


Defining Patterns and Rates of Natural vs. Drought Driven Aquatic Community Variability Indicates the Ongoing Need for Long Term Ecological Research



Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Field Sampling and Laboratory Analysis
2.3. Statistical Analyses
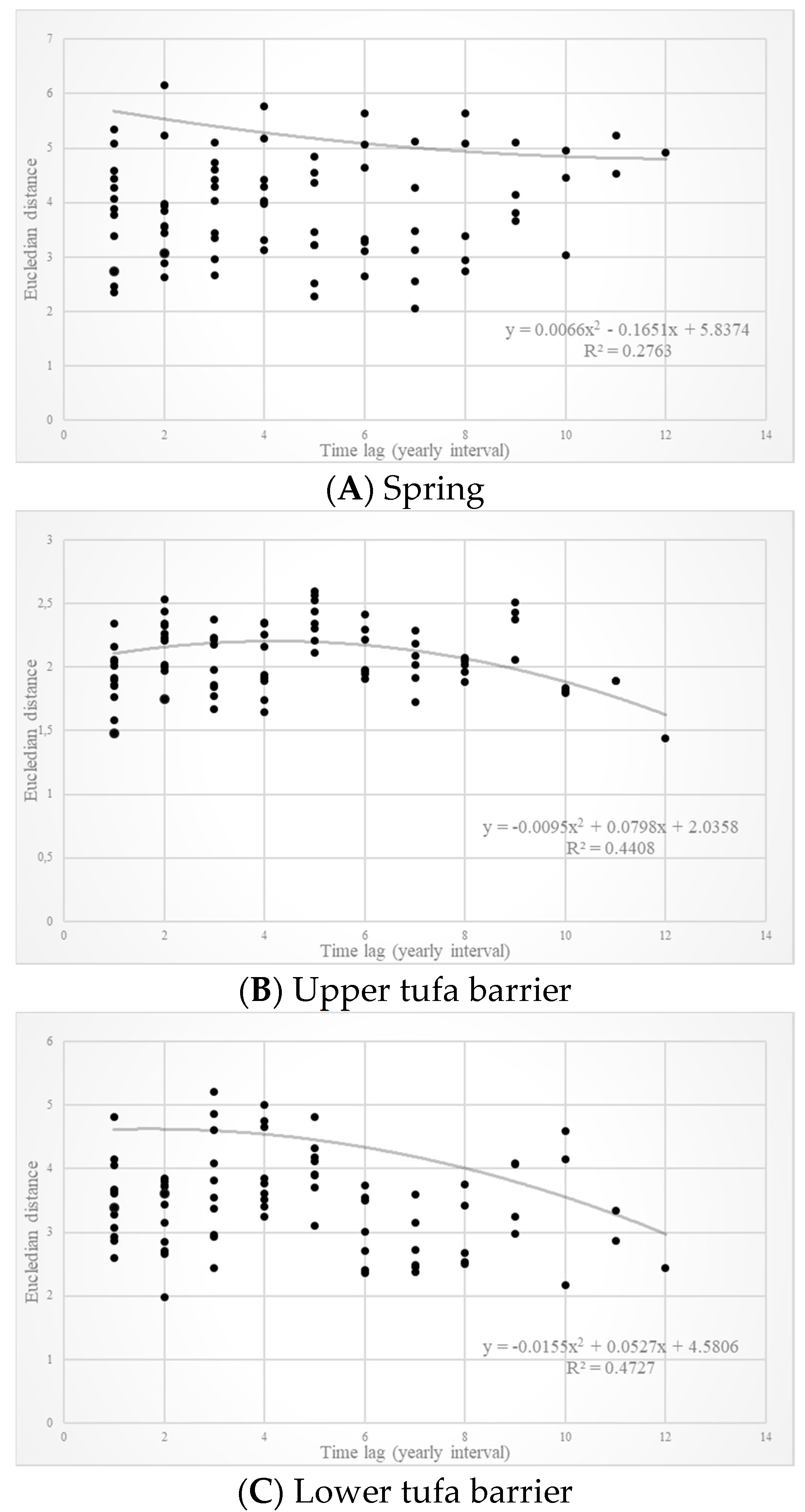
3. Results
3.1. Spring
3.2. Upper Tufa Barrier
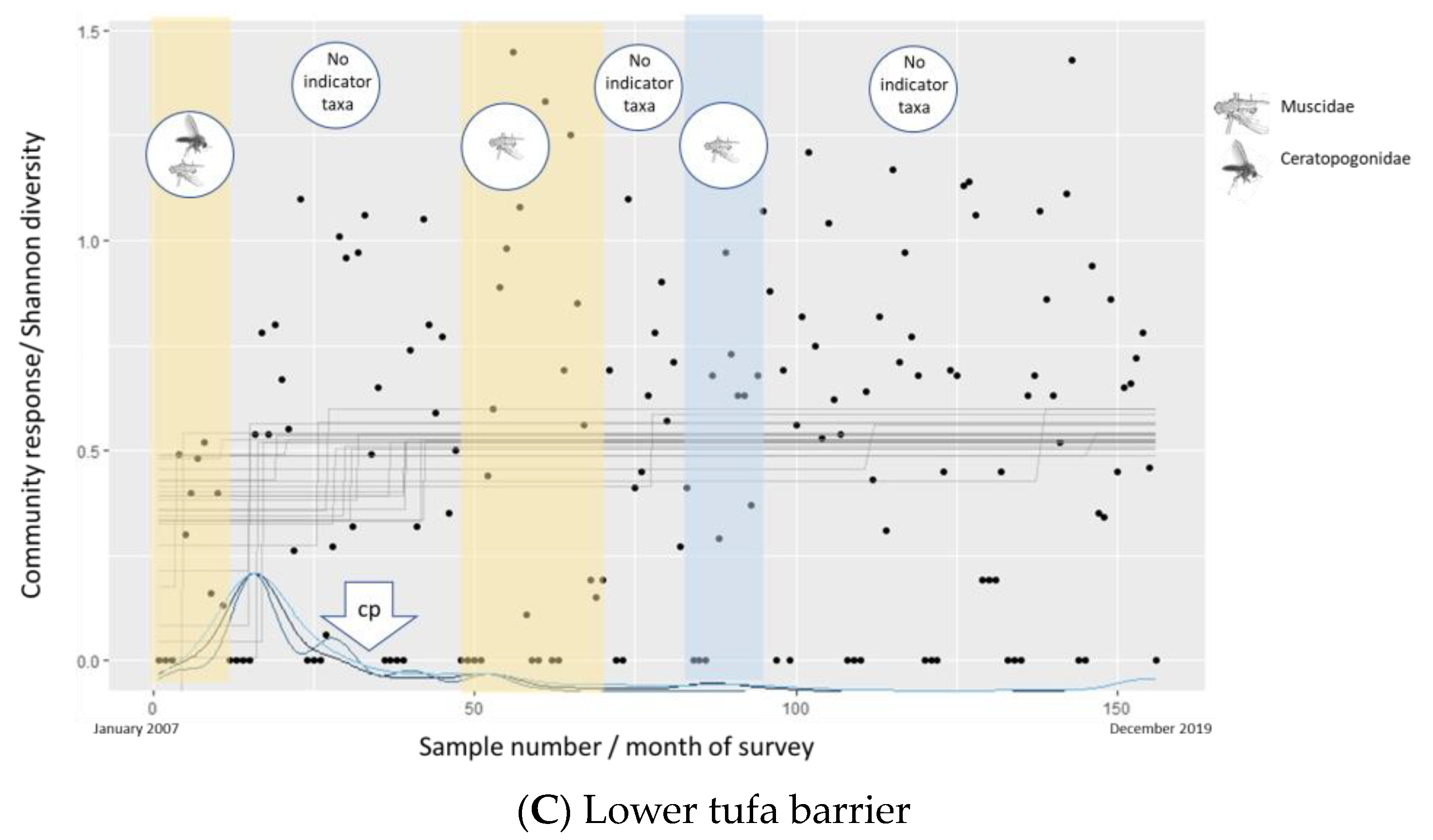
3.3. Lower Tufa Barrier
4. Discussion
4.1. Spring
4.2. Tufa Barriers
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ormerod, S.J.; Dobson, M.; Hildrew, A.G.; Townsend, C.R. Multiple stressors in freshwater ecosystems. Freshw. Biol. 2010, 55, 1–4. [Google Scholar] [CrossRef]
- Jackson, M.C.; Loewen, C.J.G.; Vinebrooker, R.D.; Chimimba, C.T. Net effects of multiple stressors in freshwater ecosystems: A meta-analysis. Glob. Change Biol. 2016, 22, 180–189. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Čmrlec, K.; Ivković, M.; Šemnički, P.; Mihaljević, Z. Emergence phenology and microhabitat distribution of aquatic Diptera community at the outlets of barrage lakes: Effect of temperature, substrate and current velocity. Pol. J. Ecol. 2013, 61, 135–144. [Google Scholar]
- Ivanković, L.; Ivković, M.; Stanković, I. Perennial phenology patterns and ecological traits of Dixidae (Insecta, Diptera) in lotic habitats of a barrage lake system. Limnologica 2019, 76, 11–18. [Google Scholar] [CrossRef]
- Ivković, M.; Mičetić Stanković, V.; Mihaljević, Z. Emergence patterns and microhabitat preference of aquatic dance flies (Empididae; Clinocerinae and Hemerodromiinae) on a longitudinal gradient of barrage lake system. Limnologica 2012, 42, 43–49. [Google Scholar] [CrossRef]
- Ivković, M.; Miliša, M.; Previšić, A.; Popijač, A.; Mihaljević, Z. Environmental factors and emergence patterns: Case study of changes in hourly and daily emergence of aquatic insects at constant and variable water temperatures. Int. Rev. Hydrobiol. 2013, 98, 104–115. [Google Scholar] [CrossRef]
- Ivković, M.; Kesić, M.; Mihaljević, Z.; Kudelá, M. Emergence patterns and ecological associations of hematophagous black flies along oligotrophic hydrosystem. Med. Vet. Entomol. 2014, 28, 94–102. [Google Scholar] [CrossRef] [PubMed]
- Ivković, M.; Miliša, M.; Baranov, V.; Mihaljević, Z. Environmental drivers of biotic traits and phenology patterns of Diptera assemblages in karst springs: The role of canopy uncovered. Limnologica 2015, 54, 44–57. [Google Scholar] [CrossRef]
- Ivković, M.; Pont, A.C. Long-time emergence patterns of Limnophora species (Diptera, Muscidae) in specific karst habitats: Tufa barriers. Limnologica 2016, 61, 29–35. [Google Scholar] [CrossRef]
- Pozojević, I.; Ivković, M.; Cetinić, K.A.; Previšić, A. Peeling the Layers of Caddisfly Diversity on a Longitudinal Gradient in Karst Freshwater Habitats Reveals Community Dynamics and Stability. Insects 2021, 12, 234. [Google Scholar] [CrossRef]
- Ridl, A.; Vilenica, M.; Ivković, M.; Popijač, A.; Sivec, I.; Miliša, M.; Mihaljević, Z. Environmental drivers influencing stonefly assemblages along a longitudinal gradient in karst lotic habitats. J. Limnol. 2018, 77, 412–427. [Google Scholar] [CrossRef]
- Šemnički, P.; Previšić, A.; Ivković, M.; Čmrlec, K.; Mihaljević, Z. Tufa barriers from a Caddisfly’s point of view: Streams or Lake outlets? Int. Rev. Hydrobiol. 2012, 97, 465–484. [Google Scholar] [CrossRef]
- Vilenica, M.; Ivković, M.; Sartori, M.; Mihaljević, Z. Mayfly emergence along an oligotrophic Dinaric karst hydrosystem: Spatial and temporal patterns, and species–environment relationship. Aquat. Ecol. 2017, 51, 417–433. [Google Scholar] [CrossRef]
- Van den Besselaar, E.J.M.; Klein Tank, A.M.G.; Buishand, T.A. Trends in European precipitation extremes over 1951–2010. Int. J. Climatol. 2013, 33, 2682–2689. [Google Scholar] [CrossRef]
- Branković, Č.; Patarčić, M.; Güttler, I.; Srnec, L. Near-future climate change over Europe and Croatia in an ensemble of regional climate model simulations. Clim. Res. 2012, 52, 227–251. [Google Scholar] [CrossRef] [Green Version]
- Cindrić, K.; Telišman Prtenjak, M.; Herceg-Bulić, I.; Mihajlović, D.; Pasarić, Z. Analysis of the extraordinary 2011/2012 drought in Croatia. Theor. Appl. Climatol. 2016, 123, 503–522. [Google Scholar] [CrossRef]
- Walters, A.W.; Post, D.P. How Low Can You Go? Impacts of a Low-Flow Disturbance on Aquatic Insect Communities. Ecol. Appl. 2011, 21, 163–174. [Google Scholar] [CrossRef]
- Wood, P.J.; Armitage, P.D. The Response of the Macroinvertebrate Community to Low-Flow Variability and Supra-Seasonal Drought within a Groundwater Dominated Stream. Archiv. Hydrobiol. 2004, 161, 1–20. [Google Scholar] [CrossRef]
- Lake, P.S. Resistance, Resilience and Restoration. Ecol. Manag. Restor. 2013, 14, 20–24. [Google Scholar] [CrossRef]
- Adler, P.H.; Courtney, G.W. Ecological and Societal Services of Aquatic Diptera. Insects 2019, 10, 70. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marshall, S.A. Flies: The Natural History and Diversity of Diptera, 1st ed.; Firefly Books: Richmond Hill, ON, Canada, 2012. [Google Scholar]
- Dorić, V.; Pozojević, I.; Vučković, N.; Ivković, M.; Mihaljević, Z. Lentic chironomid performance in species-based bioassessment proving: High-level taxonomy is not a dead end in monitoring. Ecol. Indic. 2021, 121, 107041. [Google Scholar] [CrossRef]
- Collins, S.L.; Micheli, F.; Hartt, L. A method to determine rates and patterns of variability in ecological communities. Oikos 2000, 91, 285–293. [Google Scholar] [CrossRef]
- Stilinović, B.; Božičević, S. The Plitvice lakes. A natural phenomen in the middle of the Dinaridic Karst in Croatia. Eur. Water Manag. 1998, 1, 15–24. [Google Scholar]
- Šegota, T.; Filipčić, A. Köppenova podjela klima i hrvatsko nazivlje. Geoadria 2003, 8, 17–23. (In Croatian) [Google Scholar] [CrossRef] [Green Version]
- Ivković, M.; Dorić, V.; Baranov, V.; Mihaljević, Z.; Kolcsár, L.P.; Kvifte, G.M.; Nerudova, J.; Pont, A.C. Checklist of aquatic Diptera (Insecta) of Plitvice Lakes National Park, Croatia, a UNESCO world heritage site. ZooKeys 2020, 918, 99–142. [Google Scholar] [CrossRef]
- Nilsson, A. Aquatic Insects of North Europe: A Taxonomic Handbook; Volume 2: Odonata—Diptera; Apollo Books: Stenstrup, Denmark, 1997. [Google Scholar]
- Oosterbroek, P. The European Families of the Diptera: Identification, Diagnosis, Biology; KNNV Publishing: Utrecht, The Netherlands, 2006. [Google Scholar]
- TIBCO Software Inc. Statistica (Data Analysis Software System), Version 13. Available online: http://statistica.io (accessed on 7 March 2021).
- Clarke, K.R.; Gorley, R.N. PRIMER v7: User Manual/Tutorial; PRIMER-E: Plymouth, UK, 2015. [Google Scholar]
- Lindeløv, J.K. mcp: An R Package for Regression with Multiple Change Points. OSF Prepr. 2020. [Google Scholar] [CrossRef]
- De Caceres, M.; Legendre, P. Associations between species and groups of sites: Indices and statistical inference. Ecology 2009, 90, 3566–3574. [Google Scholar] [CrossRef] [PubMed]
- Lake, P.S. Human-Induced Exacerbation of Drought Effects on Aquatic Ecosystems. In Drought and Aquatic Ecosystems: Effects and Responses; Wiley-Blackwell: Oxford, UK, 2011; pp. 265–289. [Google Scholar] [CrossRef]
- McDonnell, J.J. A Rationale for Old Water Discharge through Macropores in a Steep, Humid Catchment. Water Resour. Res. 1990, 26, 2821–2832. [Google Scholar] [CrossRef]
- Takhteev, V.V.; Galimzyanova, A.V.; Ambrosova, E.V.; Kravtsova, L.S.; Rozhkova, N.A.; Okuneva, G.L.; Semerno, V.P.; Pomazkova, G.I.; Lopatovskaya, O.G. Zoobenthos Communities and Their Seasonal Dynamics in Nonfreezing Springs of Baikal Region. Biol. Bull. 2010, 37, 638–646. [Google Scholar] [CrossRef]
- Dolezil, Z.; Rozkošný, R. Diptera syrphidae, Hover Flies. In Aquatic Insects of North Europe; Nilsson, A., Ed.; ApolloBooks: Stenstrup, Denmark, 1997; Volume 2, pp. 347–361. [Google Scholar]
- Cervi, F.; Petronicia, F.; Castellarin, A.; Marcaccio, M.; Bertolini, A.; Borgatti, L. Climate-change potential effects on the hydrological regime of freshwater springs in the Italian Northern Apennines. Sci. Total Environ. 2018, 622–623, 337–348. [Google Scholar] [CrossRef]
- Pozojević, I.; Brigić, A.; Gottstein, S. Water mite (Acari: Hydrachnidia) diversity and distribution in undisturbed Dinaric karst springs. Exp. Appl. Acarol. 2018, 76, 123–138. [Google Scholar] [CrossRef]
- Miliša, M.; Belančić, A.; Matoničkin Kepčija, R.; Sertić-Perić, M.; Ostojić, A.; Habdija, I. Calcite deposition in karst waters is promoted by leaf litter breakdown and vice versa. Int. J. Limnol. 2010, 46, 225–232. [Google Scholar] [CrossRef]
- Hofsvang, T. Diptera tipulidae, Crane Flies. In Aquatic Insects of North Europe; Nilsson, A., Ed.; ApolloBooks: Stenstrup, Denmark, 1997; Volume 2, pp. 93–98. [Google Scholar]
- Eveleens, R.A.; McIntosh, A.R.; Warburton, H.J. Interactive community responses to disturbance in streams: Disturbance history moderates the influence of disturbance types. Oikos 2019, 128, 1170–1181. [Google Scholar] [CrossRef]
- Amundrud, S.L.; Srivastava, D.S. Trophic interactions determine the effects of drought on an aquatic ecosystem. Ecology 2016, 97, 1475–1483. [Google Scholar] [CrossRef] [PubMed]
- Drummond, L.R.; McIntosh, A.R.; Larned, S.T. Invertebrate community dynamics and insect emergence in response to pool drying in a temporary river. Freshw. Biol. 2015, 60, 1596–1612. [Google Scholar] [CrossRef]
- Chase, J.M.; Knight, T.M. Drought-induced mosquito outbreaks in wetlands. Ecol. Lett. 2003, 6, 1017–1024. [Google Scholar] [CrossRef] [Green Version]
- Baranov, V.; Jourdan, J.; Pilotto, F.; Wagner, R.; Haase, P. Complex and nonlinear climate-driven changes in freshwater insect communities over 42 years. Conserv. Biol. 2020, 34, 1241–1251. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Site | IBR | BL | BKM | |
|---|---|---|---|---|
| Latitude | N 44°50″05′ | N 44°52″17′ | N 44°53″39′ | |
| Longitude | E 15°33″43′ | E 15°35″59′ | E 15°36″32′ | |
| Longitudinal position/habitat type in karst barrage lake ecosystem | Spring | Upper barrier | Lower barrier | |
| Altitude (m) | 720 | 630 | 546 | |
| Substrate | Pebbles and sand, Macrophytes, Moss | Pebbles, Moss on tufa, Tufa with detritus | Pebbles, Moss on tufa, Tufa with detritus, Silt | |
| Water temperature (°C) without 2011/2012 | Min | 6.8 | 1.848233871 | 1.91925 |
| Average ± SD | 7.884 ± 0.48 | 10.900 ± 5.45 | 12.296 ± 6.53 | |
| CV | 6% | 50% | 53% | |
| Max | 9.9 | 20.33 | 22.95 | |
| Water temperature (°C) in 2011/2012 | Min | 7.2 | 1.663 | 1.673 |
| Average ± SD | 7.846 ± 0.48 | 11.078 ± 5.45 | 12.603 ± 6.53 | |
| CV | 5% | 57% | 58% | |
| Max | 8.6 | 20.063 | 22.683 | |
| Water temperature (°C) in vegetative season (April–October) without 2011/2012 | Min | 7.2 | 8.35 | 8.18 |
| Average ± SD | 8.117 ± 0.45 | 14.655 ± 3.62 | 16.776 ± 4.17 | |
| CV | 6% | 25% | 25% | |
| Max | 9.9 | 20.33 | 22.95 | |
| Water temperature (°C) in vegetative season (April–October) 2011/2012 | Min | 7.3 | 9.167 | 9.666 |
| Average ± SD | 8.064 ± 0.39 | 15.629 ± 3.85 | 17.721 ± 4.39 | |
| CV | 5% | 25% | 25% | |
| Max | 8.6 | 20.063 | 22.683 | |
| Discharge (m3/s) without 2011/2012 | Min | 0.001 | 0.495 | 0.613 |
| Average ± SD | 0.547 ± 0.42 | 2.464 ± 1.893 | 3.211 ± 2.06 | |
| CV | 77% | 77% | 64% | |
| Max | 2.83 | 15.6 | 29.8 | |
| Discharge (m3/s) in 2011/2012 | Min | 0.011 | 0.525 | 0.625 |
| Average ± SD | 0.225 ± 0.201 | 1.349 ± 0.794 | 1.595 ± 0.845 | |
| CV | 89% | 59% | 53% | |
| Max | 0.909 | 6.36 | 7.23 | |
| Discharge (m3/s) in vegetative season (April–October) without 2011/2012 | Min | 0.001 | 0.495 | 0.6781 |
| Average ± SD | 0.502 ± 0.43 | 2.008 ± 1.79 | 2.688 ± 2.03 | |
| CV | 85% | 89% | 76% | |
| Max | 2.83 | 15.6 | 8.794 | |
| Discharge (m3/s) in vegetative season (April–October) 2011/2012 | Min | 0.033 | 0.525 | 0.776 |
| Average ± SD | 0.190 ± 0.13 | 1.077 ± 0.47 | 1.354 ± 0.51 | |
| CV | 70% | 43% | 38% | |
| Max | 0.578 | 3.6 | 2.461 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pozojević, I.; Dorić, V.; Miliša, M.; Ternjej, I.; Ivković, M. Defining Patterns and Rates of Natural vs. Drought Driven Aquatic Community Variability Indicates the Ongoing Need for Long Term Ecological Research. Biology 2023, 12, 590. https://doi.org/10.3390/biology12040590
Pozojević I, Dorić V, Miliša M, Ternjej I, Ivković M. Defining Patterns and Rates of Natural vs. Drought Driven Aquatic Community Variability Indicates the Ongoing Need for Long Term Ecological Research. Biology. 2023; 12(4):590. https://doi.org/10.3390/biology12040590
Chicago/Turabian StylePozojević, Ivana, Valentina Dorić, Marko Miliša, Ivančica Ternjej, and Marija Ivković. 2023. "Defining Patterns and Rates of Natural vs. Drought Driven Aquatic Community Variability Indicates the Ongoing Need for Long Term Ecological Research" Biology 12, no. 4: 590. https://doi.org/10.3390/biology12040590
APA StylePozojević, I., Dorić, V., Miliša, M., Ternjej, I., & Ivković, M. (2023). Defining Patterns and Rates of Natural vs. Drought Driven Aquatic Community Variability Indicates the Ongoing Need for Long Term Ecological Research. Biology, 12(4), 590. https://doi.org/10.3390/biology12040590