
Intra-Population Alteration on Voltinism of Asian Corn Borer in Response to Climate Warming


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. ACB Populations
2.2. Selection for Univoltine, Multivoltine, and Non-Diapausing Strains
2.3. Photoperiodic and Thermal Response
2.4. Calculation of Effective Accumulated Temperature
2.5. Data Analysis
3. Results
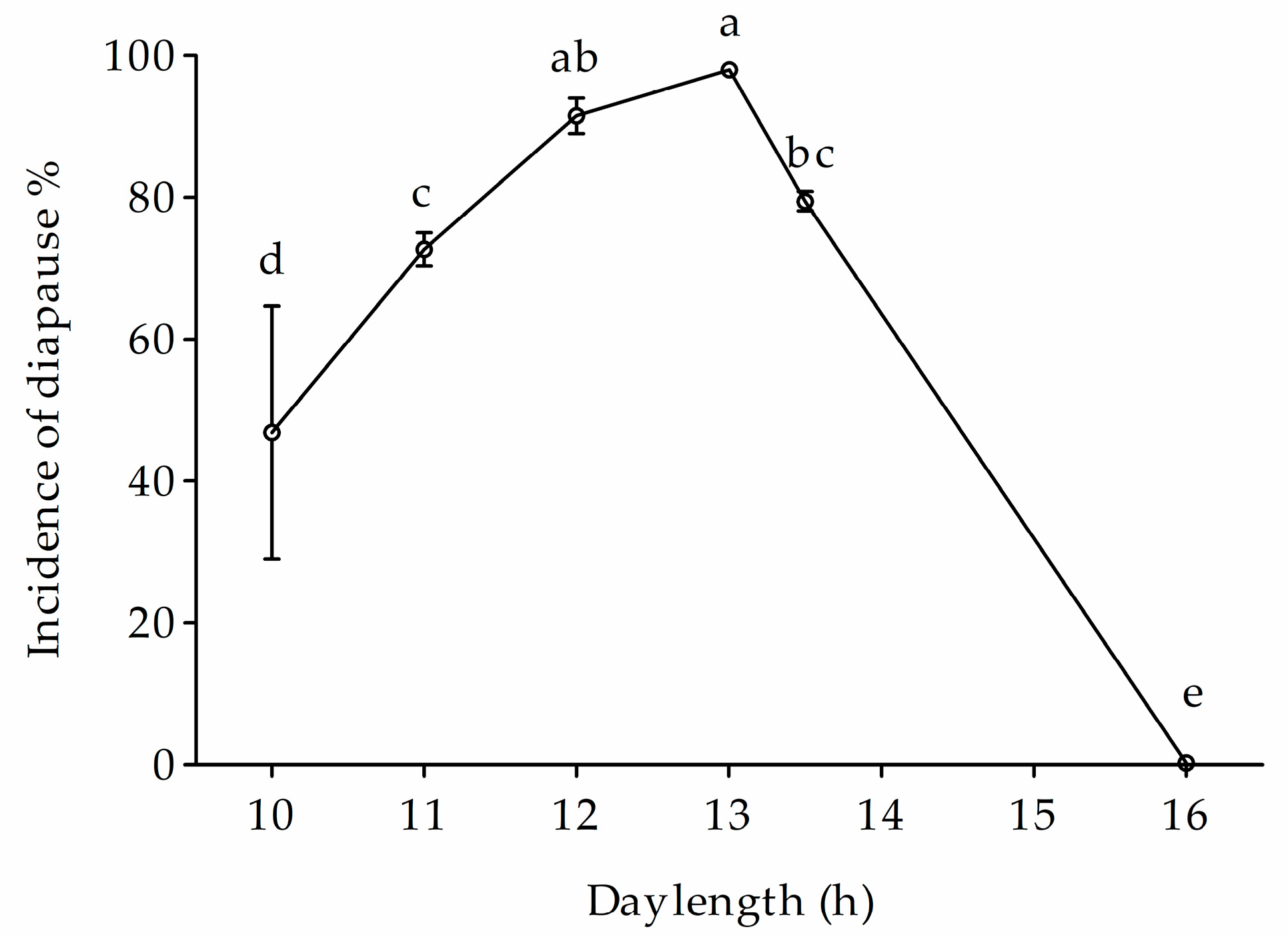
3.1. Variation of Voltinism in HH Population
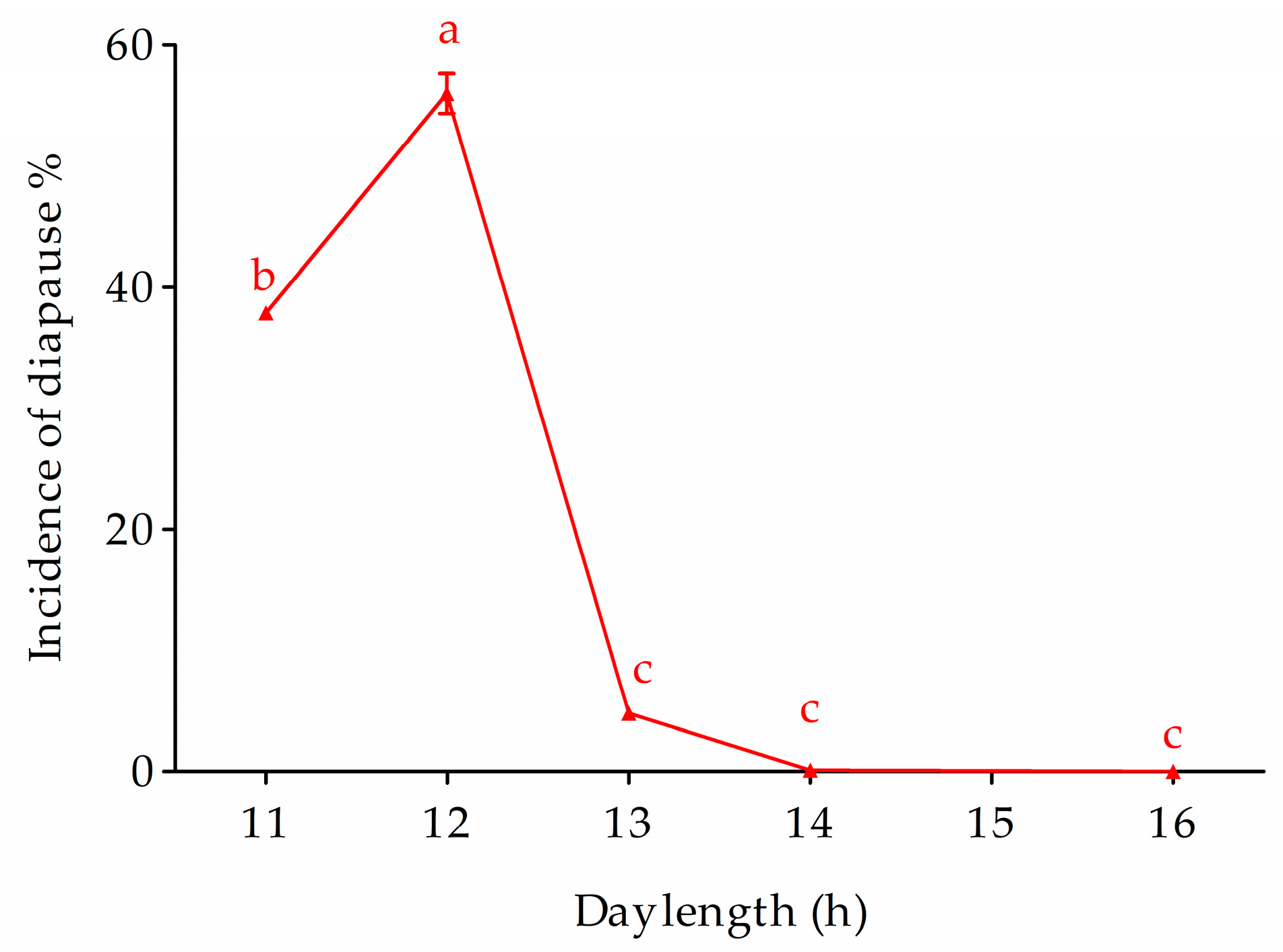
3.2. Development of Univoltine Traits under Selection
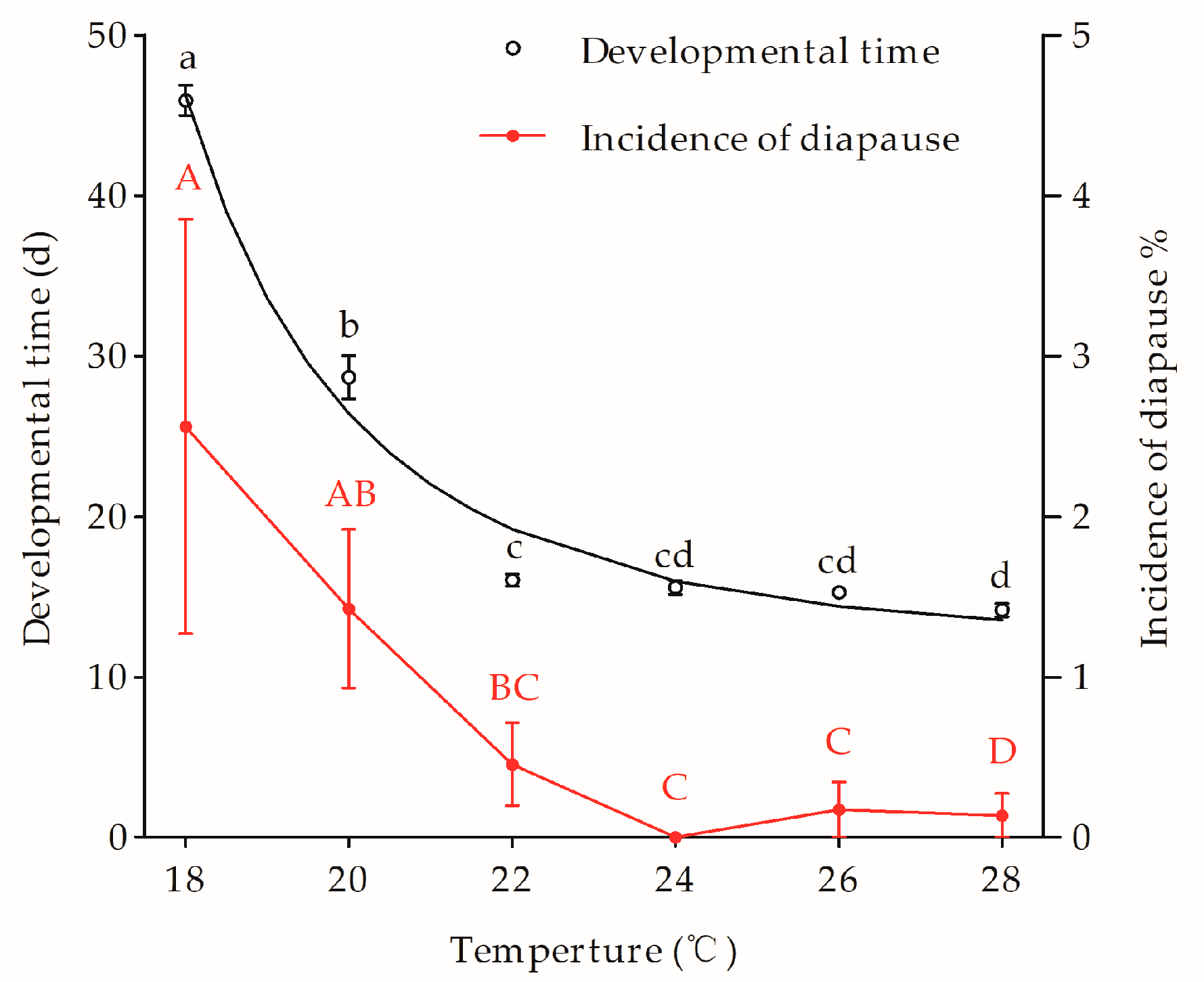
3.3. Multivoltine Strain Lm and Its Response to the Photo and Thermal Induction
- R2 = 0.980
- t(T) is development time (day) and T is the temperature (°C).
3.4. Non-Diapausing Strain Ln and Its Response to the Photo and Thermal Induction
- R2 = 0.993.

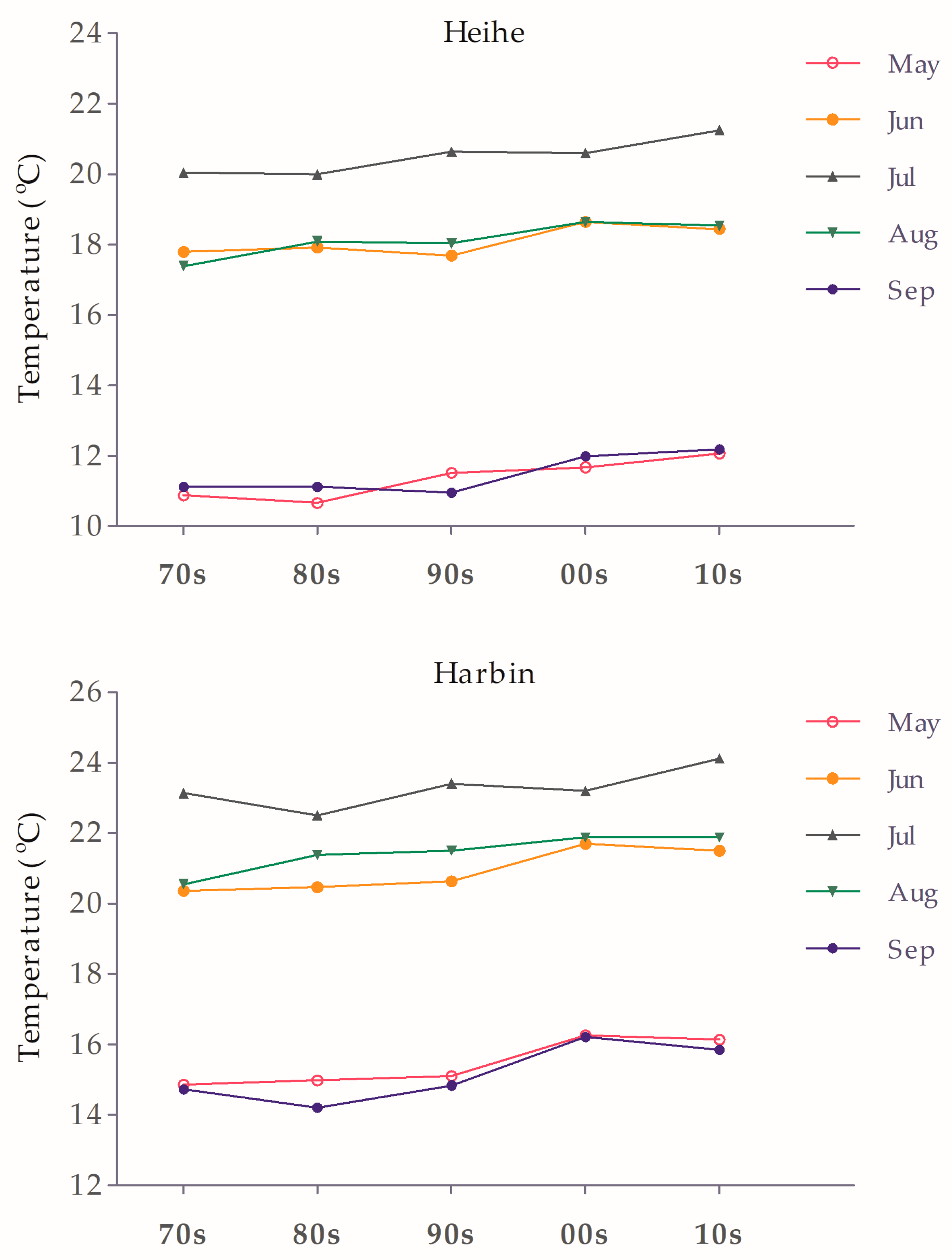
3.5. Voltinism under Climate Warming
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
Appendix A


Appendix B

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Daylength (h) | Temperature (°C) | Selection Procedure |
|---|---|---|
| 16 | 26 | Population HH: 851 pupae from overwintering larvae; placed in 7 oviposition cages (103 + 188 + 168 + 163 + 113 + 80 + 36) according to the pupation time; HH1: 1649 diapause larvae |
| 16 | 27 | HH1: 867 pupae, bulk mating; HH2: obtained 2095 diapause larvae. |
| 16 | 27 | HH2: 662 pupae, bulk mating. |
| 16 | 28 | HH2: 662 pupa, bulk mating; HH3: 943 diapause larvae. |
| 16 | 28 | HH3: sibling mate 4 lines with 668 months emergence, HH4: 201 diapause larvae. |
| 16 | 30 | HH3: sibling mate 15 lines with 668 months emergence, HH4: 159 diapause larvae |
| 16 | 28 | HH4: 234 pupae from pooled diapause larvae (201+159), sibling 7 paired-mating, HH5: 265 diapause larvae. |
| 16 | 28 | HH5: 135 pupae, bulking mating, HH6: 265 diapause larvae. |
| 16 | 28 | HH6: 156 pupae, sibling mate 5 lines HH7: 601 diapause larvae. uni-voltine strain Lu |
References
- Danks, H.V. Insect Dormancy: An Ecological Perspective; Biological Survey of Canada (Terrestrial Arthropods): Ottawa, ON, Canada, 1987. [Google Scholar]
- Koštál, V. Eco-Physiological Phases of Insect Diapause. J. Insect Physiol. 2006, 52, 113–127. [Google Scholar] [CrossRef] [PubMed]
- Shintani, Y. Artificial Selection for Responsiveness to Photoperiodic Change Alters the Response to Stationary Photoperiods in Maternal Induction of Egg Diapause in the Rice Leaf Bug Trigonotylus Caelestialium. J. Insect Physiol. 2009, 55, 818–824. [Google Scholar] [CrossRef]
- Kiran, R.; Bharath, K.N.; Shivkumar, S.; Mir Nisar, A.; Sukhen Roy, C. Variation of Embryonic Diapause Induction in Bivoltine Silkworm Bombyx Mori L (Lepidoptera: Bombycidae) under Controlled Conditions. Int. J. Ind. Entomol. 2021, 43, 37–44. [Google Scholar]
- Shintani, Y.; Hirose, Y.; Terao, M. Effects of Temperature, Photoperiod and Soil Humidity on Induction of Pseudopupal Diapause in the Bean Blister Beetle Epicauta Gorhami. Physiol. Entomol. 2011, 36, 14–20. [Google Scholar] [CrossRef]
- Xu, L.R.; Ni, X.; Wang, Z.Y.; He, K.L. Effects of Photoperiod and Temperature on Diapause Induction in Conogethes Punctiferalis (Lepidoptera: Pyralidae). Insect Sci. 2014, 21, 556–563. [Google Scholar] [CrossRef]
- Tobin, P.C.; Nagarkatti, S.; Saunders, M.C. Diapause Maintenance and Termination in Grape Berry Moth (Lepidoptera: Tortricidae). Environ. Entomol. 2002, 31, 708–713. [Google Scholar] [CrossRef] [Green Version]
- Chen, C.; Xia, Q.W.; Fu, S.; Wu, X.F.; Xue, F.S. Effect of Photoperiod and Temperature on the Intensity of Pupal Diapause in the Cotton Bollworm, Helicoverpa Armigera (Lepidoptera: Noctuidae). B Entomol. Res. 2014, 104, 12–18. [Google Scholar] [CrossRef]
- Dalin, P. Diapause Induction and Termination in a Commonly Univoltine Leaf Beetle (Phratora Vulgatissima). Insect Sci. 2011, 18, 443–450. [Google Scholar] [CrossRef]
- Sokolova, I.V. Univoltine Seasonal Cycle and Obligate Diapause in the Noctuid Moth Charanyca Trigrammica Hufn.(Lepidoptera, Noctuidae). Entomol. Rev. 2007, 87, 793–798. [Google Scholar] [CrossRef]
- Denlinger, D.L. Regulation of Diapause. Annu. Rev. Entomol. 2002, 47, 93. [Google Scholar] [CrossRef]
- Beck, S.D. Insect Photoperiodism; Academic Press Inc.: New York, NY, USA, 1980. [Google Scholar]
- Saunders, D.S.; Lewis, R.D.; Warman, G.R. Photoperiodic Induction of Diapause: Opening the Black Box. Physiol. Entomol. 2004, 29, 1–15. [Google Scholar] [CrossRef]
- Saunders, D.S. Insect Photoperiodism: Effects of Temperature on the Induction of Insect Diapause and Diverse Roles for the Circadian System in the Photoperiodic Response. Entomol. Sci. 2014, 17, 25–40. [Google Scholar] [CrossRef]
- Tauber, M.J.; Tauber, C.A.; Masaki, S. Seasonal Adaptations of Insects; Oxford University Press.: New York, NY, USA, 1986. [Google Scholar]
- Saunders, D.S. Insect Clocks; Elsevier: Amsterdam, The Netherlands, 2002. [Google Scholar]
- Ju, H.; Xiong, W.; Xu, Y.L.; Lin, E.D. Climate Change and Its Impacts in Northeast China. Chin. Agric. Sci. Bull. 2007, 23, 345–349. [Google Scholar]
- Yang, X.; Lin, E.; Ma, S.M.; Ju, H.; Guo, L.P.; Xiong, W.; Li, Y.; Xu, Y.L. Adaptation of Agriculture to Warming in Northeast China. Clim. Change 2007, 84, 45–58. [Google Scholar] [CrossRef]
- Tao, F.L.; Yokozawa, M.; Xu, Y.L.; Hayashi, Y.; Zhang, Z. Climate Changes and Trends in Phenology and Yields of Field Crops in China, 1981–2000. Agric. For. Meteorol. 2006, 138, 82–92. [Google Scholar] [CrossRef]
- Liu, Z.; Li, S.M.; Mi, C.Q.; Wang, H.; Ma, Q.; Li, L.; Zhang, X.D.; Zhu, D.H. A Spatial-Temporal Diffusion Study of Maize Variety in China Using a New Two-Dimensional Representation. J. Food Agric. Environ. 2012, 10, 331–337. [Google Scholar]
- Forister, M.L.; Shapiro, A.M. Climatic Trends and Advancing Spring Flight of Butterflies in Lowland California. Glob. Change Biol. 2003, 9, 1130–1135. [Google Scholar] [CrossRef]
- Tobin, P.C.; Nagarkatti, S.; Loeb, G.; Saunders, M.C. Historical and Projected Interactions between Climate Change and Insect Voltinism in a Multivoltine Species. Glob. Change Biol. 2008, 14, 951–957. [Google Scholar] [CrossRef]
- Jönsson, A.M.; Appelberg, G.; Harding, S.; Bärring, L. Spatio-Temporal Impact of Climate Change on the Activity and Voltinism of the Spruce Bark Beetle, Ips Typographus. Glob. Change Biol. 2009, 15, 486–499. [Google Scholar] [CrossRef]
- Altermatt, F. Climatic Warming Increases Voltinism in European Butterflies and Moths. Proc. R. Soc. B Biol. Sci. 2010, 277, 1281–1287. [Google Scholar] [CrossRef] [Green Version]
- Stoeckli, S.; Hirschi, M.; Spirig, C.; Calanca, P.; Rotach, M.W.; Samietz, J. Impact of Climate Change on Voltinism and Prospective Diapause Induction of a Global Pest Insect–Cydia Pomonella (L.). PLoS ONE 2012, 7, e35723. [Google Scholar] [CrossRef] [PubMed]
- Zhou, D.R.; He, K.L.; Wang, Z.Y.; Ye, Z.H.; Wen, L.P.; Gao, Y.X.; Song, Y.Y. Asian Corn Borer and Its Integrated Management; Golden Shield Press: Beijing, China, 1995. [Google Scholar]
- Gui, C.M.; Li, L.Q.; Hun, W.X.; Liu, J.R. A Review on Subspecies of the Asian Corn Borer in Jilin Province. Plant. Dis. Insect Pests 1991, 28–31. [Google Scholar]
- Li, J.P.; Wang, Y.S.; Xie, W.M.; Yang, G.H. A Preliminary Study on the Ecotype of Ostrinia furnacalis in Northern China. J. Maize Sci. 1992, 00, 69–72. [Google Scholar]
- Gong, H.F.; Chen, P.; Wang, R.; Lian, N.L.; Xia, Z.H.; Yi, Y. The Influence of Photoperiod and Temperature on the Diapause of the Asian Corn Borer Ostrinia furnacallis (Guenee). Acta Entomol. Sin. 1984, 27, 280–286. [Google Scholar] [CrossRef]
- Lu, X.; Zhou, D.R. On the Photoperiodic Reactions in Different Voltinism Ecotypes of Asian Corn Borer. J. Plant Prot. 2000, 27, 12–16. [Google Scholar] [CrossRef]
- Guo, J.Q.; Zhang, H.G.; Wang, Z.Y.; He, K.L. Effects of Photoperiod and Temperature on Diapause Induction in Ostrinia furnacalis. Acta Entomol. Sin. 2013, 56, 996–1003. [Google Scholar] [CrossRef]
- Huang, L.L.; Chen, C.; Xiao, L.; Xia, Q.W.; Hu, L.T.; Xue, F.S. Geographical Variation and Inheritance of the Photoperiodic Response Controlling Larval Diapause in Two Distinct Voltine Ecotypes of the Asian Cornborer, Ostrinia Furnacalis. Physiol. Entomol. 2013, 38, 126–132. [Google Scholar] [CrossRef]
- Huang, L.L.; Tang, J.J.; Chen, C.; He, H.M.; Gao, Y.L.; Xue, F.S. Diapause Incidence and Critical Day Length of Asian Corn Borer (Ostrinia furnacalis) Populations Exhibit a Latitudinal Cline in Both Pure and Hybrid Strains. J. Pest. Sci. 2020, 93, 559–568. [Google Scholar] [CrossRef]
- Xia, Q.W.; Chen, C.; Tu, X.Y.; Yang, H.Z.; Xue, F.S. Inheritance of Photoperiodic Induction of Larval Diapause in the Asian Corn Borer Ostrinia furnacalis. Physiol. Entomol. 2012, 37, 185–191. [Google Scholar] [CrossRef]
- Xiao, L.; He, H.M.; Zhong, P.S.; Fu, S.; Chen, C.; Xue, F.S. Inheritance of Photoperiodic Control of Larval Diapause in the Asian Corn Borer Ostrinia furnacalis (Guenée). B Entomol. Res. 2015, 105, 326–334. [Google Scholar] [CrossRef]
- Fu, S.; Chen, C.; Xiao, L.; He, H.M.; Xue, F.S. Inheritance of Diapause in Crosses between the Northernmost and the Southernmost Strains of the Asian Corn Borer Ostrinia furnacalis. PLoS ONE 2015, 10, e0118186. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.G.; Lu, X.; He, K.L.; Wang, Z.Y. Cold Hardiness and the Strategies of Ostrinia furnacalis Survival at Supercooled Temperature. J. Plant Prot. 2010, 37, 398–402. [Google Scholar]
- Xie, H.C.; Li, D.S.; Zhang, H.G.; Mason, C.E.; Wang, Z.Y.; Xin, L.; Cai, W.Z. Seasonal and Geographical Variation in Diapause and Cold Hardiness of the Asian Corn Borer, Ostrinia furnacalis. Insect Sci. 2015, 22, 578–586. [Google Scholar] [CrossRef] [PubMed]
- Liu, K.Q.; Wang, L.X.; Zhang, T.T.; Bai, S.X.; Wang, K.Q.; Wang, Z.Y.; He, K.L.; Hutchison, W.D. Voltine Ecotypes of the Asian Corn Borer and Their Response to Climate Warming. Insects 2021, 12, 232. [Google Scholar] [CrossRef]
- Lu, X.; Zhou, D.R. On the Relationship between Voltinism and Development Duration of Asian Corn Borer in Jilin Province. J. Plant Prot. 1999, 26, 1–6. [Google Scholar] [CrossRef]
- Cheu, S.P.; Chow, D.R.; Tung, H.F.; Lee, C.H. Yield Reduction of Corn Caused by the European Corn Borer at Different Development Stages. Acta Phytophylacica Sin. 1964, 3, 307–312. [Google Scholar]
- Zhang, Z.T.; Lei, C.P.; Sun, W.; Zhang, T.S.; Dong, H.; Qian, H.T.; Cong, B. Effect of Global Warming on Generations of Ostrinia furnacalis in Shenyang Region. J. Maize Sci. 2014, 22, 3. [Google Scholar] [CrossRef]
- Lu, X.; Zhou, S.X.; Li, L.J.; Chen, L.L.; Chang, X.; Zhang, G.H. Generation Distribution Change of the Asian Corn Borer Ostrinia furnacalis (Guenée) in Jilin Province. Acta Phytophylacica Sin. 2015, 42, 978–984. [Google Scholar]
- Sparks, A.N.; Brindley, T.A.; Penny, N.D. Laboratory and Field Studies of F1 Progenies from Reciprocal Matings of Biotypes of the European Corn Borer. J. Econ. Entomol. 1966, 59, 915–921. [Google Scholar] [CrossRef]
- Showers, W.B.; Brindley, T.A.; Reed, G.L. Survival and Diapause Characteristics of Hybrids of Three Geographical Races of the European Corn Borer. Ann. Entomol. Soc. Am. Vol. 1972, 65, 450–457. [Google Scholar] [CrossRef]
- Showers, W.B.; Chiang, H.C.; Keaster, A.J.; Hill, R.E.; Reed, G.L.; Sparks, A.N.; Musick, G.J. Ecotypes of the European Corn Borer in North America. Environ. Entomol. 1975, 4, 753–760. [Google Scholar] [CrossRef]
- Mcleod, D.G.R. Genetics of Diapause Induction and Termination in the European Corn Borer, Ostrinia Nubilalis (Lepidoptera: Pyralidae), in Southwestern Ontario. Can. Entomol. 1978, 110, 1351–1353. [Google Scholar] [CrossRef]
- Reed, G.L.; Guthrie, W.D.; Showers, W.B.; Barry, B.D.; Cox, D.F. Sex-Linked Inheritance of Diapause in the European Corn Borer: Its Significance to Diapause Physiology and Environmental Response of the Insect. Ann. Entomol. Soc. Am. 1981, 74, 1–8. [Google Scholar] [CrossRef]
- Ikten, C.; Skoda, S.R.; Hunt, T.E.; Molina-Ochoa, J.; Foster, J.E. Genetic Variation and Inheritance of Diapause Induction in Two Distinct Voltine Ecotypes of Ostrinia Nubilalis (Lepidoptera: Crambidae). Ann. Entomol. Soc. Am. 2011, 104, 567–575. [Google Scholar] [CrossRef] [Green Version]
- Jin, R.H.; Zhang, J.X. A Preliminary Study on the Diapause Ratios of Different Generations of the Natural Population of Asian Corn Borer in Beijing. Acta Agric. Univ. Pekin 1983, 1, 107–108. [Google Scholar]
- Chen, P.; Gong, H.; Wang, F.R.; Yan, Y. Studies on the Relationship between Photoperiod, Temperature and Diapause Development of Asian Corn Borer, Ostrinia furnacalis. J. Beijing Univ. Agric. 1986, 1–5. [Google Scholar]
- Beck, S.D.; Apple, J.W. Effects of Temperature and Photoperiod on Voltinism of Geographical Populations of the European Corn Borer, Pyrausta Nubilalis. J. Econ. Entomol. 1961, 54, 550–558. [Google Scholar] [CrossRef]
- Mcleod, D.G.R.; Beck, S.D. Photoperiodic Termination of Diapause in an Insect. Biol. Bull. 1963, 124, 84–96. [Google Scholar] [CrossRef]
- Zhou, D.R.; Ye, Z.H.; Wang, Z.Y. Artificial Rearing Technique for Asian Corn Borer, Ostrinia furnacalis (Guenée) and Its Applications in Pest Management Research; Westview Press: San Francisco, CA, USA, 1992. [Google Scholar]
- Showers, W.B.; Keaster, A.J.; Witkowski, J.F.; Clement, S.L.; Chiang, H.C.; Sparks, A.N. Manipulation of Larval Diapause of the European Corn Borer (Lepidoptera: Pyralidae) as a Potential Mechanism of Integrated Pest Management. Environ. Entomol. 1990, 19, 1311–1319. [Google Scholar] [CrossRef]
- Zhu, C.Y.; Zhang, Z.M. Estimation of Temperature Threshold for Development in Asian Corn Borer. Heilongjiang Agric. Sci. 1988, 26–28. [Google Scholar]
- Wang, G.Q.; Xin, Y.F. Studies on the Persistent Times and Thermal Constants of Asian Maize Borer Ostrinia furnacalis (Guenee) in Different Voltinism in Shenyang Area. J. Shenyang Agric. Univ. 2000, 31, 4. [Google Scholar] [CrossRef]
- Lu, X.; Li, J.P.; Wang, Y.S. Preliminary Study on Voltinism Types of Ostrinia furnacalis (Acb). Maize Sci. 1995, 3, 75–78. [Google Scholar]
- Arbuthnot, K.D. Strains of the European Corn Borer in the United States. Tech. Bull. 1944, 869, 1–20. [Google Scholar] [CrossRef]
- Du, Z.W.; Cai, W.Q. Preliminary Report on Response of Corn Borer to Photoperiod in Jiangsu Province. Acta Entomol. Sin. 1964, 13, 129–132. [Google Scholar]
- Kawakami, Y.; Numata, H.; Ito, K.; Goto, S.G. Dominant and Recessive Inheritance Patterns of Diapause in the Two-Spotted Spider Mite Tetranychus Urticae. J. Hered. 2010, 101, 20–25. [Google Scholar] [CrossRef]
- Zhang, H.Y.; Ge, Y.F.; Hu, X.; Dong, A.S.; Zou, C.S.; Yang, K.J.; Wang, L.Y. Occurence Dynamics of Ostrinia furnacalis in Nenjiang Region of Heilongjiang and Release of Trichogramma Dendrolimi for Its Control. Plant Prot. 2019, 45, 6. [Google Scholar] [CrossRef]






| Years | The Earliest Date of 1st Generation Egg-Laying | The Latest Date of 1st Generation Laid Eggs That Could Regenerate a Second Generation | The Earliest Date of 2nd Generation Egg-Laying | The Latest Date of 2nd Generation Laid Eggs That Could Allow Larva Develop to Mature and Enter Diapause before Winter |
|---|---|---|---|---|
| Heihe (50°14′ N, 127°14′ E) | ||||
| 1970s | 21 Jun | - | - | - |
| 1980s | 21 Jun | - | - | - |
| 1990s | 19 Jun | - | - | - |
| 2000s | 17 Jun | - | - | - |
| 2010s | 16 Jun | 18 Jun | 6 Aug | 6 Aug |
| Harbin (45°38′ N, 126°34′ E) | ||||
| 1970s | 9 Jun | 28 Jun | 28 Jul | 12 Aug |
| 1980s | 9 Jun | 29 Jun | 28 Jul | 13 Aug |
| 1990s | 8Jun | 2 Jul | 26 Jul | 13 Aug |
| 2000s | 4 Jun | 6 Jul | 21 Jul | 19 Aug |
| 2010s | 5 Jun | 7 Jul | 22 Jul | 18 Aug |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, K.; Wang, Z.; Zhang, T.; He, K. Intra-Population Alteration on Voltinism of Asian Corn Borer in Response to Climate Warming. Biology 2023, 12, 187. https://doi.org/10.3390/biology12020187
Liu K, Wang Z, Zhang T, He K. Intra-Population Alteration on Voltinism of Asian Corn Borer in Response to Climate Warming. Biology. 2023; 12(2):187. https://doi.org/10.3390/biology12020187
Chicago/Turabian StyleLiu, Kaiqiang, Zhenying Wang, Tiantao Zhang, and Kanglai He. 2023. "Intra-Population Alteration on Voltinism of Asian Corn Borer in Response to Climate Warming" Biology 12, no. 2: 187. https://doi.org/10.3390/biology12020187
APA StyleLiu, K., Wang, Z., Zhang, T., & He, K. (2023). Intra-Population Alteration on Voltinism of Asian Corn Borer in Response to Climate Warming. Biology, 12(2), 187. https://doi.org/10.3390/biology12020187