
Mother–Infant Brain-to-Brain Synchrony Patterns Reflect Caregiving Profiles

{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Participants
2.2. Paradigm
2.3. Dual-EEG Data Acquisition
2.4. EEG Preprocessing
2.5. Connectivity Analysis
2.6. Maternal Behavioral Profiles; Sensitivity and Intrusiveness
3. Results
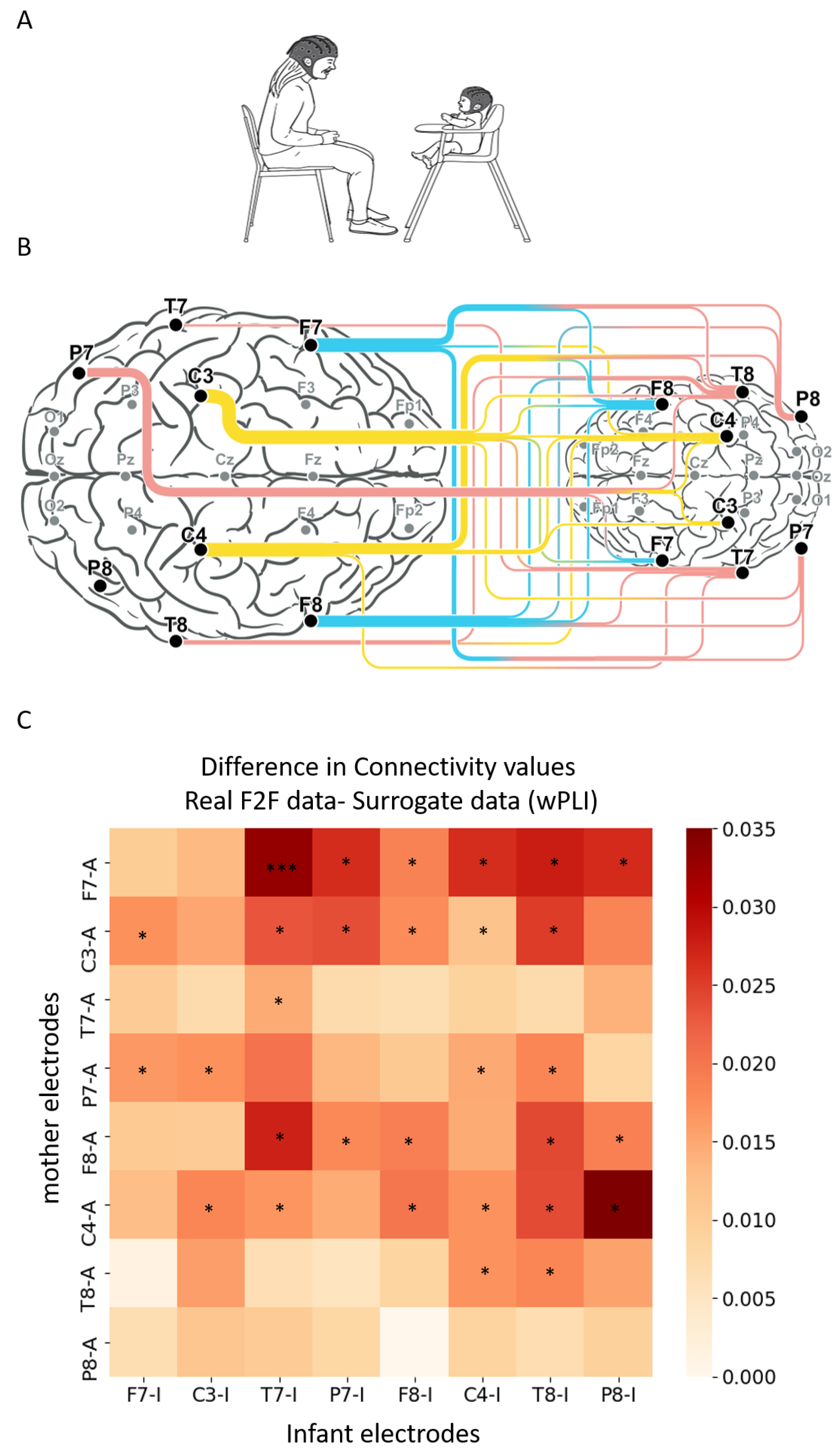
3.1. Inter-Brain Synchrony Increases during Mother–Infant Face-to-Face Interactions
3.2. Links between Inter-Brain Synchrony Patterns and Maternal Sensitivity and Intrusiveness
3.2.1. Connectivity with the Infant’s Right and Left Temporal Regions and Brain–Behavior Correlations
3.2.2. Mother’s Right and Left Frontal Regions and Brain–Behavior Correlations
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ainsworth, M.D.S.; Blehar, M.C.; Waters, E.; Wall, S. Patterns of Attachment: A Psychological Study of the Strange Situation; Hillsdale, N.J., Ed.; Lawrence Erlbaum Associates: New York, NY, USA, 1978. [Google Scholar]
- van Ijzendoorn, M.; Kroonenberg, P.M. Cross-Cultural Patterns of Attachment: A Meta-Analysis of the Strange Situation. Child Dev. 1988, 59, 147. [Google Scholar] [CrossRef]
- De Wolff, M.S.; van Ijzendoorn, M.H. Sensitivity and Attachment: A Meta-Analysis on Parental Antecedents of Infant Attachment. Child Dev. 1997, 68, 571–591. [Google Scholar] [CrossRef] [PubMed]
- Bakermans-Kranenburg, M.J.; van Ijzendoorn, M.H.; Juffer, F. Less is More: Meta-Analyses of Sensitivity and Attachment Interventions in Early Childhood. Psychol. Bull. 2003, 129, 195–215. [Google Scholar] [CrossRef] [PubMed]
- Stams, G.-J.J.M.; Juffer, F.; van Ijzendoorn, M.H. Maternal sensitivity, infant attachment, and temperament in early childhood predict adjustment in middle childhood: The case of adopted children and their biologically unrelated parents. Dev. Psychol. 2002, 38, 806–821. [Google Scholar] [CrossRef] [PubMed]
- Raby, K.L.; Roisman, G.I.; Fraley, R.C.; Simpson, J.A. The Enduring Predictive Significance of Early Maternal Sensitivity: Social and Academic Competence Through Age 32 Years. Child Dev. 2015, 86, 695–708. [Google Scholar] [CrossRef]
- Leerkes, E.M.; Blankson, A.N.; O’Brien, M. Differential Effects of Maternal Sensitivity to Infant Distress and Nondistress on Social-Emotional Functioning. Child Dev. 2009, 80, 762–775. [Google Scholar] [CrossRef]
- Feldman, R.; Eidelman, A.I.; Rotenberg, N. Parenting stress, infant emotion regulation, maternal sensitivity, and the cognitive development of triplets: A model for parent and child influences in a unique ecology. Child Dev. 2004, 75, 1774–1791. [Google Scholar] [CrossRef]
- Fraley, R.C.; Roisman, G.I.; Haltigan, J.D. The legacy of early experiences in development: Formalizing alternative models of how early experiences are carried forward over time. Dev. Psychol. 2013, 49, 109–126. [Google Scholar] [CrossRef]
- Olson, S.L.; Bates, J.E.; Sandy, J.M.; Lanthier, R. Early developmental precursors of externalizing behavior in middle childhood and adolescence. J. Abnorm. Child Psychol. 2000, 28, 119–133. [Google Scholar] [CrossRef]
- Shaw, D.S.; Keenan, K.; Vondra, J.I. Developmental Precursors of Externalizing Behavior: Ages 1 to 3. Dev. Psychol. 1994, 30, 355–364. [Google Scholar] [CrossRef]
- Egeland, B.; Farber, E.A. Infant-mother attachment: Factors related to its development and changes over time. Child Dev. 1984, 55, 753–771. [Google Scholar] [CrossRef]
- Park, K.A.; Waters, E. Security of attachment and preschool friendships. Child Dev. 1989, 60, 1076–1081. [Google Scholar] [CrossRef] [PubMed]
- Isabella, R.A.; Belsky, J. Interactional Synchrony and the Origins of Infant-Mother Attachment: A Replication Study. Child Dev. 1991, 62, 373–384. [Google Scholar] [CrossRef] [PubMed]
- Erickson, M.F.; Sroufe, L.A.; Egeland, B. The Relationship between Quality of Attachment and Behavior Problems in Preschool in a High-Risk Sample. Monogr. Soc. Res. Child Dev. 1985, 50, 147. [Google Scholar] [CrossRef] [PubMed]
- Nelson, K. Structure and Strategy in Learning to Talk. Monogr. Soc. Res. Child Dev. 1973, 38, 1–135. [Google Scholar] [CrossRef]
- Keown, L.J.; Woodward, L.J.; Field, J. Language Development of Pre-School Children Born to Teenage Mothers. Infant Child Dev. 2001, 10, 129–145. [Google Scholar] [CrossRef]
- Conway, L.J.; Levickis, P.; Smith, J.; Mensah, F.; Wake, M.; Reilly, S. Maternal communicative behaviours and interaction quality as predictors of language development: Findings from a community-based study of slow-to-talk toddlers. Int. J. Lang. Commun. Disord. 2018, 53, 339–354. [Google Scholar] [CrossRef]
- Feldman, R. Social Behavior as a Transdiagnostic Marker of Resilience. Annu. Rev. Clin. Psychol. 2021, 17, 153–180. [Google Scholar] [CrossRef]
- Belsky, J.; Taylor, D.G. The Pennsylvania Infant and Family Development Project, III: The origins of individual differences in infant-mother attachment: Maternal and infant contributions. Child Dev. 1984, 55, 718–728. [Google Scholar] [CrossRef]
- Feldman, R. The relational basis of adolescent adjustment: Trajectories of mother–child interactive behaviors from infancy to adolescence shape adolescents’ adaptation. Attach. Hum. Dev. 2010, 12, 173–192. [Google Scholar] [CrossRef]
- Yaniv, A.U.; Salomon, R.; Waidergoren, S.; Shimon-Raz, O.; Djalovski, A.; Feldman, R. Synchronous caregiving from birth to adulthood tunes humans’ social brain. Proc. Natl. Acad. Sci. USA 2021, 118. [Google Scholar] [CrossRef]
- Ulmer-Yaniv, A.; Djalovski, A.; Yirmiya, K.; Halevi, G.; Zagoory-Sharon, O.; Feldman, R. Maternal immune and affiliative biomarkers and sensitive parenting mediate the effects of chronic early trauma on child anxiety. Psychol. Med. 2018, 48, 1020–1033. [Google Scholar] [CrossRef] [PubMed]
- Yirmiya, K.; Motsan, S.; Zagoory-Sharon, O.; Feldman, R. Human attachment triggers different social buffering mechanisms under high and low early life stress rearing. Int. J. Psychophysiol. 2020, 152, 72–80. [Google Scholar] [CrossRef]
- Feldman, R.; Gordon, I.; Zagoory-Sharon, O. Maternal and paternal plasma, salivary, and urinary oxytocin and parent-infant synchrony: Considering stress and affiliation components of human bonding. Dev. Sci. 2011, 14, 752–761. [Google Scholar] [CrossRef]
- Yirmiya, K.; Djalovski, A.; Motsan, S.; Zagoory-Sharon, O.; Feldman, R. Stress and immune biomarkers interact with parenting behavior to shape anxiety symptoms in trauma-exposed youth. Psychoneuroendocrinology 2018, 98, 153–160. [Google Scholar] [CrossRef] [PubMed]
- Ulmer-Yaniv, A.; Djalovski, A.; Priel, A.; Zagoory-Sharon, O.; Feldman, R. Maternal depression alters stress and immune biomarkers in mother and child. Depress. Anxiety 2018, 35, 1145–1157. [Google Scholar] [CrossRef] [PubMed]
- Pratt, M.; Goldstein, A.; Levy, J.; Feldman, R. Maternal Depression Across the First Years of Life Impacts the Neural Basis of Empathy in Preadolescence. J. Am. Acad. Child Adolesc. Psychiatry 2017, 56, 20–29.e3. [Google Scholar] [CrossRef]
- Pratt, M.; Goldstein, A.; Feldman, R. Child brain exhibits a multi-rhythmic response to attachment cues. Soc. Cogn. Affect. Neurosci. 2018, 13, 957–966. [Google Scholar] [CrossRef]
- Levy, J.; Goldstein, A.; Feldman, R. The neural development of empathy is sensitive to caregiving and early trauma. Nat. Commun. 2019, 10, 1905. [Google Scholar] [CrossRef]
- Zeev-Wolf, M.; Dor-Ziderman, Y.; Pratt, M.; Goldstein, A.; Feldman, R. Investigating default mode network connectivity disruption in children of mothers with depression. Br. J. Psychiatry 2022, 220, 130–139. [Google Scholar] [CrossRef]
- Zeev-Wolf, M.; Levy, J.; Goldstein, A.; Zagoory-Sharon, O.; Feldman, R. Chronic Early Stress Impairs Default Mode Network Connectivity in Preadolescents and Their Mothers. Biol. Psychiatry Cogn. Neurosci. Neuroimaging 2019, 4, 72–80. [Google Scholar] [CrossRef] [PubMed]
- Deoni, S.C.L.; Mercure, E.; Blasi, A.; Gasston, D.; Thomson, A.; Johnson, M.; Williams, S.C.R.; Murphy, D.G.M. Mapping infant brain myelination with magnetic resonance imaging. J. Neurosci. 2011, 31, 784–791. [Google Scholar] [CrossRef] [PubMed]
- Lemaître, H.; Augé, P.; Saitovitch, A.; Vinçon-Leite, A.; Tacchella, J.-M.; Fillon, L.; Calmon, R.; Dangouloff-Ros, V.; Lévy, R.; Grévent, D.; et al. Rest Functional Brain Maturation during the First Year of Life. Cereb. Cortex 2020, 31, 1776–1785. [Google Scholar] [CrossRef]
- Gao, W.; Alcauter, S.; Elton, A.; Hernandez-Castillo, C.R.; Smith, J.K.; Ramirez, J.; Lin, W. Functional network development during the first year: Relative sequence and socioeconomic correlations. Cereb. Cortex 2015, 25, 2919–2928. [Google Scholar] [CrossRef]
- van der Velde, B.; White, T.; Kemner, C. The emergence of a theta social brain network during infancy. Neuroimage 2021, 240, 118298. [Google Scholar] [CrossRef] [PubMed]
- Feldman, R. Parent-infant synchrony: A biobehavioral model of mutual influences in the formation of affiliative bonds. Monogr. Soc. Res. Child Dev. 2012, 77, 42–51. [Google Scholar] [CrossRef]
- Feldman, R. The neurobiology of mammalian parenting and the biosocial context of human caregiving. Horm. Behav. 2016, 77, 3–17. [Google Scholar] [CrossRef]
- Feldman, R. What is resilience: An affiliative neuroscience approach. World Psychiatry 2020, 19, 132–150. [Google Scholar] [CrossRef]
- Feldman, R. The Neurobiology of Human Attachments. Trends Cogn. Sci. 2017, 21, 80–99. [Google Scholar] [CrossRef]
- Nguyen, T.; Schleihauf, H.; Kayhan, E.; Matthes, D.; Vrtička, P.; Hoehl, S. Neural synchrony in mother–child conversation: Exploring the role of conversation patterns. Soc. Cogn. Affect. Neurosci. 2021, 16, 93–102. [Google Scholar] [CrossRef]
- Nguyen, T.; Schleihauf, H.; Kayhan, E.; Matthes, D.; Vrtička, P.; Hoehl, S. The effects of interaction quality on neural synchrony during mother-child problem solving. Cortex 2019, 124, 235–249. [Google Scholar] [CrossRef] [PubMed]
- Endevelt-Shapira, Y.; Djalovski, A.; Dumas, G.; Feldman, R. Maternal chemosignals Enhance Infant-Adult Brain-to-Brain Synchrony. Sci. Adv. 2021, 7, eabg6867. [Google Scholar] [CrossRef] [PubMed]
- Schwartz, L.; Levy, J.; Endevelt-Shapira, Y.; Djalovski, A.; Hayut, O.; Dumas, G.; Feldman, R. Technologically-assisted communication attenuates inter-brain synchrony. Neuroimage 2022, 264, 119677. [Google Scholar] [CrossRef] [PubMed]
- Lin, J.-F.L.; Imada, T.; Meltzoff, A.N.; Hiraishi, H.; Ikeda, T.; Takahashi, T.; Hasegawa, C.; Yoshimura, Y.; Kikuchi, M.; Hirata, M.; et al. Dual-MEG interbrain synchronization during turn-taking verbal interactions between mothers and children. Cereb. Cortex 2022, bhac330. [Google Scholar] [CrossRef]
- Kaitz, M.; Maytal, H. Interactions between anxious mothers and their infants: An integration of theory and research findings. Infant Ment. Health J. 2005, 26, 570–597. [Google Scholar] [CrossRef]
- Feldman, R. Sensitive periods in human social development: New insights from research on oxytocin, synchrony, and high-risk parenting. Dev. Psychopathol. 2015, 27, 369–395. [Google Scholar] [CrossRef]
- Feldman, R. Coding Interactive Behavior; BarIlan University Israel: Ramat Gan, Israel, 1998. [Google Scholar]
- Djalovski, A.; Dumas, G.; Kinreich, S.; Feldman, R. Human Attachments Shape Interbrain Synchrony toward Efficient Performance of Social Goals. Neuroimage 2020, 226, 117600. [Google Scholar] [CrossRef]
- Jas, M.; Engemann, D.A.; Bekhti, Y.; Raimondo, F.; Gramfort, A. Autoreject: Automated artifact rejection for MEG and EEG data. Neuroimage 2017, 159, 417–429. [Google Scholar] [CrossRef]
- Viola, F.C.; Thorne, J.; Edmonds, B.; Schneider, T.R.; Eichele, T.; Debener, S. Semi-automatic identification of independent components representing EEG artifact. Clin. Neurophysiol. 2009, 120, 868–877. [Google Scholar] [CrossRef]
- Orekhova, E.; Stroganova, T.; Posikera, I.; Elam, M. EEG theta rhythm in infants and preschool children. Clin. Neurophysiol. 2006, 117, 1047–1062. [Google Scholar] [CrossRef]
- Dikker, S.; Michalareas, G.; Oostrik, M.; Serafimaki, A.; Kahraman, H.M.; Struiksma, M.E.; Poeppel, D. Crowdsourcing neuroscience: Inter-brain coupling during face-to-face interactions outside the laboratory. Neuroimage 2021, 227, 117436. [Google Scholar] [CrossRef] [PubMed]
- Vinck, M.; Oostenveld, R.; van Wingerden, M.; Battaglia, F.; Pennartz, C.M.A. An improved index of phase-synchronization for electrophysiological data in the presence of volume-conduction, noise and sample-size bias. NeuroImage 2011, 55, 1548–1565. [Google Scholar] [CrossRef] [PubMed]
- Kinreich, S.; Djalovski, A.; Kraus, L.; Louzoun, Y.; Feldman, R. Brain-to-Brain Synchrony during Naturalistic Social Interactions. Sci. Rep. 2017, 7, 1–12. [Google Scholar] [CrossRef]
- Hirsch, J.; Zhang, X.; Noah, J.A.; Ono, Y. Frontal temporal and parietal systems synchronize within and across brains during live eye-to-eye contact. Neuroimage 2017, 157, 314–330. [Google Scholar] [CrossRef] [PubMed]
- Leong, V.; Byrne, E.; Clackson, K.; Georgieva, S.; Lam, S.; Wass, S. Speaker gaze increases information coupling between infant and adult brains. Proc. Natl. Acad. Sci. USA 2017, 114, 13290–13295. [Google Scholar] [CrossRef]
- Feldman, R. Parenting Behavior as the Environment Where Children Grow. In The Cambridge Handbook of Environment in Human Development; Mayes, L., Lewis, M., Eds.; Cambridge University Press: Cambridge, UK, 2012; pp. 535–567. [Google Scholar] [CrossRef]
- Feldman, R.; Eidelman, A.I. Biological and environmental initial conditions shape the trajectories of cognitive and social-emotional development across the first years of life. Dev. Sci. 2009, 12, 194–200. [Google Scholar] [CrossRef]
- Pratt, M.; Zeev-Wolf, M.; Goldstein, A.; Feldman, R. Exposure to early and persistent maternal depression impairs the neural basis of attachment in preadolescence. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2019, 93, 21–30. [Google Scholar] [CrossRef]
- Santamaria, L.; Noreika, V.; Georgieva, S.; Clackson, K.; Wass, S.; Leong, V. Emotional valence modulates the topology of the parent-infant inter-brain network. Neuroimage 2020, 207, 116341. [Google Scholar] [CrossRef]
- Feldman, R. Bio-behavioral synchrony: A model for integrating biological and microsocial behavioral processes in the study of parenting. Parenting 2012, 12, 154–164. [Google Scholar] [CrossRef]
- Wass, S.V.; Whitehorn, M.; Haresign, I.M.; Phillips, E.; Leong, V. Interpersonal Neural Entrainment during Early Social Interaction. Trends Cogn. Sci. 2020, 24, 329–342. [Google Scholar] [CrossRef]
- Carozza, S.; Leong, V. The Role of Affectionate Caregiver Touch in Early Neurodevelopment and Parent–Infant Interactional Synchrony. Front. Neurosci. 2021, 14. [Google Scholar] [CrossRef]
- Levy, J.; Goldstein, A.; Feldman, R. Perception of social synchrony induces mother–child gamma coupling in the social brain. Soc. Cogn. Affect. Neurosci. 2017, 12, 1036–1046. [Google Scholar] [CrossRef] [PubMed]
- Frith, U.; Frith, C. The biological basis of social interaction. Curr. Dir. Psychol. Sci. 2001, 10, 151–155. [Google Scholar] [CrossRef]
- Atzil, S.; Hendler, T.; Zagoory-Sharon, O.; Winetraub, Y.; Feldman, R. Synchrony and specificity in the maternal and the paternal brain: Relations to oxytocin and vasopressin. J. Am. Acad. Child Adolesc. Psychiatry 2012, 51, 798–811. [Google Scholar] [CrossRef]
- Urbain, C.; Vogan, V.M.; Ye, A.X.; Pang, E.W.; Doesburg, S.M.; Taylor, M.J. Desynchronization of fronto-temporal networks during working memory processing in autism. Hum. Brain Mapp. 2016, 37, 153–164. [Google Scholar] [CrossRef]
- A Hofer, M. Early relationships as regulators of infant physiology and behavior. Acta Paediatr. 1994, 83, 9–18. [Google Scholar] [CrossRef] [PubMed]
- Cirulli, F.; Berry, A.; Alleva, E. Early disruption of the mother–infant relationship: Effects on brain plasticity and implications for psychopathology. Neurosci. Biobehav. Rev. 2003, 27, 73–82. [Google Scholar] [CrossRef]
- Feldman, R. The adaptive human parental brain: Implications for children’s social development. Trends Neurosci. 2015, 38, 387–399. [Google Scholar] [CrossRef]
- Feldman, R.; Granat, A.; Pariente, C.; Kanety, H.; Kuint, J.; Gilboa-Schechtman, E. Maternal Depression and Anxiety Across the Postpartum Year and Infant Social Engagement, Fear Regulation, and Stress Reactivity. J. Am. Acad. Child Adolesc. Psychiatry 2009, 48, 919–927. [Google Scholar] [CrossRef] [Green Version]
- Feldman, R. Parent-infant synchrony and the construction of shared timing; physiological precursors, developmental outcomes, and risk conditions. J. Child Psychol. Psychiatry 2007, 48, 329–354. [Google Scholar] [CrossRef]
- Shultz, S.; Vouloumanos, A.; Pelphrey, K. The Superior Temporal Sulcus Differentiates Communicative and Noncommunicative Auditory Signals. J. Cogn. Neurosci. 2012, 24, 1224–1232. [Google Scholar] [CrossRef] [PubMed]
- Redcay, E. The superior temporal sulcus performs a common function for social and speech perception: Implications for the emergence of autism. Neurosci. Biobehav. Rev. 2008, 32, 123–142. [Google Scholar] [CrossRef] [PubMed]
- Kuhl, P.K. Brain Mechanisms in Early Language Acquisition. Neuron 2010, 67, 713–727. [Google Scholar] [CrossRef] [PubMed]
- Huffmeijer, R.; Bakermans-Kranenburg, M.J.; Gervain, J. Maternal intrusiveness predicts infants’ event-related potential responses to angry and happy prosody independent of infant frontal asymmetry. Infancy 2020, 25, 246–263. [Google Scholar] [CrossRef] [PubMed] [Green Version]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Endevelt-Shapira, Y.; Feldman, R. Mother–Infant Brain-to-Brain Synchrony Patterns Reflect Caregiving Profiles. Biology 2023, 12, 284. https://doi.org/10.3390/biology12020284
Endevelt-Shapira Y, Feldman R. Mother–Infant Brain-to-Brain Synchrony Patterns Reflect Caregiving Profiles. Biology. 2023; 12(2):284. https://doi.org/10.3390/biology12020284
Chicago/Turabian StyleEndevelt-Shapira, Yaara, and Ruth Feldman. 2023. "Mother–Infant Brain-to-Brain Synchrony Patterns Reflect Caregiving Profiles" Biology 12, no. 2: 284. https://doi.org/10.3390/biology12020284
APA StyleEndevelt-Shapira, Y., & Feldman, R. (2023). Mother–Infant Brain-to-Brain Synchrony Patterns Reflect Caregiving Profiles. Biology, 12(2), 284. https://doi.org/10.3390/biology12020284