
Cumulus Cell and Oocyte Gene Expression in Prepubertal Gilts and Sows Identifies Cumulus Cells as a Prime Informative Parameter of Oocyte Quality
 , , ,
, , , 
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Chemicals and Media
2.2. Animal Material and Ethics
2.3. Experimental Design
2.4. Selection of Target Genes
2.5. Cumulus–Oocyte Complex Collection and In Vitro Maturation
2.6. RNA Extraction and cDNA Synthesis
2.7. Quantitative PCR
2.8. Statistical Analysis
3. Results
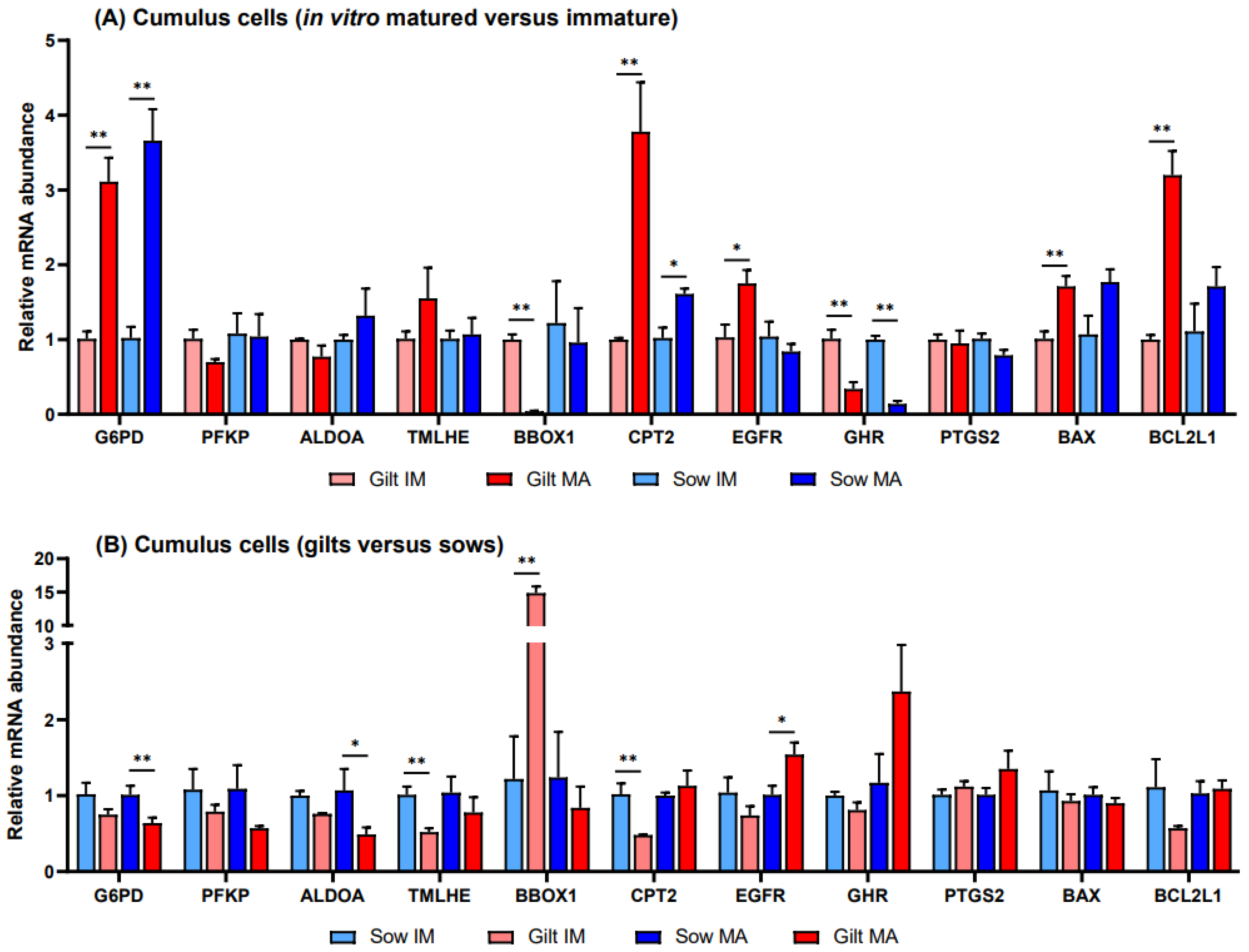
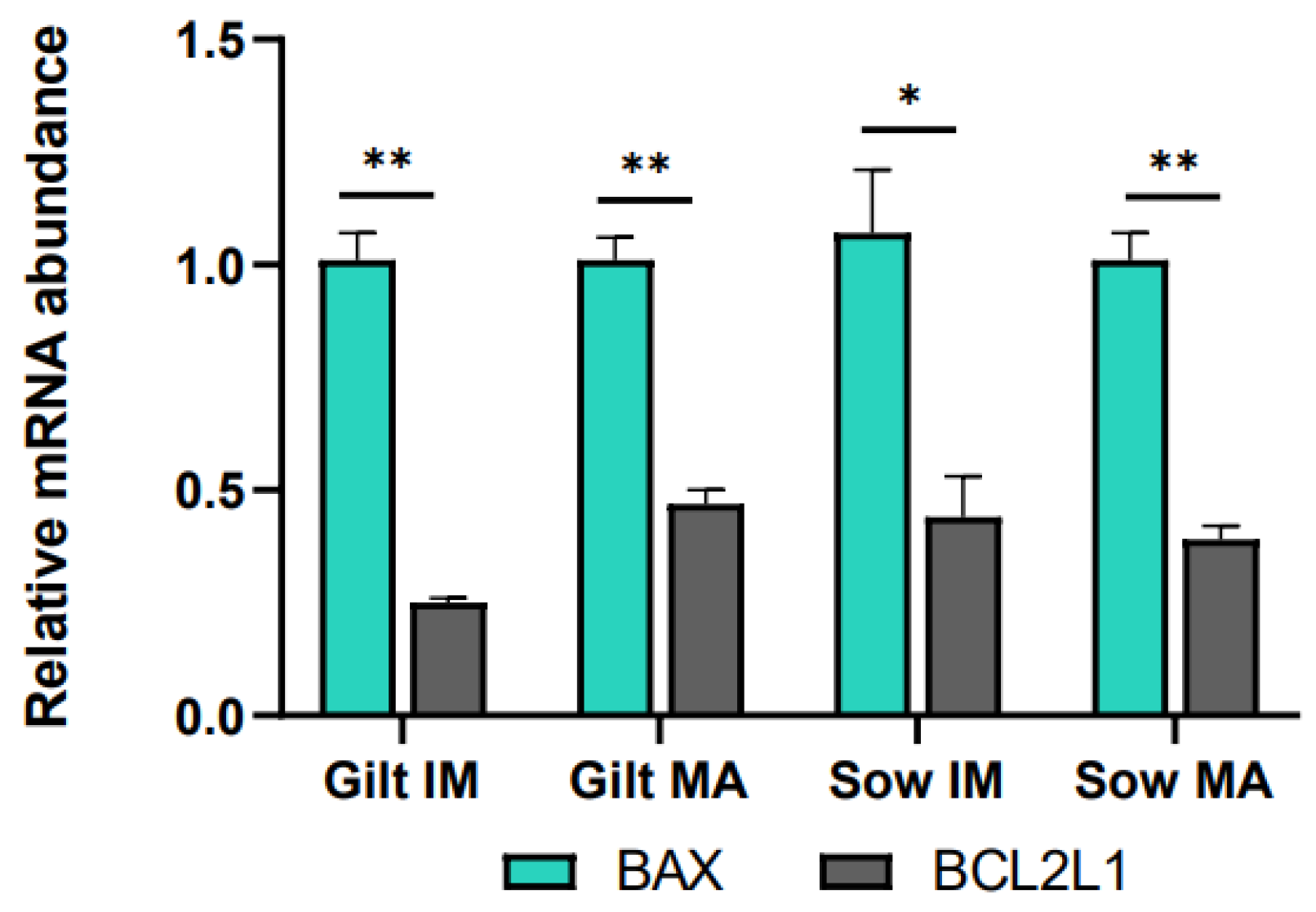
3.1. Gene Expression in Porcine Cumulus Cells
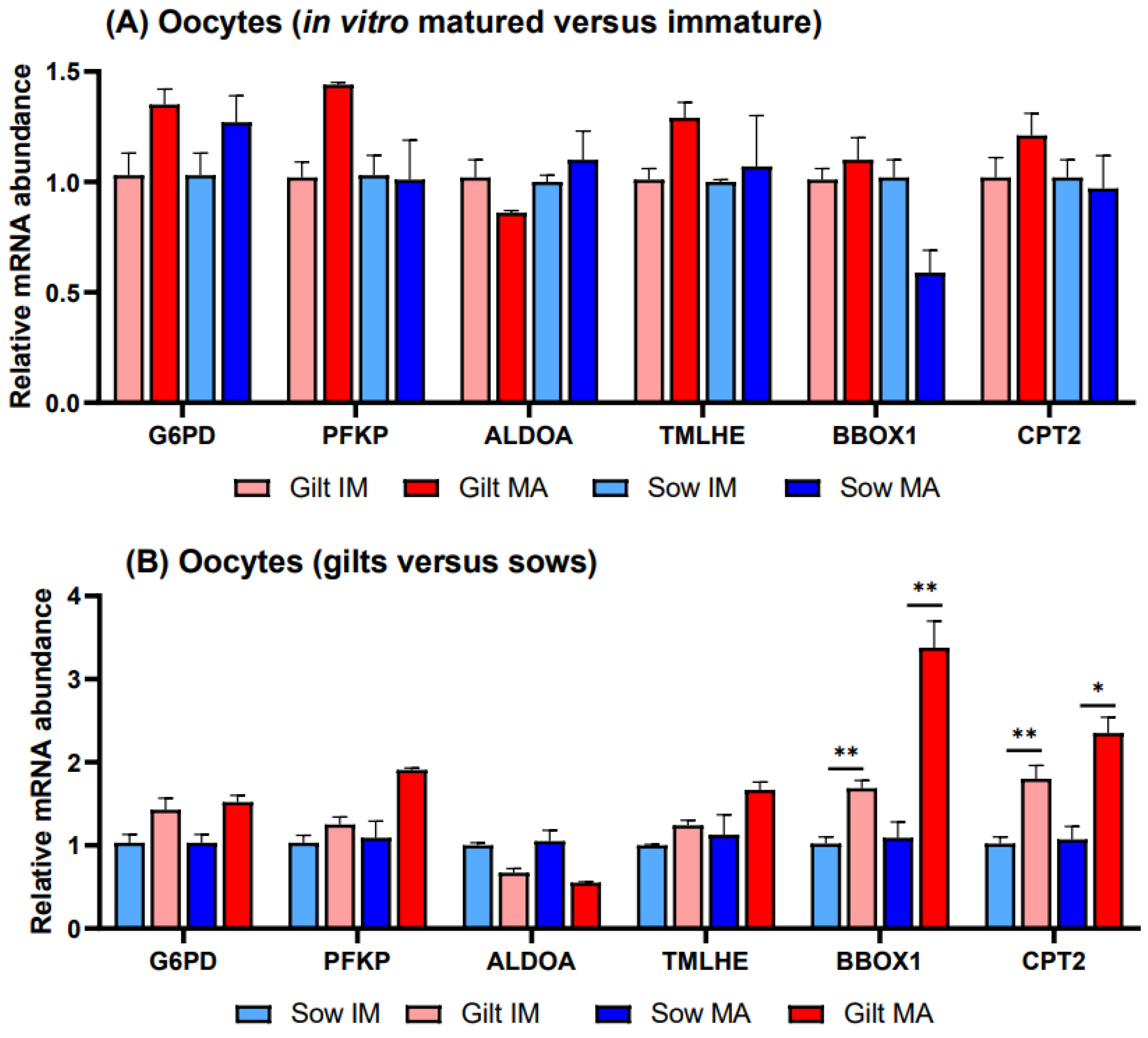
3.2. Gene Expression in Porcine Oocytes
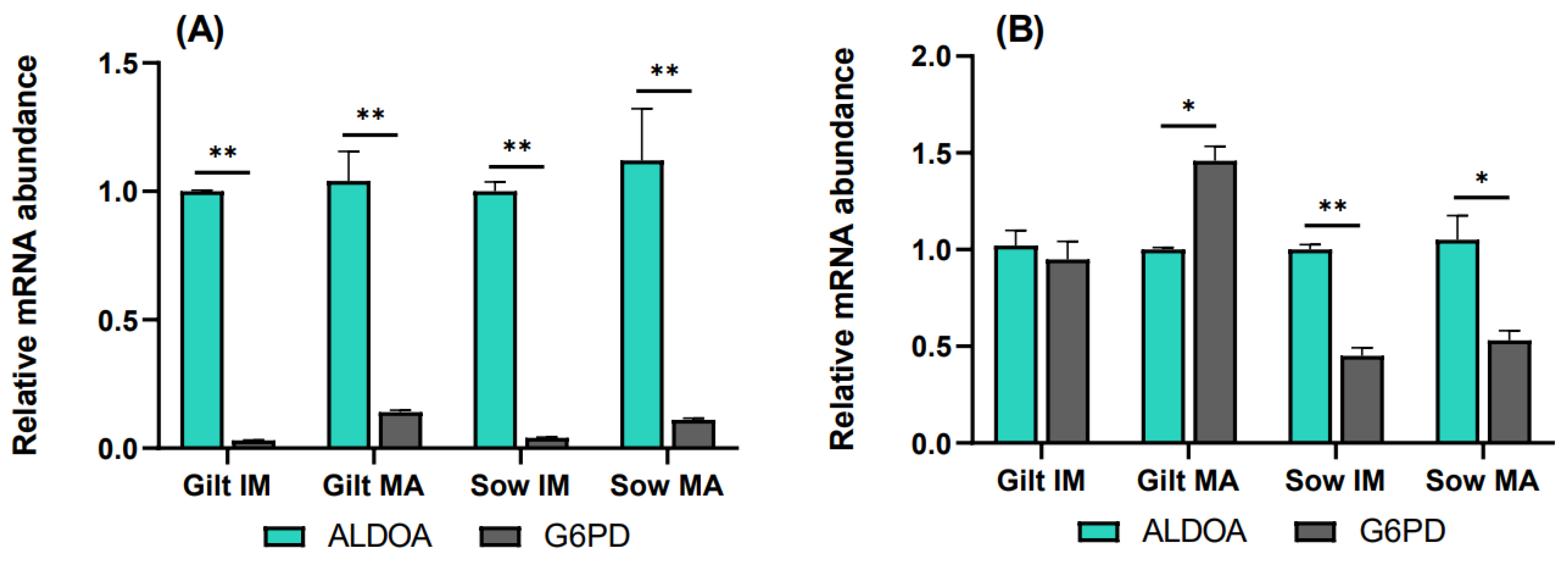
3.3. The Rate of the Pentose Phosphate Pathway Relative to Glycolysis
4. Discussion
4.1. Glucose Metabolism
4.2. Fatty Acid Oxidation
4.3. L-Carnitine Biosynthesis
4.4. Apoptosis
4.5. Gene Expression of PTGS2, GHR and EGFR
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Symbol | TaqMan® Assay | Accession Numbers of Transcript Variants |
|---|---|---|
| G6PD | Ss02690824_g1 | XM_021080744.1 (X1), XM_003360515.5 (X2) |
| ALDOA | Ss06920688_m1 | XM_021087995.1 (X1), XM_021087996.1 (X2), XM_021087997.1 (X3), XM_021087998.1 (X4), XM_021088000.1 (X6), XM_021088001.1 (X7), XM_021088002.1 (X8), XM_021088004.1 (X9) |
| TMLHE | Ss06886117_m1 | XM_003135511.4 (X1), XM_021080447.1 (X2), XM_021080448.1 (X3), XM_021080450.1 (X4), XM_005674022.3 (X5) |
| BBOX1 | Ss06906097_m1 | XM_021083234.1 (X1), XM_021083235.1 (X2), XM_021083236.1 (X3) |
| GHR | Ss03383662_u1 | XM_021076567.1 (X1), XM_013990636.2 (X2), XM_021076568.1 (X3), XM_021076569.1 (X4), XM_021076570.1 (X5), XM_021076571.1 (X6), XM_021076572.1 (X7), XM_021076573.1 (X8), XM_021076574.1 (X9), XM_013990637.2 (X10), XM_021076575.1 (X11) |
| BAX | Ss03375842_u1 | XM_003127290.5 (X1), XM_013998624.2 (X2) |
| BCL2L1 | Ss03383783_s1 | XM_021077292.1 (X1), XM_021077293.1 (X2), XM_021077294.1 (X3), XM_021077295.1 (X4), XM_021077296.1 (X5), XM_021077297.1 (X6), XM_021077298.1 (X7) |
| ACTB | Ss03376563_uH | XM_003124280.5 (X1), XM_021086047.1 (X2) |
| GAPDH | Ss03375629_u1 | XM_021091114.1 (X2) |
| HMBS | Ss03388782_g1 | XM_021102144.1 (X1), XM_021102146.1 (X2), XM_021102147.1 (X3), XM_021102148.1 (X4) |
| HPRT1 | Ss03388274_m1 | XM_021079503.1 (X1), XM_021079504.1 (X2) |
References
- Hunter, M.G. Oocyte Maturation and Ovum Quality in Pigs. Rev. Reprod. 2000, 5, 122–130. [Google Scholar] [CrossRef]
- Krisher, R.L.; Brad, A.M.; Herrick, J.R.; Sparman, M.L.; Swain, J.E. A Comparative Analysis of Metabolism and Viability in Porcine Oocytes during in Vitro Maturation. Anim. Reprod. Sci. 2007, 98, 72–96. [Google Scholar] [CrossRef]
- Hyttel, P.; Fair, T.; Callesen, H.; Greve, T. Oocyte Growth, Capacitation and Final Maturation in Cattle. Theriogenology 1997, 47, 23–32. [Google Scholar] [CrossRef]
- Brevini Gandolfi, T.A.L.; Gandolfi, F. The Maternal Legacy to the Embryo: Cytoplasmic Components and Their Effects on Early Development. Theriogenology 2001, 55, 1255–1276. [Google Scholar] [CrossRef]
- Ledda, S.; Bebbere, D.; Ariu, F.; Pescatori, M.; Pau, S.; Teresa Zedda, M.; Bogliolo, L. Unveiling MRNA Changes During Meiotic Progression and Pre-Implantation Development: Help from Large Animal Models. Curr. Pharm. Des. 2012, 18, 256–263. [Google Scholar] [CrossRef]
- Masala, L.; Burrai, G.P.; Bellu, E.; Ariu, F.; Bogliolo, L.; Ledda, S.; Bebbere, D. Methylation Dynamics during Folliculogenesis and Early Embryo Development in Sheep. Reproduction 2017, 153, 605–619. [Google Scholar] [CrossRef] [PubMed]
- Krisher, R.L. The Effect of Oocyte Quality on Development. J. Anim. Sci. 2004, 82, E14–E23. [Google Scholar] [PubMed]
- Cui, M.S.; Fan, Y.P.; Wu, Y.; Hao, Z.D.; Liu, S.; Chen, X.J.; Zeng, S.M. Porcine Cumulus Cell Influences Ooplasmic Mitochondria-Lipid Distributions, GSH-ATP Contents and Calcium Release Pattern after Electro-Activation. Theriogenology 2009, 71, 412–421. [Google Scholar] [CrossRef]
- Sutton, M.L.; Gilchrist, R.B.; Thompson, J.G. Effects of In-Vivo and in-Vitro Environments on the Metabolism of the Cumulus-Oocyte Complex and Its Influence on Oocyte Developmental Capacity. Hum. Reprod. Update 2003, 9, 35–48. [Google Scholar] [CrossRef] [PubMed]
- Zhou, C.J.; Wu, S.N.; Shen, J.P.; Wang, D.H.; Kong, X.W.; Lu, A.; Li, Y.J.; Zhou, H.X.; Zhao, Y.F.; Liang, C.G. The Beneficial Effects of Cumulus Cells and Oocyte-Cumulus Cell Gap Junctions Depends on Oocyte Maturation and Fertilization Methods in Mice. PeerJ 2016, 4, e1761. [Google Scholar] [CrossRef] [PubMed]
- Auclair, S.; Uzbekov, R.; Elis, S.; Sanchez, L.; Kireev, I.; Lardic, L.; Dalbies-Tran, R.; Uzbekova, S. Absence of Cumulus Cells during in Vitro Maturation Affects Lipid Metabolism in Bovine Oocytes. Am. J. Physiol. Endocrinol. Metab. 2013, 304, E599–E613. [Google Scholar] [CrossRef]
- Wen, J.; Wang, G.L.; Yuan, H.J.; Zhang, J.; Xie, H.L.; Gong, S.; Han, X.; Tan, J.H. Effects of Glucose Metabolism Pathways on Nuclear and Cytoplasmic Maturation of Pig Oocytes. Sci. Rep. 2020, 10, 2782. [Google Scholar] [CrossRef]
- Regassa, A.; Rings, F.; Hoelker, M.; Cinar, U.; Tholen, E.; Looft, C.; Schellander, K.; Tesfaye, D. Transcriptome Dynamics and Molecular Cross-Talk between Bovine Oocyte and Its Companion Cumulus Cells. BMC Genom. 2011, 12, 57. [Google Scholar] [CrossRef] [PubMed]
- Bradley, J.; Swann, K. Mitochondria and Lipid Metabolism in Mammalian Oocytes and Early Embryos. Int. J. Dev. Biol. 2019, 63, 93–103. [Google Scholar] [CrossRef] [PubMed]
- Chen, P.; Redel, B.; Kerns, K.C.; Spate, L.; Prather, R. Challenges and Considerations during In Vitro Production of Porcine Embryos. Cells 2021, 10, 2770. [Google Scholar] [CrossRef] [PubMed]
- Sturmey, R.G.; Leese, H.J. Energy Metabolism in Pig Oocytes and Early Embryos. Reproduction 2003, 126, 197–204. [Google Scholar] [CrossRef] [PubMed]
- Krisher, R.L.; Prather, R.S.; Library, W.O. A Role for the Warburg Effect in Preimplantation Embryo Development: Metabolic Modification to Support Rapid Cell Proliferation. Mol. Reprod. Dev. 2012, 79, 311–320. [Google Scholar] [CrossRef] [PubMed]
- Paczkowski, M.; Silva, E.; Schoolcraft, W.B.; Krisher, R.L. Comparative Importance of Fatty Acid Beta-Oxidation to Nuclear Maturation, Gene Expression, and Glucose Metabolism in Mouse, Bovine, and Porcine Cumulus Oocyte Complexes. Biol. Reprod. 2013, 88, 111–112. [Google Scholar] [CrossRef] [PubMed]
- Prates, E.G.; Nunes, J.T.; Pereira, R.M. A Role of Lipid Metabolism during Cumulus-Oocyte Complex Maturation: Impact of Lipid Modulators to Improve Embryo Production. Mediat. Inflamm. 2014, 2014, 692067. [Google Scholar] [CrossRef]
- Sutton-McDowall, M.L.; Feil, D.; Robker, R.L.; Thompson, J.G.; Dunning, K.R. Utilization of Endogenous Fatty Acid Stores for Energy Production in Bovine Preimplantation Embryos. Theriogenology 2012, 77, 1632–1641. [Google Scholar] [CrossRef]
- Takahashi, T.; Inaba, Y.; Somfai, T.; Kaneda, M.; Geshi, M.; Nagai, T.; Manabe, N. Supplementation of Culture Medium with L-Carnitine Improves Development and Cryotolerance of Bovine Embryos Produced in Vitro. Reprod. Fertil. Dev. 2013, 25, 589–599. [Google Scholar] [CrossRef]
- Wu, G.Q.; Jia, B.Y.; Li, J.J.; Fu, X.W.; Zhou, G.B.; Hou, Y.P.; Zhu, S.E. L-Carnitine Enhances Oocyte Maturation and Development of Parthenogenetic Embryos in Pigs. Theriogenology 2011, 76, 785–793. [Google Scholar] [CrossRef]
- Montjean, D.; Entezami, F.; Lichtblau, I.; Belloc, S.; Gurgan, T.; Menezo, Y. Carnitine Content in the Follicular Fluid and Expression of the Enzymes Involved in Beta Oxidation in Oocytes and Cumulus Cells. J. Assist. Reprod. Genet. 2012, 29, 1221–1225. [Google Scholar] [CrossRef]
- Saint-Dizier, M.; Schoen, J.; Chen, S.; Banliat, C.; Mermillod, P. Composing the Early Embryonic Microenvironment: Physiology and Regulation of Oviductal Secretions. Int. J. Mol. Sci. 2020, 21, 10223. [Google Scholar] [CrossRef]
- Ringseis, R.; Keller, J.; Eder, K. Basic Mechanisms of the Regulation of L-Carnitine Status in Monogastrics and Efficacy of L-Carnitine as a Feed Additive in Pigs and Poultry. J. Anim. Physiol. Anim. Nutr. 2018, 102, 1686–1719. [Google Scholar] [CrossRef]
- Virmani, M.A.; Cirulli, M. The Role of L-Carnitine in Mitochondria, Prevention of Metabolic Inflexibility and Disease Initiation. Int. J. Mol. Sci. 2022, 23, 2717. [Google Scholar] [CrossRef] [PubMed]
- Braga, T.F.; Silva, T.C.F.; Marques, M.G.; de Souza, A.P.; Albring, D.; Silva, L.P.; Caetano, A.R.; Dode, M.A.N.; Franco, M.M. The Dynamics of Gene Expression, Lipid Composition and DNA Methylation Reprogramming Are Different during in Vitro Maturation of Pig Oocytes Obtained from Prepubertal Gilts and Cycling Sows. Reprod. Domest. Anim. 2019, 54, 1217–1229. [Google Scholar] [CrossRef] [PubMed]
- Pawlak, P.; Renska, N.; Pers-Kamczyc, E.; Warzych, E.; Lechniak, D. The Quality of Porcine Oocytes Is Affected by Sexual Maturity of the Donor Gilt. Reprod. Biol. 2011, 11, 1–18. [Google Scholar] [CrossRef]
- Yuan, Y.Q.; Van Soom, A.; Leroy, J.L.M.R.; Dewulf, J.; Van Zeveren, A.; De Kruif, A.; Peelman, L.J. Apoptosis in Cumulus Cells, but Not in Oocytes, May Influence Bovine Embryonic Developmental Competence. Theriogenology 2005, 63, 2147–2163. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.S.; Joo, B.S.; Na, Y.J.; Yoon, M.S.; Choi, O.H.; Kim, W.W. Cumulus Cells Apoptosis as an Indicator to Predict the Quality of Oocytes and the Outcome of IVF-ET. J. Assist. Reprod. Genet. 2001, 18, 490–498. [Google Scholar] [CrossRef]
- Hussein, T.S.; Froiland, D.A.; Amato, F.; Thompson, J.G.; Gilchrist, R.B. Oocytes Prevent Cumulus Cell Apoptosis by Maintaining a Morphogenic Paracrine Gradient of Bone Morphogenetic Proteins. J. Cell Sci. 2005, 118, 5257–5268. [Google Scholar] [CrossRef] [PubMed]
- Assidi, M.; Dufort, I.; Ali, A.; Hamel, M.; Algriany, O.; Dielemann, S.; Sirard, M.A. Identification of Potential Markers of Oocyte Competence Expressed in Bovine Cumulus Cells Matured with Follicle-Stimulating Hormone and/or Phorbol Myristate Acetate in Vitro. Biol. Reprod. 2008, 79, 209–222. [Google Scholar] [CrossRef]
- Caixeta, E.S.; Ripamonte, P.; Franco, M.M.H.; Junior, J.B.; Dode, M.A.N. Effect of Follicle Size on MRNA Expression in Cumulus Cells and Oocytes of Bos Indicus: An Approach to Identify Marker Genes for Developmental Competence. Reprod. Fertil. Dev. 2009, 21, 655–664. [Google Scholar] [CrossRef]
- McKenzie, L.J.; Pangas, S.A.; Carson, S.A.; Kovanci, E.; Cisneros, P.; Buster, J.E.; Amato, P.; Matzuk, M.M. Human Cumulus Granulosa Cell Gene Expression: A Predictor of Fertilization and Embryo Selection in Women Undergoing IVF. Hum. Reprod. 2004, 19, 2869–2874. [Google Scholar] [CrossRef] [PubMed]
- Leoni, G.G.; Palmerini, M.G.; Satta, V.; Succu, S.; Pasciu, V.; Zinellu, A.; Carru, C.; Macchiarelli, G.; Nottola, S.A.; Naitana, S.; et al. Differences in the Kinetic of the First Meiotic Division and in Active Mitochondrial Distribution between Prepubertal and Adult Oocytes Mirror Differences in Their Developmental Competence in a Sheep Model. PLoS ONE 2015, 10, e0124911. [Google Scholar] [CrossRef]
- Leoni, G.G.; Succu, S.; Berlinguer, F.; Rosati, I.; Bebbere, D.; Bogliolo, L.; Ledda, S.; Naitana, S. Delay on the in Vitro Kinetic Development of Prepubertal Ovine Embryos. Anim. Reprod. Sci. 2006, 92, 373–383. [Google Scholar] [CrossRef] [PubMed]
- Sherrer, E.S.; Rathbun, T.J.; Davis, D.L. Fertilization and Blastocyst Development in Oocytes Obtained from Prepubertal and Adult Pigs. J. Anim. Sci. 2004, 82, 102–108. [Google Scholar] [CrossRef]
- Grupen, C.G.; McIlfatrick, S.M.; Ashman, R.J.; Boquest, A.C.; Armstrong, D.T.; Nottle, M.B. Relationship between Donor Animal Age, Follicular Fluid Steroid Content and Oocyte Developmental Competence in the Pig. Reprod. Fertil. Dev. 2003, 15, 81–87. [Google Scholar] [CrossRef]
- Fang, Y.; Zhang, X.; Zhang, J.; Zhong, R.; Zhou, D. Global DNA Methylation and Related MRNA Profiles in Sheep Oocytes and Early Embryos Derived from Pre-Pubertal and Adult Donors. Anim. Reprod. Sci. 2016, 164, 144–151. [Google Scholar] [CrossRef] [PubMed]
- Marchal, R.; Feugang, J.M.; Perreau, C.; Venturi, E.; Terqui, M.; Mermillod, P. Meiotic and Developmental Competence of Prepubertal and Adult Swine Oocytes. Theriogenology 2001, 56, 17–29. [Google Scholar] [CrossRef]
- Romar, R.; De Santis, T.; Papillier, P.; Perreau, C.; Thélie, A.; Dell’Aquila, M.E.; Mermillod, P.; Dalbiès-Tran, R. Expression of Maternal Transcripts during Bovine Oocyte in Vitro Maturation Is Affected by Donor Age. Reprod. Domest. Anim. 2011, 46, e23–e30. [Google Scholar] [CrossRef] [PubMed]
- Leoni, G.G.; Bebbere, D.; Succu, S.; Berlinguer, F.; Mossa, F.; Galioto, M.; Bogliolo, L.; Ledda, S.; Naitana, S. Relations between Relative MRNA Abundance and Developmental Competence of Ovine Oocytes. Mol. Reprod. Dev. 2007, 74, 249–257. [Google Scholar] [CrossRef] [PubMed]
- Silva, T.C.F.; Dode, M.A.N.; Braga, T.F.; Marques, M.G.; Vargas, L.N.; de Faria, O.A.C.; de Souza, A.P.; Albring, D.; Caetano, A.R.; Franco, M.M. Cumulus-Oocyte Complexes from Sows Show Differences in Lipid Metabolism Compared to Cumulus-Oocyte Complexes from Prepubertal Gilts during in Vitro Maturation. Mol. Reprod. Dev. 2023, 90, 323–335. [Google Scholar] [CrossRef]
- Yoshioka, K.; Suzuki, C.; Onishi, A. Defined System for In Vitro Production of Porcine Embryos Using a Single Basic Medium. J. Reprod. Dev. 2008, 54, 208–213. [Google Scholar] [CrossRef]
- Carrasco, L.C.; Romar, R.; Avilés, M.; Gadea, J.; Coy, P. Determination of Glycosidase Activity in Porcine Oviductal Fluid at the Different Phases of the Estrous Cycle. Reproduction 2008, 136, 833–842. [Google Scholar] [CrossRef]
- Bu, G.; Zhu, W.; Liu, X.; Zhang, J.; Yu, L.; Zhou, K.; Wang, S.; Li, Z.; Fan, Z.; Wang, T.; et al. Coordination of Zygotic Genome Activation Entry and Exit by H3K4me3 and H3K27me3 in Porcine Early Embryos. Genome Res. 2022, 32, 1487–1501. [Google Scholar] [CrossRef] [PubMed]
- Haug, L.M.; Jochems, R.; Gaustad, A.H.; Kommisrud, E.; Myromslien, F.D.; Grindflek, E.; Alm-Kristiansen, A.H. Liquid Storage of Porcine in Vitro-Produced Blastocysts; a Practical Approach for Short Storage. Zygote 2023, 31, 441–450. [Google Scholar] [CrossRef]
- Liu, H.; Gao, Y.; Zhai, B.; Jiang, H.; Ding, Y.; Zhang, L.; Li, C.; Deng, Q.; Yu, X.; Zhang, J. The Effects of Polyadenylation Status on MPFs During In Vitro Porcine Oocyte Maturation. Cell. Physiol. Biochem. 2016, 39, 1735–1745. [Google Scholar] [CrossRef]
- Wang, Y.K.; Li, X.; Song, Z.Q.; Yang, C.X. Methods of RNA Preparation Affect MRNA Abundance Quantification of Reference Genes in Pig Maturing Oocytes. Reprod. Domest. Anim. 2017, 52, 722–730. [Google Scholar] [CrossRef]
- Zhu, G.Y.; Feng, S.T.; Li, J.T.; Mu, Y.L.; Pan, D.K.; Guo, B.R. Comparison of Gene Expression Patterns between Porcine Cumulus-Oocyte Complexes and Naked Oocytes. S. Afr. J. Anim. Sci. 2007, 37, 57–63. [Google Scholar] [CrossRef]
- Shi, L.; Zhai, Y.; Zhao, Y.; Kong, X.; Zhang, D.; Yu, H.; Li, Z. ELF4 Is Critical to Zygotic Gene Activation and Epigenetic Reprogramming during Early Embryonic Development in Pigs. Front. Vet. Sci. 2022, 9, 954601. [Google Scholar] [CrossRef] [PubMed]
- Yuan, Y.; Ida, J.M.; Paczkowski, M.; Krisher, R.L. Identification of Developmental Competence-Related Genes in Mature Porcine Oocytes. Mol. Reprod. Dev. 2011, 78, 565–575. [Google Scholar] [CrossRef] [PubMed]
- Brązert, M.; Kranc, W.; Nawrocki, M.J.; Sujka–Kordowska, P.; Konwerska, A.; Jankowski, M.; Kocherova, I.; Celichowski, P.; Jeseta, M.; Ożegowska, K.; et al. New Markers for Regulation of Transcription and Macromolecule Metabolic Process in Porcine Oocytes during in Vitro Maturation. Mol. Med. Rep. 2020, 21, 1537. [Google Scholar] [CrossRef]
- Budna, J.; Bryja, A.; Celichowski, P.; Kahan, R.; Kranc, W.; Ciesiółka, S.; Rybska, M.; Borys, S.; Jeseta, M.; Bukowska, D.; et al. Genes of Cellular Components of Morphogenesis in Porcine Oocytes before and after IVM. Reproduction 2017, 154, 535–545. [Google Scholar] [CrossRef]
- Kulus, M.; Kranc, W.; Sujka-Kordowska, P.; Celichowski, P.; Konwerska, A.; Jankowski, M.; Jeseta, M.; Skowroński, M.T.; Piotrowska-Kempisty, H.; Bukowska, D.; et al. Transcriptomic Analysis of Expression of Genes Regulating Cell Cycle Progression in Porcine Ovarian Granulosa Cells during Short-Term in Vitro Primary Culture. Histochem. Cell Biol. 2020, 153, 397–412. [Google Scholar] [CrossRef]
- Caetano, L.C.; Miranda-Furtado, C.L.; Batista, L.A.; Pitangui-Molina, C.P.; Higa, T.T.; Padovan, C.C.; Rosa-e-Silva, A.C.J.d.S. Validation of Reference Genes for Gene Expression Studies in Bovine Oocytes and Cumulus Cells Derived from in Vitro Maturation. Anim. Reprod. 2019, 16, 290–296. [Google Scholar] [CrossRef]
- Andersen, C.L.; Jensen, J.L.; Ørntoft, T.F. Normalization of Real-Time Quantitative Reverse Transcription-PCR Data: A Model-Based Variance Estimation Approach to Identify Genes Suited for Normalization, Applied to Bladder and Colon Cancer Data Sets. Cancer Res. 2004, 64, 5245–5250. [Google Scholar] [CrossRef]
- Pfaffl, M.W. A New Mathematical Model for Relative Quantification in Real-Time RT-PCR. Nucleic Acids Res. 2001, 29, E45. [Google Scholar] [CrossRef]
- Ramakers, C.; Ruijter, J.M.; Lekanne Deprez, R.H.; Moorman, A.F.M. Assumption-Free Analysis of Quantitative Real-Time Polymerase Chain Reaction (PCR) Data. Neurosci. Lett. 2003, 339, 62–66. [Google Scholar] [CrossRef] [PubMed]
- Untergasser, A.; Ruijter, J.M.; Benes, V.; van den Hoff, M.J.B. Web-Based LinRegPCR: Application for the Visualization and Analysis of (RT)-QPCR Amplification and Melting Data. BMC Bioinform. 2021, 22, 398. [Google Scholar] [CrossRef]
- Paczkowski, M.; Krisher, R. Aberrant Protein Expression Is Associated with Decreased Developmental Potential in Porcine Cumulus-Oocyte Complexes. Mol. Reprod. Dev. 2010, 77, 51–58. [Google Scholar] [CrossRef]
- Cetica, P.; Pintos, L.; Dalvit, G.; Beconi, M. Activity of Key Enzymes Involved in Glucose and Triglyceride Catabolism during Bovine Oocyte Maturation in Vitro. Reproduction 2002, 124, 675–681. [Google Scholar] [CrossRef]
- Brad, A.M.; Bormann, C.L.; Swain, J.E.; Durkin, R.E.; Johnson, A.E.; Clifford, A.L.; Krisher, R.L. Glutathione and Adenosine Triphosphate Content of in Vivo and in Vitro Matured Porcine Oocytes. Mol. Reprod. Dev. 2003, 64, 492–498. [Google Scholar] [CrossRef] [PubMed]
- Sutton-McDowall, M.L.; Gilchrist, R.B.; Thompson, J.G. The Pivotal Role of Glucose Metabolism in Determining Oocyte Developmental Competence. Reproduction 2010, 139, 685–695. [Google Scholar] [CrossRef] [PubMed]
- Lonergan, P.; Gutiérrez-Adán, A.; Rizos, D.; Pintado, B.; de La Fuente, J.; Boland, M.P. Relative Messenger RNA Abundance in Bovine Oocytes Collected in Vitro or in Vivo before and 20 Hr after the Preovulatory Luteinizing Hormone Surge. Mol. Reprod. Dev. 2003, 66, 297–305. [Google Scholar] [CrossRef]
- Lee, Y.; Kim, J.; Kim, H.; Han, J.E.; Kim, S.; Kang, K.H.; Kim, D.; Kim, J.M.; Koh, H. Pyruvate Dehydrogenase Kinase Protects Dopaminergic Neurons from Oxidative Stress in Drosophila DJ-1 Null Mutants. Mol. Cells 2022, 45, 454–464. [Google Scholar] [CrossRef]
- Dumollard, R.; Ward, Z.; Carroll, J.; Duchen, M.R. Regulation of Redox Metabolism in the Mouse Oocyte and Embryo. Development 2007, 134, 455–465. [Google Scholar] [CrossRef] [PubMed]
- Lowe, J.L.; Bartolac, L.K.; Bathgate, R.; Grupen, C.G. Supplementation of Culture Medium with L-Carnitine Improves the Development and Cryotolerance of in Vitro-Produced Porcine Embryos. Reprod. Fertil. Dev. 2017, 29, 2357–2366. [Google Scholar] [CrossRef]
- Pawlak, P.; Chabowska, A.; Malyszka, N.; Lechniak, D. Mitochondria and Mitochondrial DNA in Porcine Oocytes and Cumulus Cells—A Search for Developmental Competence Marker. Mitochondrion 2016, 27, 48–55. [Google Scholar] [CrossRef] [PubMed]
- Torner, H.; Ghanem, N.; Ambros, C.; Hölker, M.; Tomek, W.; Phatsara, C.; Alm, H.; Sirard, M.A.; Kanitz, W.; Schellander, K.; et al. Molecular and Subcellular Characterisation of Oocytes Screened for Their Developmental Competence Based on Glucose-6-Phosphate Dehydrogenase Activity. Reproduction 2008, 135, 197–212. [Google Scholar] [CrossRef]
- Pawlak, P.; Malyszka, N.; Szczerbal, I.; Kolodziejski, P. Fatty Acid Induced Lipolysis Influences Embryo Development, Gene Expression and Lipid Droplet Formation in the Porcine Cumulus Cells. Biol. Reprod. 2020, 103, 36–48. [Google Scholar] [CrossRef] [PubMed]
- Dvoran, M.; Nemcova, L.; Kalous, J. An Interplay between Epigenetics and Translation in Oocyte Maturation and Embryo Development: Assisted Reproduction Perspective. Biomedicines 2022, 10, 1689. [Google Scholar] [CrossRef] [PubMed]
- Picton, H.; Briggs, D.; Gosden, R. The Molecular Basis of Oocyte Growth and Development. Mol. Cell. Endocrinol. 1998, 145, 27–37. [Google Scholar] [CrossRef] [PubMed]
- Thélie, A.; Papillier, P.; Pennetier, S.; Perreau, C.; Traverso, J.M.; Uzbekova, S.; Mermillod, P.; Joly, C.; Humblot, P.; Dalbiès-Tran, R. Differential Regulation of Abundance and Deadenylation of Maternal Transcripts during Bovine Oocyte Maturation In Vitro and In Vivo. BMC Dev. Biol. 2007, 7, 125. [Google Scholar] [CrossRef] [PubMed]
- He, M.; Zhang, T.; Yang, Y.; Wang, C. Mechanisms of Oocyte Maturation and Related Epigenetic Regulation. Front. Cell Dev. Biol. 2021, 9, 654028. [Google Scholar] [CrossRef]
- Ma, P.; Schultz, R.M. HDAC1 and HDAC2 in Mouse Oocytes and Preimplantation Embryos: Specificity versus Compensation. Cell Death Differ. 2016, 23, 1119–1127. [Google Scholar] [CrossRef]
- Mito, T.; Hoshi, H. In Vitro Culture of Late Stage Pig Embryos in a Chemically Defined Medium, Porcine Blastocyst Medium (PBM). In Comparative Embryo Culture. Methods in Molecular Biology; Herrick, J., Ed.; Humana: New York, NY, USA, 2019; pp. 105–113. [Google Scholar]
- Godárová, A.; Litzlbauer, E.; Brunner, S.; Agu, A.C.; Lohninger, A.; Hofbauer, R. L-Carnitine Regulates MRNA Expression Levels of the Carnitine Acyltransferases-CPT1A, CPT2, and CRAT. Monatshefte Chem. 2005, 136, 1349–1363. [Google Scholar] [CrossRef]
- Macaulay, A.D.; Gilbert, I.; Scantland, S.; Fournier, E.; Ashkar, F.; Bastien, A.; Shojaei Saadi, H.A.; Gagné, D.; Sirard, M.A.; Khandjian, É.W.; et al. Cumulus Cell Transcripts Transit to the Bovine Oocyte in Preparation for Maturation. Biol. Reprod. 2016, 94, 16. [Google Scholar] [CrossRef]
- Sugimura, S.; Ritter, L.J.; Rose, R.D.; Thompson, J.G.; Smitz, J.; Mottershead, D.G.; Gilchrist, R.B. Promotion of EGF Receptor Signaling Improves the Quality of Low Developmental Competence Oocytes. Dev. Biol. 2015, 403, 139–149. [Google Scholar] [CrossRef]
- Somfai, T.; Kaneda, M.; Akagi, S.; Watanabe, S.; Haraguchi, S.; Mizutani, E.; Dang-Nguyen, T.Q.; Geshi, M.; Kikuchi, K.; Nagai, T. Enhancement of Lipid Metabolism with L-Carnitine during in Vitro Maturation Improves Nuclear Maturation and Cleavage Ability of Follicular Porcine Oocytes. Reprod. Fertil. Dev. 2011, 23, 912–920. [Google Scholar] [CrossRef]
- Chankitisakul, V.; Somfai, T.; Inaba, Y.; Techakumphu, M.; Nagai, T. Supplementation of Maturation Medium with L-Carnitine Improves Cryo-Tolerance of Bovine in Vitro Matured Oocytes. Theriogenology 2013, 79, 590–598. [Google Scholar] [CrossRef]
- Carrillo-González, D.F.; Rodríguez-Osorio, N.; Long, C.R.; Vásquez-Araque, N.A.; Maldonado-Estrada, J.G. L-Carnitine Supplementation during In Vitro Maturation and In Vitro Culture Does Not Affect the Survival Rates after Vitrification and Warming but Alters Inf-T and Ptgs2 Gene Expression. Int. J. Mol. Sci. 2020, 21, 5601. [Google Scholar] [CrossRef]
- Xu, H.; Jia, C.; Cheng, W.; Zhang, T.; Tao, R.; Ma, Y.; Si, L.; Xu, Y.; Li, J. The Effect of L-Carnitine Additive during In Vitro Maturation on the Vitrification of Pig Oocytes. Cell. Reprogram. 2020, 22, 198–207. [Google Scholar] [CrossRef]
- Ramos-Ibeas, P.; Gimeno, I.; Cañón-Beltrán, K.; Gutiérrez-Adán, A.; Rizos, D.; Gómez, E. Senescence and Apoptosis during In Vitro Embryo Development in a Bovine Model. Front. Cell Dev. Biol. 2020, 8, 619902. [Google Scholar] [CrossRef] [PubMed]
- Zuccotti, M.; Merico, V.; Cecconi, S.; Redi, C.A.; Garagna, S. What Does It Take to Make a Developmentally Competent Mammalian Egg? Hum. Reprod. Update 2011, 17, 525–540. [Google Scholar] [CrossRef] [PubMed]
- Luciano, A.M.; Franciosi, F.; Dieci, C.; Lodde, V. Changes in Large-Scale Chromatin Structure and Function during Oogenesis: A Journey in Company with Follicular Cells. Anim. Reprod. Sci. 2014, 149, 3–10. [Google Scholar] [CrossRef]




| Gene Symbol | Function | Accession Number | TaqMan® Assay |
|---|---|---|---|
| G6PD | Pentose phosphate pathway | XM_003360515.5 †† | Ss02690824_g1 |
| PFKP | Glycolysis | XM_021065066.1 | Ss06887532_m1 |
| ALDOA | Glycolysis | XM_021087995.1 †† | Ss06920688_m1 |
| TMLHE | Carnitine biosynthesis | XM_003135511.4 †† | Ss06886117_m1 |
| BBOX1 | Carnitine biosynthesis | XM_021083234.1 †† | Ss06906097_m1 |
| CPT2 | Fatty acid metabolism | NM_001246243.1 | Ss04322743_m1 |
| GHR † | Growth hormone receptor | NM_214254.2 †† | Ss03383662_u1 |
| EGFR † | Epidermal growth factor receptor | NM_214007.1 | Ss03393423_u1 |
| PTGS2 † | Cumulus cell expansion and function | NM_214321.1 | Ss03394694_m1 |
| BAX † | Pro-apoptotic | XM_003127290.5 †† | Ss03375842_u1 |
| BCL2L1 † | Anti-apoptotic | NM_214285.1 †† | Ss03383783_s1 |
| ACTB | Cytoskeletal structural protein | XM_003124280.5 †† | Ss03376563_uH |
| GAPDH | Glycolysis | NM_001206359.1 †† | Ss03375629_u1 |
| HMBS | Heme biosynthesis | NM_001097412.1 †† | Ss03388782_g1 |
| HPRT1 | Recycling of purines | NM_001032376.2 | Ss03388274_m1 |
| YWHAG | Signal transduction | XM_005661962.3 | Ss06938931_s1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Haug, L.M.; Wilson, R.C.; Gaustad, A.H.; Jochems, R.; Kommisrud, E.; Grindflek, E.; Alm-Kristiansen, A.H. Cumulus Cell and Oocyte Gene Expression in Prepubertal Gilts and Sows Identifies Cumulus Cells as a Prime Informative Parameter of Oocyte Quality. Biology 2023, 12, 1484. https://doi.org/10.3390/biology12121484
Haug LM, Wilson RC, Gaustad AH, Jochems R, Kommisrud E, Grindflek E, Alm-Kristiansen AH. Cumulus Cell and Oocyte Gene Expression in Prepubertal Gilts and Sows Identifies Cumulus Cells as a Prime Informative Parameter of Oocyte Quality. Biology. 2023; 12(12):1484. https://doi.org/10.3390/biology12121484
Chicago/Turabian StyleHaug, Linda Marijke, Robert C. Wilson, Ann Helen Gaustad, Reina Jochems, Elisabeth Kommisrud, Eli Grindflek, and Anne Hege Alm-Kristiansen. 2023. "Cumulus Cell and Oocyte Gene Expression in Prepubertal Gilts and Sows Identifies Cumulus Cells as a Prime Informative Parameter of Oocyte Quality" Biology 12, no. 12: 1484. https://doi.org/10.3390/biology12121484
APA StyleHaug, L. M., Wilson, R. C., Gaustad, A. H., Jochems, R., Kommisrud, E., Grindflek, E., & Alm-Kristiansen, A. H. (2023). Cumulus Cell and Oocyte Gene Expression in Prepubertal Gilts and Sows Identifies Cumulus Cells as a Prime Informative Parameter of Oocyte Quality. Biology, 12(12), 1484. https://doi.org/10.3390/biology12121484