System and Neurological Diseases: The Gut-Brain Axis. J. Neuroinflammation 2019, 16, 53. [Google Scholar] [CrossRef]
- Sudo, N.; Chida, Y.; Aiba, Y.; Sonoda, J.; Oyama, N.; Yu, X.-N.; Kubo, C.; Koga, Y. Postnatal Microbial Colonization Programs the Hypothalamic-Pituitary-Adrenal System for Stress Response in Mice: Commensal Microbiota and Stress Response. J. Physiol. 2004, 558, 263–275. [Google Scholar] [CrossRef] [PubMed]
- Huo, R.; Zeng, B.; Zeng, L.; Cheng, K.; Li, B.; Luo, Y.; Wang, H.; Zhou, C.; Fang, L.; Li, W.; et al. Microbiota Modulate Anxiety-Like Behavior and Endocrine Abnormalities in Hypothalamic-Pituitary-Adrenal Axis. Front. Cell. Infect. Microbiol. 2017, 7, 489. [Google Scholar] [CrossRef] [PubMed]
- Sarkar, A.; Lehto, S.M.; Harty, S.; Dinan, T.G.; Cryan, J.F.; Burnet, P.W.J. Psychobiotics and the Manipulation of Bacteria-Gut-Brain Signals. Trends Neurosci. 2016, 39, 763–781. [Google Scholar] [CrossRef]
- Hill, C.; Guarner, F.; Reid, G.; Gibson, G.R.; Merenstein, D.J.; Pot, B.; Morelli, L.; Canani, R.B.; Flint, H.J.; Salminen, S.; et al. The International Scientific Association for Probiotics and Prebiotics Consensus Statement on the Scope and Appropriate Use of the Term Probiotic. Nat. Rev. Gastroenterol. Hepatol. 2014, 11, 506–514. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.; Sun, J.; Li, G.; Zhang, M.; Niu, T.; Kang, X.; Zhao, H.; Chen, J.; Sun, E.; Li, Y. Effect of Bifidobacterium Animalis Subsp. Lactis MN-Gup on Constipation and the Composition of Gut Microbiota. Benef. Microbes 2021, 12, 31–42. [Google Scholar] [CrossRef]
- Matricardi, P.M. Probiotics against Allergy: Data, Doubts, and Perspectives. Allergy 2002, 57, 185–187. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Yin, Y.; Xiao, D.; Zou, Y. Supplemental Bifid Triple Viable Capsule Treatment Improves Inflammatory Response and T Cell Frequency in Ulcerative Colitis Patients. BMC Gastroenterol. 2021, 21, 314. [Google Scholar] [CrossRef] [PubMed]
- Saviano, A.; Brigida, M.; Migneco, A.; Gunawardena, G.; Zanza, C.; Candelli, M.; Franceschi, F.; Ojetti, V. Lactobacillus Reuteri DSM 17938 (Limosilactobacillus Reuteri) in Diarrhea and Constipation: Two Sides of the Same Coin? Med. Kaunas Lith. 2021, 57, 643. [Google Scholar] [CrossRef] [PubMed]
- Xu, H.-L.; Zou, L.-L.; Chen, M.-B.; Wang, H.; Shen, W.-M.; Zheng, Q.-H.; Cui, W.-Y. Efficacy of Probiotic Adjuvant Therapy for Irritable Bowel Syndrome in Children: A Systematic Review and Meta-Analysis. PLoS ONE 2021, 16, e0255160. [Google Scholar] [CrossRef] [PubMed]
- Ansari, F.; Pourjafar, H.; Tabrizi, A.; Homayouni, A. The Effects of Probiotics and Prebiotics on Mental Disorders: A Review on Depression, Anxiety, Alzheimer, and Autism Spectrum Disorders. Curr. Pharm. Biotechnol. 2020, 21, 555–565. [Google Scholar] [CrossRef]
- Yong, S.J.; Tong, T.; Chew, J.; Lim, W.L. Antidepressive Mechanisms of Probiotics and Their Therapeutic Potential. Front. Neurosci. 2020, 13, 1361. [Google Scholar] [CrossRef] [PubMed]
- van de Wouw, M.; Boehme, M.; Lyte, J.M.; Wiley, N.; Strain, C.; O’Sullivan, O.; Clarke, G.; Stanton, C.; Dinan, T.G.; Cryan, J.F. Short-Chain Fatty Acids: Microbial Metabolites That Alleviate Stress-Induced Brain-Gut Axis Alterations. J. Physiol. 2018, 596, 4923–4944. [Google Scholar] [CrossRef]
- Dinan, T.G.; Butler, M.I.; Cryan, J.F. Psychobiotics: Evolution of Novel Antidepressants. In Modern Trends in Psychiatry; Cowan, C.S.M., Leonard, B.E., Eds.; S. Karger AG: Basel, Switzerland, 2021; Volume 32, pp. 134–143. ISBN 978-3-318-06855-9. [Google Scholar]
- Bercik, P.; Park, A.J.; Sinclair, D.; Khoshdel, A.; Lu, J.; Huang, X.; Deng, Y.; Blennerhassett, P.A.; Fahnestock, M.; Moine, D.; et al. The Anxiolytic Effect of Bifidobacterium Longum NCC3001 Involves Vagal Pathways for Gut-Brain Communication. Neurogastroenterol. Motil. Off. J. Eur. Gastrointest. Motil. Soc. 2011, 23, 1132–1139. [Google Scholar] [CrossRef] [PubMed]
- Kunze, W.A.; Mao, Y.-K.; Wang, B.; Huizinga, J.D.; Ma, X.; Forsythe, P.; Bienenstock, J. Lactobacillus Reuteri Enhances Excitability of Colonic AH Neurons by Inhibiting Calcium-Dependent Potassium Channel Opening. J. Cell. Mol. Med. 2009, 13, 2261–2270. [Google Scholar] [CrossRef] [PubMed]
- Cryan, J.F.; Dinan, T.G. Mind-Altering Microorganisms: The Impact of the Gut Microbiota on Brain and Behaviour. Nat. Rev. Neurosci. 2012, 13, 701–712. [Google Scholar] [CrossRef] [PubMed]
- Dhakal, R.; Bajpai, V.K.; Baek, K.-H. Production of Gaba (γ-Aminobutyric Acid) by Microorganisms: A Review. Braz. J. Microbiol. Publ. Braz. Soc. Microbiol. 2012, 43, 1230–1241. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Cao, Y. Lactic Acid Bacterial Cell Factories for Gamma-Aminobutyric Acid. Amino Acids 2010, 39, 1107–1116. [Google Scholar] [CrossRef]
- Strandwitz, P.; Kim, K.H.; Terekhova, D.; Liu, J.K.; Sharma, A.; Levering, J.; McDonald, D.; Dietrich, D.; Ramadhar, T.R.; Lekbua, A.; et al. GABA-Modulating Bacteria of the Human Gut Microbiota. Nat. Microbiol. 2019, 4, 396–403. [Google Scholar] [CrossRef] [PubMed]
- Mayer, E.A.; Knight, R.; Mazmanian, S.K.; Cryan, J.F.; Tillisch, K. Gut Microbes and the Brain: Paradigm Shift in Neuroscience. J. Neurosci. 2014, 34, 15490–15496. [Google Scholar] [CrossRef] [PubMed]
- Dinan, T.G.; Cryan, J.F. Microbes, Immunity and Behavior: Psychoneuroimmunology Meets the Microbiome. Neuropsychopharmacology 2017, 42, 178–192. [Google Scholar] [CrossRef] [PubMed]
- Guzman-Rodriguez, M.; McDonald, J.A.K.; Hyde, R.; Allen-Vercoe, E.; Claud, E.C.; Sheth, P.M.; Petrof, E.O. Using Bioreactors to Study the Effects of Drugs on the Human Microbiota. Methods 2018, 149, 31–41. [Google Scholar] [CrossRef] [PubMed]
- Mousavi, R.; Mottawea, W.; Hassan, H.; Gomaa, A.; Audet, M.-C.; Hammami, R. Screening, Characterization, and Growth of γ-Aminobutyric Acid-Producing Probiotic Candidates from Food Origin under Simulated Colonic Conditions. J. Appl. Microbiol. 2022, 132, 4452–4465. [Google Scholar] [CrossRef] [PubMed]
- Macfarlane, G.T.; Macfarlane, S.; Gibson, G.R. Validation of a Three-Stage Compound Continuous Culture System for Investigating the Effect of Retention Time on the Ecology and Metabolism of Bacteria in the Human Colon. Microb. Ecol. 1998, 35, 180–187. [Google Scholar] [CrossRef] [PubMed]
- Mottawea, W.; Sultan, S.; Landau, K.; Bordenave, N.; Hammami, R. Evaluation of the Prebiotic Potential of a Commercial Synbiotic Food Ingredient on Gut Microbiota in an Ex Vivo Model of the Human Colon. Nutrients 2020, 12, 2669. [Google Scholar] [CrossRef]
- Le Blay, G.; Hammami, R.; Lacroix, C.; Fliss, I. Stability and Inhibitory Activity of Pediocin PA-1 Against Listeria Sp. in Simulated Physiological Conditions of the Human Terminal Ileum. Probiotics Antimicrob. Proteins 2012, 4, 250–258. [Google Scholar] [CrossRef] [PubMed]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Peña, A.G.; Goodrich, J.K.; Gordon, J.I.; et al. QIIME Allows Analysis of High-Throughput Community Sequencing Data. Nat. Methods 2010, 7, 335–336. [Google Scholar] [CrossRef] [PubMed]
- Fernandez, B.; Savard, P.; Fliss, I. Survival and Metabolic Activity of Pediocin Producer Pediococcus Acidilactici UL5: Its Impact on Intestinal Microbiota and Listeria Monocytogenes in a Model of the Human Terminal Ileum. Microb. Ecol. 2016, 72, 931–942. [Google Scholar] [CrossRef]
- Byun, R.; Nadkarni, M.A.; Chhour, K.-L.; Martin, F.E.; Jacques, N.A.; Hunter, N. Quantitative Analysis of Diverse Lactobacillus Species Present in Advanced Dental Caries. J. Clin. Microbiol. 2004, 42, 3128–3136. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stachelska, M.A. Identification of Lactobacillus Delbrueckii and Streptococcus Thermophilus Strains Present in Artisanal Raw Cow Milk Cheese Using Real-Time PCR and Classic Plate Count Methods. Pol. J. Microbiol. 2017, 66, 491–499. [Google Scholar] [CrossRef] [PubMed]
- Kwon, H.-S.; Yang, E.-H.; Lee, S.-H.; Yeon, S.-W.; Kang, B.-H.; Kim, T.-Y. Rapid Identification of Potentially Probiotic Bifidobacterium Species by Multiplex PCR Using Species-Specific Primers Based on the Region Extending from 16S RRNA through 23S RRNA. FEMS Microbiol. Lett. 2005, 250, 55–62. [Google Scholar] [CrossRef] [PubMed]
- Monk, J.M.; Zhang, C.P.; Wu, W.; Zarepoor, L.; Lu, J.T.; Liu, R.; Pauls, K.P.; Wood, G.A.; Tsao, R.; Robinson, L.E.; et al. White and Dark Kidney Beans Reduce Colonic Mucosal Damage and Inflammation in Response to Dextran Sodium Sulfate. J. Nutr. Biochem. 2015, 26, 752–760. [Google Scholar] [CrossRef] [PubMed]
- Bolyen, E.; Rideout, J.R.; Dillon, M.R.; Bokulich, N.A.; Abnet, C.C.; Al-Ghalith, G.A.; Alexander, H.; Alm, E.J.; Arumugam, M.; Asnicar, F.; et al. Reproducible, Interactive, Scalable and Extensible Microbiome Data Science Using QIIME 2. Nat. Biotechnol. 2019, 37, 852–857. [Google Scholar] [CrossRef]
- Segata, N.; Izard, J.; Waldron, L.; Gevers, D.; Miropolsky, L.; Garrett, W.S.; Huttenhower, C. Metagenomic Biomarker Discovery and Explanation. Genome Biol. 2011, 12, R60. [Google Scholar] [CrossRef]
- Mottawea, W.; Chiang, C.-K.; Mühlbauer, M.; Starr, A.E.; Butcher, J.; Abujamel, T.; Deeke, S.A.; Brandel, A.; Zhou, H.; Shokralla, S.; et al. Altered Intestinal Microbiota–Host Mitochondria Crosstalk in New Onset Crohn’s Disease. Nat. Commun. 2016, 7, 13419. [Google Scholar] [CrossRef]
- Mottawea, W.; Butcher, J.; Li, J.; Abujamel, T.; Manoogian, J.; Mack, D.; Stintzi, A. The Mucosal–Luminal Interface: An Ideal Sample to Study the Mucosa-Associated Microbiota and the Intestinal Microbial Biogeography. Pediatr. Res. 2019, 85, 895–903. [Google Scholar] [CrossRef]
- Poeker, S.A.; Geirnaert, A.; Berchtold, L.; Greppi, A.; Krych, L.; Steinert, R.E.; de Wouters, T.; Lacroix, C. Understanding the Prebiotic Potential of Different Dietary Fibers Using an in Vitro Continuous Adult Fermentation Model (PolyFermS). Sci. Rep. 2018, 8, 4318. [Google Scholar] [CrossRef]
- Winter, G.; Hart, R.A.; Charlesworth, R.P.G.; Sharpley, C.F. Gut Microbiome and Depression: What We Know and What We Need to Know. Rev. Neurosci. 2018, 29, 629–643. [Google Scholar] [CrossRef]
- Zheng, P.; Zeng, B.; Zhou, C.; Liu, M.; Fang, Z.; Xu, X.; Zeng, L.; Chen, J.; Fan, S.; Du, X.; et al. Gut Microbiome Remodeling Induces Depressive-like Behaviors through a Pathway Mediated by the Host’s Metabolism. Mol. Psychiatry 2016, 21, 786–796. [Google Scholar] [CrossRef]
- Bengesser, S.A.; Mörkl, S.; Painold, A.; Dalkner, N.; Birner, A.; Fellendorf, F.T.; Platzer, M.; Queissner, R.; Hamm, C.; Maget, A.; et al. Epigenetics of the Molecular Clock and Bacterial Diversity in Bipolar Disorder. Psychoneuroendocrinology 2019, 101, 160–166. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Wu, X.; Xia, Y.; Wang, M.; Liao, S.; Li, F.; Yin, J.; Ren, W.; Tan, B.; Yin, Y. Effects of Dietary Gamma-Aminobutyric Acid Supplementation on Amino Acid Profile, Intestinal Immunity, and Microbiota in ETEC-Challenged Piglets. Food Funct. 2020, 11, 9067–9074. [Google Scholar] [CrossRef] [PubMed]
- Shi, Y.; Zhao, X.; Zhao, J.; Zhang, H.; Zhai, Q.; Narbad, A.; Chen, W. A Mixture of Lactobacillus Species Isolated from Traditional Fermented Foods Promote Recovery from Antibiotic-Induced Intestinal Disruption in Mice. J. Appl. Microbiol. 2018, 124, 842–854. [Google Scholar] [CrossRef] [PubMed]
- Shi, Y.; Zhai, Q.; Li, D.; Mao, B.; Liu, X.; Zhao, J.; Zhang, H.; Chen, W. Restoration of Cefixime-Induced Gut Microbiota Changes by Lactobacillus Cocktails and Fructooligosaccharides in a Mouse Model. Microbiol. Res. 2017, 200, 14–24. [Google Scholar] [CrossRef] [PubMed]
- Han, F.; Wu, G.; Zhang, Y.; Zheng, H.; Han, S.; Li, X.; Cai, W.; Liu, J.; Zhang, W.; Zhang, X.; et al. Streptococcus Thermophilus Attenuates Inflammation in Septic Mice Mediated by Gut Microbiota. Front. Microbiol. 2020, 11. [Google Scholar] [CrossRef]
- Veljović, K.; Dinić, M.; Lukić, J.; Mihajlović, S.; Tolinački, M.; Živković, M.; Begović, J.; Mrvaljević, I.; Golić, N.; Terzić-Vidojević, A. Promotion of Early Gut Colonization by Probiotic Intervention on Microbiota Diversity in Pregnant Sows. Front. Microbiol. 2017, 8, 2028. [Google Scholar] [CrossRef]
- Fehlbaum, S.; Prudence, K.; Kieboom, J.; Heerikhuisen, M.; van den Broek, T.; Schuren, F.; Steinert, R.; Raederstorff, D. In Vitro Fermentation of Selected Prebiotics and Their Effects on the Composition and Activity of the Adult Gut Microbiota. Int. J. Mol. Sci. 2018, 19, 3097. [Google Scholar] [CrossRef]
- Umu, Ö.C.O.; Rudi, K.; Diep, D.B. Modulation of the Gut Microbiota by Prebiotic Fibres and Bacteriocins. Microb. Ecol. Health Dis. 2017, 28, 1348886. [Google Scholar] [CrossRef]
- Lukić, I.; Getselter, D.; Ziv, O.; Oron, O.; Reuveni, E.; Koren, O.; Elliott, E. Antidepressants Affect Gut Microbiota and Ruminococcus Flavefaciens Is Able to Abolish Their Effects on Depressive-like Behavior. Transl. Psychiatry 2019, 9, 133. [Google Scholar] [CrossRef]
- Zheng, P.; Yang, J.; Li, Y.; Wu, J.; Liang, W.; Yin, B.; Tan, X.; Huang, Y.; Chai, T.; Zhang, H.; et al. Gut Microbial Signatures Can Discriminate Unipolar from Bipolar Depression. Adv. Sci. Weinh. Baden-Wurtt. Ger. 2020, 7, 1902862. [Google Scholar] [CrossRef]
- Luo, M.; Zhuang, X.; Tian, Z.; Xiong, L. Alterations in Short-Chain Fatty Acids and Serotonin in Irritable Bowel Syndrome: A Systematic Review and Meta-Analysis. BMC Gastroenterol. 2021, 21, 14. [Google Scholar] [CrossRef]
- Parada Venegas, D.; De la Fuente, M.K.; Landskron, G.; González, M.J.; Quera, R.; Dijkstra, G.; Harmsen, H.J.M.; Faber, K.N.; Hermoso, M.A. Short Chain Fatty Acids (SCFAs)-Mediated Gut Epithelial and Immune Regulation and Its Relevance for Inflammatory Bowel Diseases. Front. Immunol. 2019, 10, 277. [Google Scholar] [CrossRef] [PubMed]
- Tan, J.; McKenzie, C.; Potamitis, M.; Thorburn, A.N.; Mackay, C.R.; Macia, L. Chapter Three-The Role of Short-Chain Fatty Acids in Health and Disease. In Advances in Immunology; Alt, F.W., Ed.; Academic Press: Cambridge, MA, USA, 2014; Volume 121, pp. 91–119. [Google Scholar]
- Lavoie, S.; Chun, E.; Bae, S.; Brennan, C.A.; Gallini Comeau, C.A.; Lang, J.K.; Michaud, M.; Hoveyda, H.R.; Fraser, G.L.; Fuller, M.H.; et al. Expression of Free Fatty Acid Receptor 2 by Dendritic Cells Prevents Their Expression of Interleukin 27 and Is Required for Maintenance of Mucosal Barrier and Immune Response Against Colorectal Tumors in Mice. Gastroenterology 2020, 158, 1359–1372.e9. [Google Scholar] [CrossRef] [PubMed]
- Liśkiewicz, P.; Kaczmarczyk, M.; Misiak, B.; Wroński, M.; Bąba-Kubiś, A.; Skonieczna-Żydecka, K.; Marlicz, W.; Bieńkowski, P.; Misera, A.; Pełka-Wysiecka, J.; et al. Analysis of Gut Microbiota and Intestinal Integrity Markers of Inpatients with Major Depressive Disorder. Prog. Neuropsychopharmacol. Biol. Psychiatry 2020, 106, 110076. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No | Specificity | Primer Name | Primer Type | Sequence (5′-3′) | GC Content (%) | Melting Temp (°C) | Ref. |
|---|---|---|---|---|---|---|---|
| 1 | Lactobacillus delbrueckii subsp. bulgaricus | LdelbF | Forward | GGRTGATTTGTTGGACGCTAG | 47.6 | 66.9 | [31] |
| LdelbR | Reverse | GCCGCCTTTCAAACTTGAATC | 47.6 | 66. 7 | |||
| 2 | Streptococcus thermophilus | S. thermophilusF | Forward | TTATTTGAAAGGGGCAATTGCT | 36.3 | 65.2 | [32] |
| S. thermophilusR | Reverse | GTGAACTTTCCACTCTCACAC | 47.6 | 58.8 | |||
| 3 | Bifidobacterium animalis | IDB61F | Forward | GCATGTTGCCAGCGGGTGA | 63.1 | 73.1 | [33] |
| IDBC1R | Reverse | ATCCGAACTGAGACCGGTT | 52.6 | 63.5 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mousavi, R.; Mottawea, W.; Audet, M.-C.; Hammami, R. Survival and Interplay of γ-Aminobutyric Acid-Producing Psychobiotic Candidates with the Gut Microbiota in a Continuous Model of the Human Colon. Biology 2022, 11, 1311. https://doi.org/10.3390/biology11091311
Mousavi R, Mottawea W, Audet M-C, Hammami R. Survival and Interplay of γ-Aminobutyric Acid-Producing Psychobiotic Candidates with the Gut Microbiota in a Continuous Model of the Human Colon. Biology. 2022; 11(9):1311. https://doi.org/10.3390/biology11091311
Chicago/Turabian StyleMousavi, Rojaalsadat, Walid Mottawea, Marie-Claude Audet, and Riadh Hammami. 2022. "Survival and Interplay of γ-Aminobutyric Acid-Producing Psychobiotic Candidates with the Gut Microbiota in a Continuous Model of the Human Colon" Biology 11, no. 9: 1311. https://doi.org/10.3390/biology11091311
APA StyleMousavi, R., Mottawea, W., Audet, M. -C., & Hammami, R. (2022). Survival and Interplay of γ-Aminobutyric Acid-Producing Psychobiotic Candidates with the Gut Microbiota in a Continuous Model of the Human Colon. Biology, 11(9), 1311. https://doi.org/10.3390/biology11091311