
Lunar Cycle, Climate, and Onset of Parturition in Domestic Dromedary Camels: Implications of Species-Specific Metabolic Economy and Social Ecology

 , and
, and 
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animal Sample: Candidate Selection
2.2. Farming Environment: A Historical Background
2.3. Moon Phase and Weather Variables: Data Collection
2.4. Statistical Analyses
3. Results
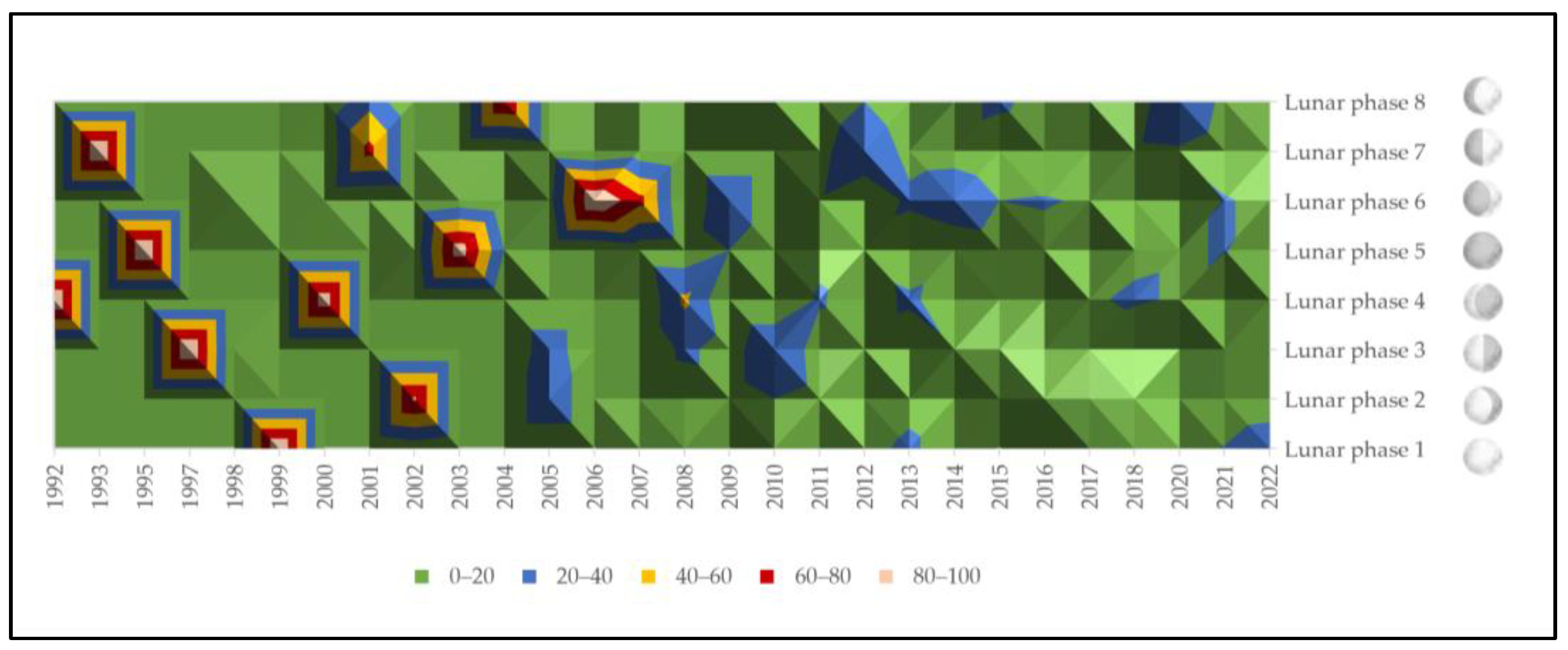
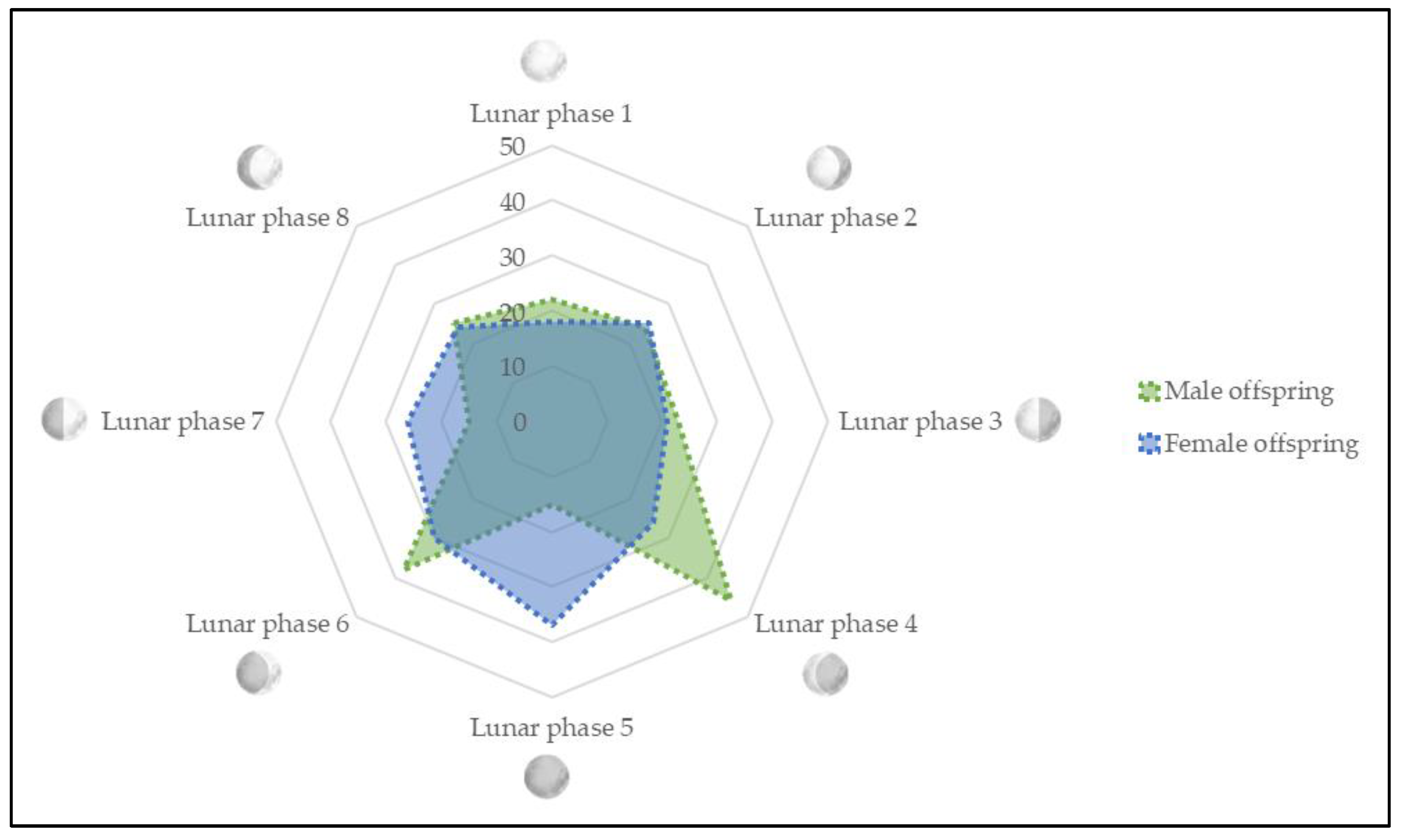
3.1. Quantitative Distribution of Spontaneous Onset of Labor across Lunar Phases
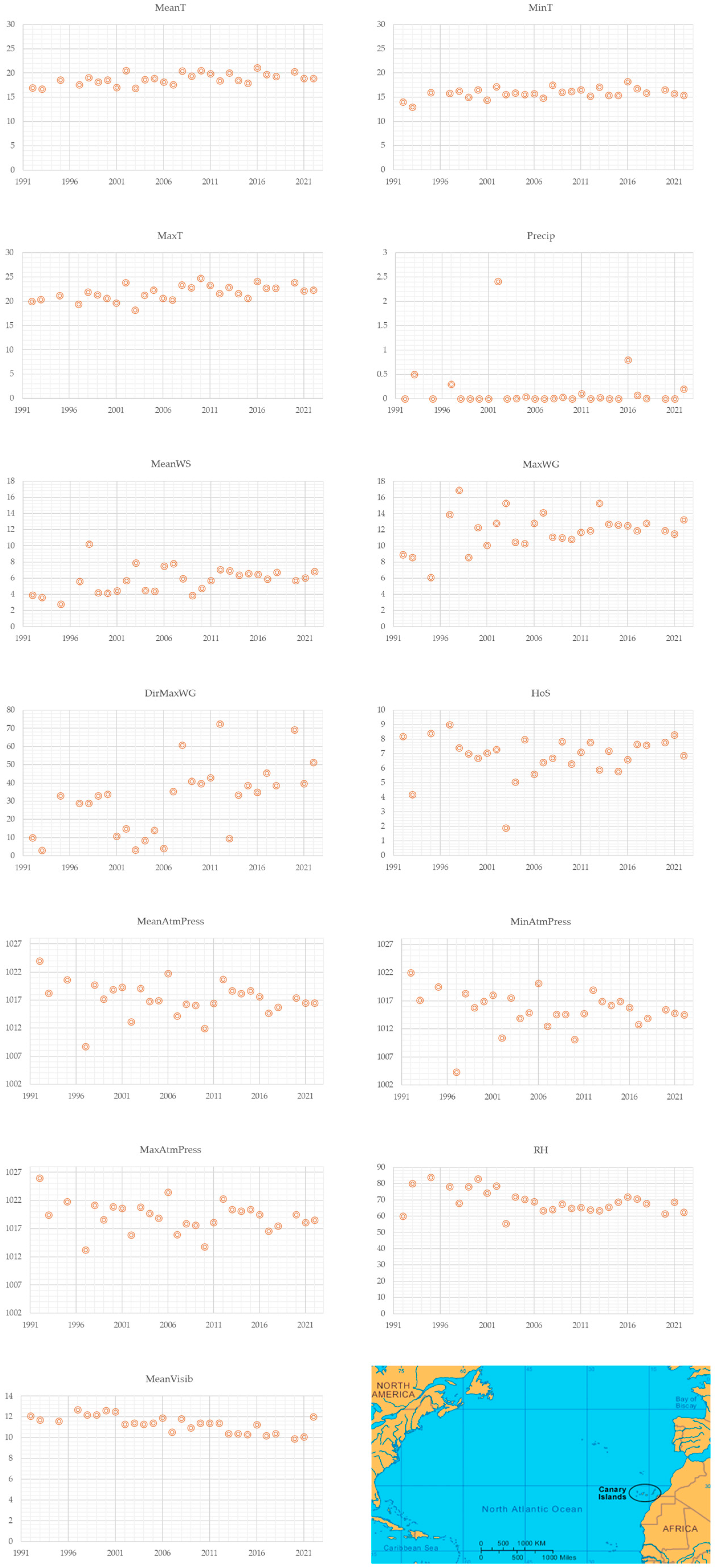
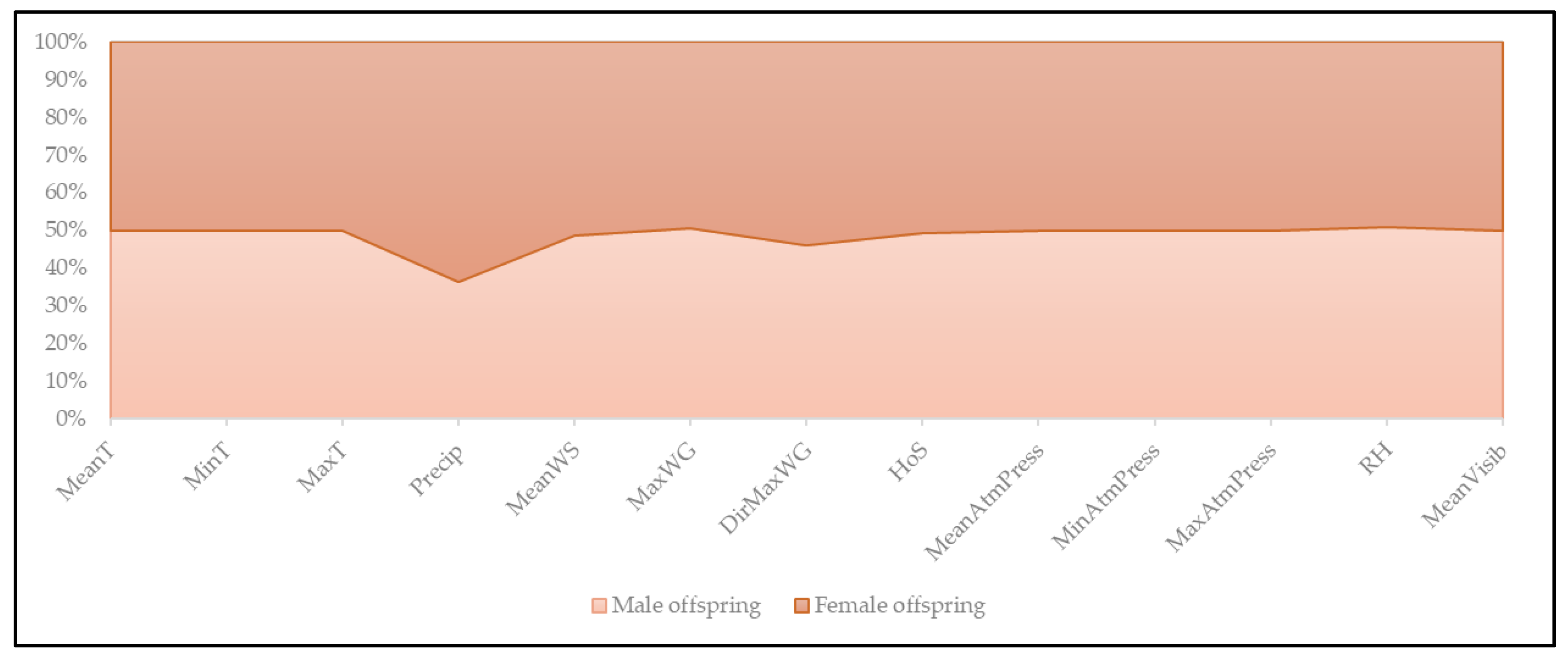
3.2. Mean Climate per Spontaneous Onset of Parturition
3.3. Prediction of Spontaneous Onset of Parturition Based on Environmental Variables
3.3.1. Multicollinearity Analysis
3.3.2. Model Performance
4. Discussion
4.1. Low Moonlight Increases Affiliative Behavior and Territorial Defense
4.2. Dissimilar but Complementary Effects of Air Velocity on Thermal Comfort and Olfactory Maternal Behavior
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Patke, A.; Young, M.W.; Axelrod, S. Molecular mechanisms and physiological importance of circadian rhythms. Nat. Rev. Mol. Cell Biol. 2020, 21, 67–84. [Google Scholar] [CrossRef]
- Vaze, K.M.; Sharma, V.K. On the adaptive significance of circadian clocks for their owners. Chronobiol. Int. 2013, 30, 413–433. [Google Scholar] [CrossRef] [PubMed]
- Benstaali, C.; Mailloux, A.; Bogdan, A.; Auzeby, A.; Touitou, Y. Circadian rhythms of body temperature and motor activity in rodents: Their relationships with the light-dark cycle. Life Sci. 2001, 68, 2645–2656. [Google Scholar] [CrossRef] [PubMed]
- López-Olmeda, J.F.; Madrid, J.A.; Sánchez-Vázquez, F.J. Light and temperature cycles as zeitgebers of zebrafish (Danio rerio) circadian activity rhythms. Chronobiol. Int. 2006, 23, 537–550. [Google Scholar] [CrossRef]
- Prokkola, J.M.; Nikinmaa, M. Circadian rhythms and environmental disturbances–underexplored interactions. J. Exp. Biol. 2018, 221, jeb179267. [Google Scholar] [CrossRef] [PubMed]
- Walton, J.C.; Weil, Z.M.; Nelson, R.J. Influence of photoperiod on hormones, behavior, and immune function. Front. Neuroendocrinol. 2011, 32, 303–319. [Google Scholar] [CrossRef]
- Reppert, S.M. A colorful model of the circadian clock. Cell 2006, 124, 233–236. [Google Scholar] [CrossRef]
- Weil, Z.M.; Bowers, S.L.; Nelson, R.J. Photoperiod alters affective responses in collared lemmings. Behav. Brain Res. 2007, 179, 305–309. [Google Scholar] [CrossRef]
- Workman, J.L.; Bowers, S.L.; Nelson, R.J. Enrichment and photoperiod interact to affect spatial learning and hippocampal dendritic morphology in white-footed mice (Peromyscus leucopus). Eur. J. Neurosci. 2009, 29, 161–170. [Google Scholar] [CrossRef]
- Grant, R.; Halliday, T.; Chadwick, E. Amphibians’ response to the lunar synodic cycle—A review of current knowledge, recommendations, and implications for conservation. Behav. Ecol. 2013, 24, 53–62. [Google Scholar] [CrossRef]
- Kronfeld-Schor, N.; Dominoni, D.; De la Iglesia, H.; Levy, O.; Herzog, E.D.; Dayan, T.; Helfrich-Forster, C. Chronobiology by moonlight. Proc. R. Soc. B Biol. Sci. 2013, 280, 20123088. [Google Scholar] [CrossRef] [PubMed]
- Abecia, J.-A.; Arrébola, F.; Palacios, C. Offspring sex ratio in sheep, cattle, goats and pigs: Influence of season and lunar phase at conception. Biol. Rhythm Res. 2017, 48, 417–424. [Google Scholar] [CrossRef]
- Fernández-Duque, E.; De La Iglesia, H.; Erkert, H.G. Moonstruck primates: Owl monkeys (Aotus) need moonlight for nocturnal activity in their natural environment. PLoS ONE 2010, 5, e12572. [Google Scholar] [CrossRef] [PubMed]
- Palacios, C.; Abecia, J. Does lunar cycle affect lamb production after artificial insemination in sheep? Biol. Rhythm Res. 2014, 45, 869–873. [Google Scholar] [CrossRef]
- Palmer, M.S.; Fieberg, J.; Swanson, A.; Kosmala, M.; Packer, C. A ‘dynamic’landscape of fear: Prey responses to spatiotemporal variations in predation risk across the lunar cycle. Ecol. Lett. 2017, 20, 1364–1373. [Google Scholar] [CrossRef]
- Yonezawa, T.; Uchida, M.; Tomioka, M.; Matsuki, N. Lunar cycle influences spontaneous delivery in cows. PLoS ONE 2016, 11, e0161735. [Google Scholar] [CrossRef]
- Pfeuty, B.; Thommen, Q.; Corellou, F.; Djouani-Tahri, E.B.; Bouget, F.Y.; Lefranc, M. Circadian clocks in changing weather and seasons: Lessons from the picoalga Ostreococcus tauri. BioEssays 2012, 34, 781–790. [Google Scholar] [CrossRef]
- Troein, C.; Locke, J.C.; Turner, M.S.; Millar, A.J. Weather and seasons together demand complex biological clocks. Curr. Biol. 2009, 19, 1961–1964. [Google Scholar] [CrossRef]
- Boutin, S.; Lane, J.E. Climate change and mammals: Evolutionary versus plastic responses. Evol. Appl. 2014, 7, 29–41. [Google Scholar] [CrossRef]
- Chinchilla-Vargas, J.; Kerns, K.; Rothschild, M.F. Lunar and climatic effects on boar ejaculate traits. Anim. Reprod. Sci. 2018, 193, 117–125. [Google Scholar] [CrossRef]
- Linton, D.M.; Macdonald, D.W. Spring weather conditions influence breeding phenology and reproductive success in sympatric bat populations. J. Anim. Ecol. 2018, 87, 1080–1090. [Google Scholar] [CrossRef] [PubMed]
- Beltran, A.J.; Wu, J.; Laurent, O. Associations of meteorology with adverse pregnancy outcomes: A systematic review of preeclampsia, preterm birth and birth weight. Int. J. Environ. Res. Public Health 2014, 11, 91–172. [Google Scholar] [CrossRef] [PubMed]
- Sasaki, Y.; Kitai, N.; Uematsu, M.; Kitahara, G.; Osawa, T. Daily calving frequency and preterm calving is not associated with lunar cycle but preterm calving is associated with weather conditions in Japanese Black cows. PLoS ONE 2019, 14, e0220255. [Google Scholar] [CrossRef] [PubMed]
- Ammann, T.; Hässig, M.; Rüegg, S.; Bleul, U. Effects of meteorological factors and the lunar cycle on onset of parturition in cows. Prev. Vet. Med. 2016, 126, 11–18. [Google Scholar] [CrossRef]
- Noller, K.L.; Resseguie, L.J.; Vossb, V. The effect of changes in atmospheric pressure on the occurrence of the spontaneous onset of labor in term pregnancies. Am. J. Obstet. Gynecol. 1996, 174, 1192–1199. [Google Scholar] [CrossRef]
- Aguilar, J.; Cuervo-Arango, J.; Santa Juliana, L. Lunar cycles at mating do not influence sex ratio at birth in horses. Chronobiol. Int. 2015, 32, 43–47. [Google Scholar] [CrossRef]
- Alberghina, D.; Gioè, M.; Quartuccio, M.; Liotta, L. The influence of lunar cycle at the time of conception on sex offspring distribution in dogs. Chronobiol. Int. 2021, 38, 1517–1521. [Google Scholar] [CrossRef]
- Stern, E.W.; Glazer, G.L.; Sanduleak, N. Influence of the full and new moon on onset of labor and spontaneous rupture of membranes. J. Nurse Midwifery 1988, 33, 57–61. [Google Scholar] [CrossRef]
- Reyes-Lagos, J.J.; Ledesma-Ramírez, C.I.; Pliego-Carrillo, A.C.; Peña-Castillo, M.Á.; Echeverría, J.C.; Becerril-Villanueva, E.; Pavón, L.; Pacheco-López, G. Neuroautonomic activity evidences parturition as a complex and integrated neuro–immune–endocrine process. Ann. N. Y. Acad. Sci. 2019, 1437, 22–30. [Google Scholar] [CrossRef]
- Parraguez, V.H.; Gonzalez-Bulnes, A. Endocrinology of reproductive function and pregnancy at high altitudes. Curr. Opin. Endocr. Metab. Res. 2020, 11, 27–32. [Google Scholar] [CrossRef]
- Pankhurst, N.; King, H. Temperature and salmonid reproduction: Implications for aquaculture. J. Fish Biol. 2010, 76, 69–85. [Google Scholar] [CrossRef] [PubMed]
- Bova, T.L.; Chiavaccini, L.; Cline, G.F.; Hart, C.G.; Matheny, K.; Muth, A.M.; Voelz, B.E.; Kesler, D.; Memili, E. Environmental stressors influencing hormones and systems physiology in cattle. Reprod. Biol. Endocrinol. 2014, 12, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Palagini, L.; Gemignani, A.; Banti, S.; Manconi, M.; Mauri, M.; Riemann, D. Chronic sleep loss during pregnancy as a determinant of stress: Impact on pregnancy outcome. Sleep Med. 2014, 15, 853–859. [Google Scholar] [CrossRef] [PubMed]
- Mendelson, C.R.; Condon, J.C. New insights into the molecular endocrinology of parturition. J. Steroid Biochem. Mol. Biol. 2005, 93, 113–119. [Google Scholar] [CrossRef] [PubMed]
- Alderman, B.W.; Boyko, E.J.; Loy, G.L.; Jones, R.H.; Keane, E.M.; Daling, J.R. Weather and occurrence of eclampsia. Int. J. Epidemiol. 1988, 17, 582–588. [Google Scholar] [CrossRef]
- Kyriacou, C.P.; Peixoto, A.A.; Sandrelli, F.; Costa, R.; Tauber, E. Clines in clock genes: Fine-tuning circadian rhythms to the environment. Trends Genet. 2008, 24, 124–132. [Google Scholar] [CrossRef]
- Heckwolf, M.J.; Morim, T.; Riccioli, F.; Baltazar-Soares, M. Fresh start after rough rides: Understanding patterns of genetic differentiation upon human-mediated translocations. Biol. Invasions 2021, 23, 3625–3639. [Google Scholar] [CrossRef]
- Pastrana, C.I.; González, F.J.N.; Ciani, E.; Ariza, A.G.; Bermejo, J.V.D. A tool for functional selection of leisure camels: Behaviour breeding criteria may ensure long-term sustainability of a European unique breed. Res. Vet. Sci. 2021, 140, 142–152. [Google Scholar] [CrossRef]
- Faye, B. How many large camelids in the world? A synthetic analysis of the world camel demographic changes. Pastoralism 2020, 10, 1–20. [Google Scholar] [CrossRef]
- Record, M. Intercellular communication by exosomes in placenta: A possible role in cell fusion? Placenta 2014, 35, 297–302. [Google Scholar] [CrossRef]
- Dama, M.S. Parasite stress predicts offspring sex ratio. PLoS ONE 2012, 7, e46169. [Google Scholar] [CrossRef]
- Dwyer, C. Behavioural development in the neonatal lamb: Effect of maternal and birth-related factors. Theriogenology 2003, 59, 1027–1050. [Google Scholar] [CrossRef]
- Plush, K.J. Metabolic Maturity and Vigour in Neonatal Lambs, and Subsequent Impacts on Thermoregulation and Survival. Ph.D. Thesis, University of Adelaide, Docklands, Australia, 2014. [Google Scholar]
- Hoter, A.; Rizk, S.; Naim, H.Y. Cellular and molecular adaptation of Arabian camel to heat stress. Front. Genet. 2019, 10, 588. [Google Scholar] [CrossRef] [PubMed]
- Tibary, A.; El Allali, K. Dromedary camel: A model of heat resistant livestock animal. Theriogenology 2020, 154, 203–211. [Google Scholar] [CrossRef]
- Nagy, P.; Juhász, J. Pregnancy and parturition in dromedary camels I. Factors affecting gestation length, calf birth weight and timing of delivery. Theriogenology 2019, 134, 24–33. [Google Scholar] [CrossRef]
- Tokach, M.; Menegat, M.; Gourley, K.; Goodband, R. Nutrient requirements of the modern high-producing lactating sow, with an emphasis on amino acid requirements. Animal 2019, 13, 2967–2977. [Google Scholar] [CrossRef]
- Dioli, M. Observation on dromedary (Camelus dromedarius) welfare and husbandry practices among nomadic pastoralists. Pastoralism 2022, 12, 1–23. [Google Scholar] [CrossRef]
- Burger, P.A.; Ciani, E. Structural and functional genomics in Old World camels—Where do we stand and where to go. Anim. Front. 2022, 12, 30–34. [Google Scholar] [CrossRef]
- Iglesias Pastrana, C.; Navas González, F.J.; Ciani, E.; Barba Capote, C.J.; Delgado Bermejo, J.V. Effect of research impact on emerging camel husbandry, welfare and social-related awareness. Animals 2020, 10, 780. [Google Scholar] [CrossRef] [PubMed]
- Cavalli, G.; Heard, E. Advances in epigenetics link genetics to the environment and disease. Nature 2019, 571, 489–499. [Google Scholar] [CrossRef] [PubMed]
- Khanvilkar, A.; Samant, S.; Ambore, B. Reproduction in camel. Vet. World 2009, 2, 72. [Google Scholar]
- Iglesias Pastrana, C.; Navas González, F.J.; Ciani, E.; Arando Arbulu, A.; Delgado Bermejo, J.V. The Youngest, the Heaviest and/or the Darkest? Selection Potentialities and Determinants of Leadership in Canarian Dromedary Camels. Animals 2021, 11, 2886. [Google Scholar] [CrossRef] [PubMed]
- Schulz, U. El camello en Lanzarote; Aderlan: Las Palmas, Spain, 2008. [Google Scholar]
- Wilson, R.; Gutierrez, C. The one-humped camel in the Canary Islands: History and present status. Tropicultura 2015, 33, 288–298. [Google Scholar]
- Ricci, A.; Racioppi, V.; Iotti, B.; Bertero, A.; Reed, K.; Pascottini, O.; Vincenti, L. Assessment of the temperature cut-off point by a commercial intravaginal device to predict parturition in Piedmontese beef cows. Theriogenology 2018, 113, 27–33. [Google Scholar] [CrossRef]
- Mann, H.B.; Whitney, D.R. On a test of whether one of two random variables is stochastically larger than the other. Ann. Math. Stat. 1947, 18, 50–60. [Google Scholar] [CrossRef]
- Harrell, F.E. Binary logistic regression. In Regression Modeling Strategies; Springer: Berlin/Heidelberg, Germany, 2015. [Google Scholar]
- Midi, H.; Sarkar, S.K.; Rana, S. Collinearity diagnostics of binary logistic regression model. J. Interdiscip. Math. 2010, 13, 253–267. [Google Scholar] [CrossRef]
- Tripepi, G.; Jager, K.; Dekker, F.; Zoccali, C. Linear and logistic regression analysis. Kidney Int. 2008, 73, 806–810. [Google Scholar] [CrossRef]
- Warda, M.; Prince, A.; Kim, H.K.; Khafaga, N.; Scholkamy, T.; Linhardt, R.J.; Jin, H. Proteomics of old world camelid (Camelus dromedarius): Better understanding the interplay between homeostasis and desert environment. J. Adv. Res. 2014, 5, 219–242. [Google Scholar] [CrossRef]
- Mitchell, A.E.; Wolf, B.O.; Martin, T.E. Proximate and evolutionary sources of variation in offspring energy expenditure in songbirds. Glob. Ecol. Biogeogr. 2022, 31, 765–775. [Google Scholar] [CrossRef]
- Rosenberger, A.; Beijer, Å.; Schoenau, E.; Mester, J.; Rittweger, J.; Zange, J. Changes in motor unit activity and respiratory oxygen uptake during 6 weeks of progressive whole-body vibration combined with progressive, high intensity resistance training. J. Musculoskelet. Neuronal Interact. 2019, 19, 159. [Google Scholar] [PubMed]
- Buckles, K.S.; Hungerman, D.M. Season of birth and later outcomes: Old questions, new answers. Rev. Econ. Stat. 2013, 95, 711–724. [Google Scholar] [CrossRef]
- Isen, A.; Rossin-Slater, M.; Walker, R. Relationship between season of birth, temperature exposure, and later life wellbeing. Proc. Natl. Acad. Sci. USA 2017, 114, 13447–13452. [Google Scholar] [CrossRef] [PubMed]
- Shima, J.S.; Osenberg, C.W.; Alonzo, S.H.; Noonburg, E.G.; Mitterwallner, P.; Swearer, S.E. Reproductive phenology across the lunar cycle: Parental decisions, offspring responses, and consequences for reef fish. Ecology 2020, 101, e03086. [Google Scholar] [CrossRef] [PubMed]
- Havenith, G. Interaction of clothing and thermoregulation. Exo. Dermatol. 2002, 1, 221–230. [Google Scholar] [CrossRef]
- Murren, C.J.; Auld, J.R.; Callahan, H.; Ghalambor, C.K.; Handelsman, C.A.; Heskel, M.A.; Kingsolver, J.; Maclean, H.J.; Masel, J.; Maughan, H. Constraints on the evolution of phenotypic plasticity: Limits and costs of phenotype and plasticity. Heredity 2015, 115, 293–301. [Google Scholar] [CrossRef]
- Björklund, M.; Bergek, S. On the relationship between population differentiation and sampling effort: Is more always better? Oikos 2009, 118, 1127–1129. [Google Scholar] [CrossRef]
- Burger, P.A.; Ciani, E.; Faye, B. Old World camels in a modern world–a balancing act between conservation and genetic improvement. Anim. Genet. 2019, 50, 598–612. [Google Scholar] [CrossRef]
- Goswami, A.; Noirault, E.; Coombs, E.J.; Clavel, J.; Fabre, A.-C.; Halliday, T.J.; Churchill, M.; Curtis, A.; Watanabe, A.; Simmons, N.B. Attenuated evolution of mammals through the Cenozoic. Science 2022, 378, 377–383. [Google Scholar] [CrossRef]
- Bender, D.J.; Bayne, E.M.; Brigham, R.M. Lunar condition influences coyote (Canis latrans) howling. Am. Midl. Nat. 1996, 136, 413–417. [Google Scholar] [CrossRef]
- Kurvers, R.H.; Hoelker, F. Bright nights and social interactions: A neglected issue. Behav. Ecol. 2015, 26, 334–339. [Google Scholar] [CrossRef]
- Penteriani, V.; Delgado, M.d.M. Living in the dark does not mean a blind life: Bird and mammal visual communication in dim light. Philos. Trans. R. Soc. B Biol. Sci. 2017, 372, 20160064. [Google Scholar] [CrossRef]
- Pérez-Granados, C.; Schuchmann, K.-L.; Marques, M.I. Addicted to the moon: Vocal output and diel pattern of vocal activity in two Neotropical nightjars is related to moon phase. Ethol. Ecol. Evol. 2022, 34, 66–81. [Google Scholar] [CrossRef]
- York, J.E.; Young, A.J.; Radford, A.N. Singing in the moonlight: Dawn song performance of a diurnal bird varies with lunar phase. Biol. Lett. 2014, 10, 20130970. [Google Scholar] [CrossRef]
- Gutman, R.; Dayan, T.; Levy, O.; Schubert, I.; Kronfeld-Schor, N. The effect of the lunar cycle on fecal cortisol metabolite levels and foraging ecology of nocturnally and diurnally active spiny mice. PLoS ONE 2011, 6, e23446. [Google Scholar] [CrossRef] [PubMed]
- Krief, S.; Cibot, M.; Bortolamiol, S.; Seguya, A.; Krief, J.-M.; Masi, S. Wild chimpanzees on the edge: Nocturnal activities in croplands. PLoS ONE 2014, 9, e109925. [Google Scholar] [CrossRef] [PubMed]
- Penteriani, V.; Kuparinen, A.; del Mar Delgado, M.; Palomares, F.; López-Bao, J.V.; Fedriani, J.M.; Calzada, J.; Moreno, S.; Villafuerte, R.; Campioni, L. Responses of a top and a meso predator and their prey to moon phases. Oecologia 2013, 173, 753–766. [Google Scholar] [CrossRef] [PubMed]
- Sábato, M.A.L.; de Melo, L.F.B.; Magni, E.M.V.; Young, R.J.; Coelho, C.M. A note on the effect of the full moon on the activity of wild maned wolves, Chrysocyon brachyurus. Behav. Process. 2006, 73, 228–230. [Google Scholar] [CrossRef]
- Pyle, P.; Nur, N.; Henderson, R.P.; DeSante, D.F. The effects of weather and lunar cycle on nocturnal migration of landbirds at southeast Farallon Island, California. Condor 1993, 95, 343–361. [Google Scholar] [CrossRef]
- Rodríguez, A.; Chiaradia, A.; Wasiak, P.; Renwick, L.; Dann, P. Waddling on the dark side: Ambient light affects attendance behavior of little penguins. J. Biol. Rhythm. 2016, 31, 194–204. [Google Scholar] [CrossRef]
- Clarke, J.A.; Chopko, J.T.; Mackessy, S.P. The effect of moonlight on activity patterns of adult and juvenile prairie rattlesnakes (Crotalus viridis viridis). J. Herpetol. 1996, 30, 192–197. [Google Scholar] [CrossRef]
- Seligmann, H.; Anderson, S.C.; Autumn, K.; Bouskila, A.; Saf, R.; Tuniyev, B.S.; Werner, Y.L. Analysis of the locomotor activity of a nocturnal desert lizard (Reptilia: Gekkonidae: Teratoscincus scincus) under varying moonlight. Zoology 2007, 110, 104–117. [Google Scholar] [CrossRef]
- Grant, R.A.; Chadwick, E.A.; Halliday, T. The lunar cycle: A cue for amphibian reproductive phenology? Anim. Behav. 2009, 78, 349–357. [Google Scholar] [CrossRef]
- Gebresilassie, A.; Yared, S.; Aklilu, E.; Kirstein, O.D.; Moncaz, A.; Tekie, H.; Balkew, M.; Warburg, A.; Hailu, A.; Gebre-Michael, T. The influence of moonlight and lunar periodicity on the efficacy of CDC light trap in sampling Phlebotomus (Larroussius) orientalis Parrot, 1936 and other Phlebotomus sandflies (Diptera: Psychodidae) in Ethiopia. Parasites Vectors 2015, 8, 1–7. [Google Scholar] [CrossRef]
- Ray, H.; Chakraverty, M. Lunar periodicity in the conjugation of Conchophthirius lamellidens Ghosh. Nature 1934, 134, 663–664. [Google Scholar] [CrossRef]
- Souza, N.A.; Andrade-Coelho, C.A.; Silva, V.C.; Peixoto, A.A.; Rangel, E.F. Moonlight and blood-feeding behaviour of Lutzomyia intermedia and Lutzomyia whitmani (Diptera: Psychodidae: Phlebotominae), vectors of American cutaneous leishmaniasis in Brazil. Mem. Inst. Oswaldo Cruz 2005, 100, 39–42. [Google Scholar] [CrossRef] [PubMed]
- Chakraborty, U. Effects of different phases of the lunar month on living organisms. Biol. Rhythm Res. 2020, 51, 254–282. [Google Scholar] [CrossRef]
- Vignoli, L.; Luiselli, L. Better in the dark: Two Mediterranean amphibians synchronize reproduction with moonlit nights. Web Ecol. 2013, 13, 1–11. [Google Scholar] [CrossRef]
- Aguirre, A.A.; Palomares, R.A.; De Ondiz, A.D.; Soto, E.R.; Perea, M.S.; Hernández-Fonseca, H.J.; Perea, F.P. Lunar cycle influences reproductive performance of crossbred Brahman cows under tropical conditions. J. Biol. Rhythm. 2021, 36, 160–168. [Google Scholar] [CrossRef]
- Griffin, P.C.; Griffin, S.C.; Waroquiers, C.; Mills, L.S. Mortality by moonlight: Predation risk and the snowshoe hare. Behav. Ecol. 2005, 16, 938–944. [Google Scholar] [CrossRef]
- El-Darawany, A.; El-Tarabany, A.; Teama, F. Circalunar Rhythms Induced Changes in the Body Functions and Reproductive Performance in Pregnant Ewes. Arab J. Nucl. Sci. Appl. 2014, 47, 181–188. [Google Scholar]
- Munley, K.M.; Trinidad, J.C.; Deyoe, J.E.; Adaniya, C.H.; Nowakowski, A.M.; Ren, C.C.; Murphy, G.V.; Reinhart, J.M.; Demas, G.E. Melatonin-dependent changes in neurosteroids are associated with increased aggression in a seasonally breeding rodent. J. Neuroendocrinol. 2021, 33, e12940. [Google Scholar] [CrossRef]
- Bevington, M. Lunar biological effects and the magnetosphere. Pathophysiology 2015, 22, 211–222. [Google Scholar] [CrossRef] [PubMed]
- Burch, J.B.; Reif, J.S.; Yost, M.G. Geomagnetic activity and human melatonin metabolite excretion. Neurosci. Lett. 2008, 438, 76–79. [Google Scholar] [CrossRef]
- Bæk, O.; Cilieborg, M.S.; Nguyen, D.N.; Bering, S.B.; Thymann, T.; Sangild, P.T. Sex-specific survival, growth, immunity and organ development in preterm pigs as models for immature newborns. Front. Pediatr. 2021, 9, 626101. [Google Scholar] [CrossRef] [PubMed]
- Zisk, J.L.; Genen, L.H.; Kirkby, S.; Webb, D.; Greenspan, J.; Dysart, K. Do premature female infants really do better than their male counterparts? Am. J. Perinatol. 2011, 28, 241–246. [Google Scholar] [CrossRef] [PubMed]
- Elsmén, E.; Steen, M.; Hellström-Westas, L. Sex and gender differences in newborn infants: Why are boys at increased risk? J. Mens Health 2004, 1, 303–311. [Google Scholar] [CrossRef]
- McGhee, K.E.; Pintor, L.M.; Suhr, E.L.; Bell, A.M. Maternal exposure to predation risk decreases offspring antipredator behaviour and survival in threespined stickleback. Funct. Ecol. 2012, 26, 932–940. [Google Scholar] [CrossRef]
- Hammadi, I.; Chniter, M.; Atigui, M.; Brahmi, M.; Seddik, M.; Salem, W.; Lévy, F.; Nowak, R.; Hammadi, M. Dam parity and calf sex affect maternal and neonatal behaviors during the first week postpartum in stabled Maghrebi dairy camels. Animal 2021, 15, 100149. [Google Scholar] [CrossRef]
- Brandlová, K.; Bartoš, L.; Haberová, T. Camel calves as opportunistic milk thefts? The first description of allosuckling in domestic bactrian camel (Camelus bactrianus). PLoS ONE 2013, 8, e53052. [Google Scholar] [CrossRef]
- Fiialkovskyi, P.; Cassinello, J.; Brandlová, K.; Ceacero, F. Evidence of three distinct lactation stages in nursing gazelles: Implications on maternal behaviour assessment. Appl. Anim. Behav. Sci. 2022, 258, 105814. [Google Scholar] [CrossRef]
- Berube, C.H.; Festa-Bianchet, M.; Jorgenson, J.T. Reproductive costs of sons and daughters in Rocky Mountain bighorn sheep. Behav. Ecol. 1996, 7, 60–68. [Google Scholar] [CrossRef]
- Birgersoon, B.; Tillbom, M.; Ekvall, K. Male-biased investment in fallow deer: An experimental study. Anim. Behav. 1998, 56, 301–307. [Google Scholar] [CrossRef] [PubMed]
- Cassinello, J. Offspring grazing and suckling rates in a sexually dimorphic ungulate with biased maternal investment (Ammotragus lervia). Ethology 2001, 107, 173–182. [Google Scholar] [CrossRef]
- Clutton-Brock, T.H.; Albon, S.; Guinness, F. Parental investment in male and female offspring in polygynous mammals. Nature 1981, 289, 487–489. [Google Scholar] [CrossRef]
- Gloneková, M.; Brandlová, K.; Pluháček, J. Higher maternal care and tolerance in more experienced giraffe mothers. Acta Ethol. 2020, 23, 1–7. [Google Scholar] [CrossRef]
- Hejcmanová, P.; Vymyslická, P.; Koláčková, K.; Antonínová, M.; Havlíková, B.; Stejskalová, M.; Policht, R.; Hejcman, M. Suckling behavior of eland antelopes (Taurotragus spp.) under semi-captive and farm conditions. J. Ethol. 2011, 29, 161–168. [Google Scholar] [CrossRef]
- Fitak, R.R.; Mohandesan, E.; Corander, J.; Yadamsuren, A.; Chuluunbat, B.; Abdelhadi, O.; Raziq, A.; Nagy, P.; Walzer, C.; Faye, B. Genomic signatures of domestication in Old World camels. Commun. Biol. 2020, 3, 1–10. [Google Scholar] [CrossRef]
- Rancel Rodriguez, N.M. Biodiversity of Epiphyllous, Heterocyst-Forming Cyanobacteria in the Laurel Forest of the Canary Islands. Ph.D. Thesis, Universität zu Köln, Köln, German, 2016. [Google Scholar]
- Azorin-Molina, C.; Menendez, M.; McVicar, T.R.; Acevedo, A.; Vicente-Serrano, S.M.; Cuevas, E.; Minola, L.; Chen, D. Wind speed variability over the Canary Islands, 1948–2014: Focusing on trend differences at the land–ocean interface and below–above the trade-wind inversion layer. Clim. Dyn. 2018, 50, 4061–4081. [Google Scholar] [CrossRef]
- Moyes, K.; Nussey, D.H.; Clements, M.N.; Guinness, F.E.; Morris, A.; Morris, S.; Pemberton, J.M.; Kruuk, L.E.; CLUTTON-BROCK, T.H. Advancing breeding phenology in response to environmental change in a wild red deer population. Glob. Chang. Biol. 2011, 17, 2455–2469. [Google Scholar] [CrossRef]
- Sims, M.; Elston, D.A.; Larkham, A.; Nussey, D.H.; Albon, S.D. Identifying when weather influences life-history traits of grazing herbivores. J. Anim. Ecol. 2007, 76, 761–770. [Google Scholar] [CrossRef]
- Frederiksen, M.; Harris, M.P.; Daunt, F.; Rothery, P.; Wanless, S. Scale-dependent climate signals drive breeding phenology of three seabird species. Glob. Chang. Biol. 2004, 10, 1214–1221. [Google Scholar] [CrossRef]
- Virens, E.; Cree, A. Wind of change: A diurnal skink thermoregulates between cooler set-points and for an increased amount of time in the presence of wind. J. Exp. Biol. 2022, 225, jeb244038. [Google Scholar] [CrossRef] [PubMed]
- Veselý, M.; Zeiler, W. Personalized conditioning and its impact on thermal comfort and energy performance–A review. Renew. Sust. Energ. Rev. 2014, 34, 401–408. [Google Scholar] [CrossRef]
- Foster, J.; Smallcombe, J.W.; Hodder, S.; Jay, O.; Flouris, A.D.; Havenith, G. Quantifying the impact of heat on human physical work capacity; part II: The observed interaction of air velocity with temperature, humidity, sweat rate, and clothing is not captured by most heat stress indices. Int. J. Biometeorol. 2022, 66, 507–520. [Google Scholar] [CrossRef] [PubMed]
- Roche, J.; Lee, J.; Berry, D. Climatic factors and secondary sex ratio in dairy cows. J. Dairy Sci. 2006, 89, 3221–3227. [Google Scholar] [CrossRef]
- Togunov, R.R.; Derocher, A.E.; Lunn, N.J. Windscapes and olfactory foraging in a large carnivore. Sci. Rep. 2017, 7, 46332. [Google Scholar] [CrossRef]
- Stěhulová, I.; Špinka, M.; Šárová, R.; Máchová, L.; Kněz, R.; Firla, P. Maternal behaviour in beef cows is individually consistent and sensitive to cow body condition, calf sex and weight. Appl. Anim. Behav. Sci. 2013, 144, 89–97. [Google Scholar] [CrossRef]
- Lévy, F.; Keller, M.; Poindron, P. Olfactory regulation of maternal behavior in mammals. Horm. Behav. 2004, 46, 284–302. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variable | Tolerance (1 − R2) | VIF (1/Tolerance) |
|---|---|---|
| Lunar phase | 0.931 | 1.074 |
| Mean temperature | 0.000 | 4575.800 |
| Minimum temperature | 0.001 | 1132.490 |
| Maximum temperature | 0.001 | 1697.666 |
| Precipitation | 0.879 | 1.138 |
| Mean wind speed | 0.289 | 3.463 |
| Maximum wind gust | 0.314 | 3.182 |
| Direction of the maximum wind gust | 0.846 | 1.183 |
| Hours of sunshine | 0.691 | 1.448 |
| Mean atmospheric pressure | 0.060 | 16.239 |
| Minimum atmospheric pressure | 0.059 | 16.820 |
| Maximum atmospheric pressure | 0.063 | 15.838 |
| Relative humidity | 0.731 | 1.368 |
| Mean visibility | 0.888 | 1.126 |
| Omnibus Test | Nagelkerke R2 | HL Goodness-of-Fit Test | Prediction Accuracy Rate (%) | Variable | Coefficient (B) | Standard Error of B | Wald Statistic | Statistical Significance | ESTIMATED Odds Ratio (exp(B)) |
|---|---|---|---|---|---|---|---|---|---|
| 0.000 | 0.125 | 0.025 | 59.3 | Lunar phase 8 (reference category) | 17.860 | 0.013 | 1 | ||
| Lunar phase 1 | −0.404 | 0.451 | 0.802 | 0.370 | 0.668 | ||||
| Lunar phase 2 | 0.093 | 0.425 | 0.048 | 0.827 | 1.097 | ||||
| Lunar phase 3 | 0.013 | 0.434 | 0.001 | 0.977 | 1.013 | ||||
| Lunar phase 4 | −0.526 | 0.405 | 1.685 | 0.194 | 0.591 | ||||
| Lunar phase 5 | 1.024 | 0.439 | 5.430 | 0.020 | 2.783 | ||||
| Lunar phase 6 | −0.277 | 0.396 | 0.329 | 0.566 | 0.797 | ||||
| Lunar phase 7 | 0.394 | 0.455 | 0.749 | 0.387 | 1.482 | ||||
| Precipitation | 0.156 | 0.106 | 2.149 | 0.143 | 1.168 | ||||
| Mean wind speed | 0.265 | 0.075 | 12.574 | 0.000 | 1.303 | ||||
| Maximum wind gust | −0.177 | 0.056 | 10.123 | 0.001 | 0.838 | ||||
| Direction of the maximum wind gust | 0.002 | 0.003 | 0.454 | 0.500 | 1.002 | ||||
| Hours of sunshine | 0.051 | 0.043 | 1.371 | 0.242 | 1.052 | ||||
| Relative humidity | −0.012 | 0.011 | 1.212 | 0.271 | 0.988 | ||||
| Mean visibility | 0.145 | 0.104 | 1.959 | 0.162 | 1.156 | ||||
| Constant | −0.654 | 1.627 | 0.162 | 0.687 | 0.520 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Iglesias Pastrana, C.; Navas González, F.J.; Delgado Bermejo, J.V.; Ciani, E. Lunar Cycle, Climate, and Onset of Parturition in Domestic Dromedary Camels: Implications of Species-Specific Metabolic Economy and Social Ecology. Biology 2023, 12, 607. https://doi.org/10.3390/biology12040607
Iglesias Pastrana C, Navas González FJ, Delgado Bermejo JV, Ciani E. Lunar Cycle, Climate, and Onset of Parturition in Domestic Dromedary Camels: Implications of Species-Specific Metabolic Economy and Social Ecology. Biology. 2023; 12(4):607. https://doi.org/10.3390/biology12040607
Chicago/Turabian StyleIglesias Pastrana, Carlos, Francisco Javier Navas González, Juan Vicente Delgado Bermejo, and Elena Ciani. 2023. "Lunar Cycle, Climate, and Onset of Parturition in Domestic Dromedary Camels: Implications of Species-Specific Metabolic Economy and Social Ecology" Biology 12, no. 4: 607. https://doi.org/10.3390/biology12040607
APA StyleIglesias Pastrana, C., Navas González, F. J., Delgado Bermejo, J. V., & Ciani, E. (2023). Lunar Cycle, Climate, and Onset of Parturition in Domestic Dromedary Camels: Implications of Species-Specific Metabolic Economy and Social Ecology. Biology, 12(4), 607. https://doi.org/10.3390/biology12040607