
Climate Change, Extreme Temperatures and Sex-Related Responses in Spiders

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
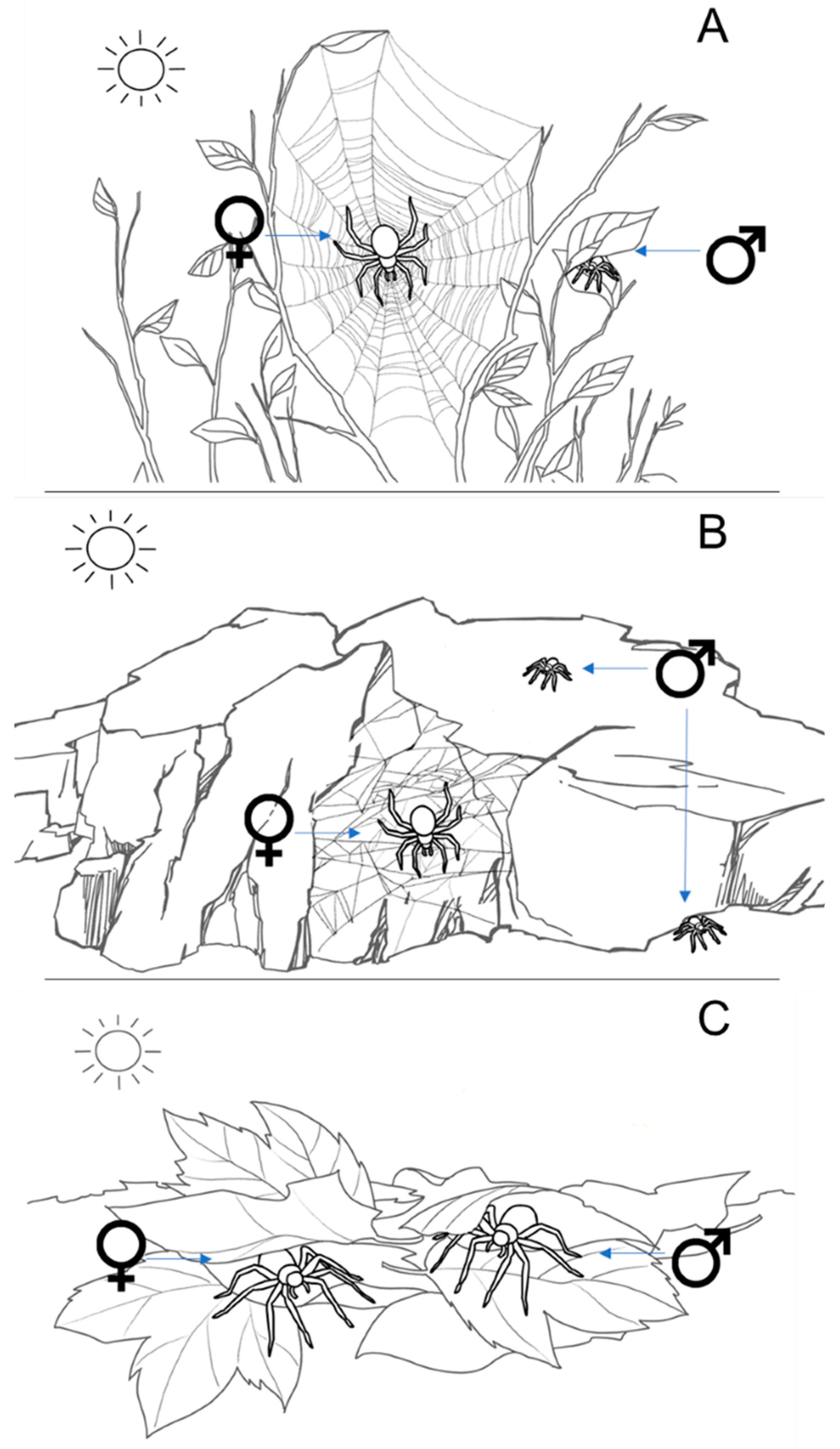
2. Spiders and Climate Change
2.1. Sexual Size-Dimorphism and Ecological Variation in Male and Female Spiders
2.2. Physiological and Behavioral Responses of Spiders to High Temperatures
3. Conclusions and Future Directions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dirzo, R.; Young, H.S.; Galetti, M.; Ceballos, G.; Isaac, N.J.B.; Collen, B. Defaunation in the Anthropocene. Science 2014, 345, 401–406. [Google Scholar] [CrossRef]
- Pievani, T. The sixth mass extinction: Anthropocene and the human impact on biodiversity. Rend. Lincei 2014, 25, 85–93. [Google Scholar] [CrossRef]
- Ceballos, G.; Ehrlich, P.R.; Dirzo, R. Biological annihilation via the ongoing sixth mass extinction signaled by vertebrate population losses and declines. Proc. Natl. Acad. Sci. USA 2017, 114, E6089–E6096. [Google Scholar] [CrossRef]
- Lande, R. Anthropogenic, ecological and genetic factors in extinction and conservation. Popul. Ecol. 1998, 40, 259–269. [Google Scholar] [CrossRef]
- Ripple, W.J.; Wolf, C.; Newsome, T.M.; Hoffmann, M.; Wirsing, A.J.; McCauley, D.J. Extinction risk is most acute for the world’s largest and smallest vertebrates. Proc. Natl. Acad. Sci. USA 2017, 114, 10678–10683. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Bayo, F.; Wyckhuys, K.A.G. Worldwide decline of the entomofauna: A review of its drivers. Biol. Conserv. 2019, 232, 8–27. [Google Scholar] [CrossRef]
- van Klink, R.; Bowler, D.E.; Gongalsky, K.B.; Swengel, A.B.; Gentile, A.; Chase, J.M. Meta-analysis reveals declines in terrestrial but increases in freshwater insect abundances. Science 2020, 368, 417–420. [Google Scholar] [CrossRef] [PubMed]
- Wagner, D.L. Insect Declines in the Anthropocene. Annu. Rev. Entomol. 2020, 65, 457–480. [Google Scholar] [CrossRef] [PubMed]
- Wagner, D.L.; Grames, E.M.; Forister, M.L.; Berenbaum, M.R.; Stopak, D. Insect decline in the Anthropocene: Death by a thousand cuts. Proc. Natl. Acad. Sci. USA 2021, 118, e2023989118. [Google Scholar] [CrossRef] [PubMed]
- Lister, B.C.; Garcia, A. Climate-driven declines in arthropod abundance restructure a rainforest food web. Proc. Natl. Acad. Sci. USA 2018, 115, E10397–E10406. [Google Scholar] [CrossRef]
- Goulson, D. Pesticides linked to bird declines. Nature 2014, 511, 295–296. [Google Scholar] [CrossRef]
- Rosenberg, K.V.; Dokter, A.M.; Blancher, P.J.; Sauer, J.R.; Smith, A.C.; Smith, P.A.; Stanton, J.C.; Panjabi, A.; Helft, L.; Parr, M.; et al. Decline of the North American avifauna. Science 2019, 366, 120–124. [Google Scholar] [CrossRef]
- Cramer, W.; Bondeau, A.; Woodward, F.I.; Prentice, I.C.; Betts, R.A.; Brovkin, V.; Cox, P.M.; Fisher, V.; Foley, J.A.; Friend, A.D.; et al. Global response of terrestrial ecosystem structure and function to CO2 and climate change: Results from six dynamic global vegetation models. Glob. Chang. Biol. 2001, 7, 357–373. [Google Scholar] [CrossRef]
- Wardle, D.A.; Bardgett, R.D.; Callaway, R.M.; Van der Putten, W.H. Terrestrial ecosystem responses to species gains and losses. Science 2011, 332, 1273–1277. [Google Scholar] [CrossRef]
- Pelini, S.L.; Maran, A.M.; Chen, A.R.; Kaseman, J.; Crowther, T.W. Higher trophic levels overwhelm climate change impacts on terrestrial ecosystem functioning. PLoS ONE 2015, 10, e0136344. [Google Scholar] [CrossRef]
- Noriega, J.A.; Hortal, J.; Azcárate, F.M.; Berg, M.P.; Bonada, N.; Briones, M.J.I.; Del Toro, I.; Goulson, D.; Ibanez, S.; Landis, D.A.; et al. Research trends in ecosystem services provided by insects. Basic Appl. Ecol. 2018, 26, 8–23. [Google Scholar] [CrossRef]
- Dangles, O.; Casas, J. Ecosystem services provided by insects for achieving sustainable development goals. Ecosyst. Serv. 2019, 35, 109–115. [Google Scholar] [CrossRef]
- Cardoso, P.; Barton, P.S.; Birkhofer, K.; Chichorro, F.; Deacon, C.; Fartmann, T.; Fukushima, C.S.; Gaigher, R.; Habel, J.C.; Hallmann, C.A.; et al. Scientists’ warning to humanity on insect extinctions. Biol. Conserv. 2020, 242, 108426. [Google Scholar] [CrossRef]
- Harvey, J.A.; Tougeron, K.; Gols, R.; Heinen, R.; Abarca, M.; Abram, P.K.; Basset, Y.; Berg, M.; Boggs, C.; Brodeur, J.; et al. Scientists’ warning on climate change and insects. Ecol. Monogr. 2023, 93, e1553. [Google Scholar] [CrossRef]
- Walther, G.-R.; Post, E.; Convey, P.; Menzel, A.; Parmesan, C.; Beebee, T.J.C.; Fromentin, J.-M.; Hoegh-Guldberg, O.; Bairlein, F. Ecological responses to recent climate change. Nature 2002, 416, 389–395. [Google Scholar] [CrossRef] [PubMed]
- Bellard, C.; Bertelsmeier, C.; Leadley, P.; Thuiller, W.; Courchamp, F. Impacts of climate change on the future of biodiversity. Ecol. Lett. 2012, 15, 365–377. [Google Scholar] [CrossRef]
- Urban, M.C.; Bocedi, G.; Hendry, A.P.; Mihoub, J.-B.; Pe’er, G.; Singer, A.; Bridle, J.R.; Crozier, L.G.; De Meester, L.; Godsoe, W.; et al. Improving the forecast for biodiversity under climate change. Science 2016, 353, aad8466. [Google Scholar] [CrossRef]
- Joachimski, M.M.; Lai, X.; Shen, S.; Jiang, H.; Luo, G.; Chen, B.; Chen, J.; Sun, Y. Climate warming in the latest Permian and the Permian–Triassic mass extinction. Geology 2012, 40, 195–198. [Google Scholar] [CrossRef]
- Lewis, S.L.; Maslin, M.A. Defining the Anthropocene. Nature 2015, 519, 171–180. [Google Scholar] [CrossRef]
- Brook, B.W.; Sodhi, N.S.; Bradshaw, C.J.A. Synergies among extinction drivers under global change. Trends Ecol. Evol. 2008, 23, 453–460. [Google Scholar] [CrossRef]
- Thomas, C.D.; Cameron, A.; Green, R.E.; Bakkenes, M.; Beaumont, L.J.; Collingham, Y.C.; Erasmus, B.F.N.; de Siqueira, M.F.; Grainger, A.; Hannah, L.; et al. Extinction risk from climate change. Nature 2004, 427, 145–148. [Google Scholar] [CrossRef] [PubMed]
- Deutsch, C.A.; Tewksbury, J.J.; Huey, R.B.; Sheldon, K.S.; Ghalambor, C.K.; Haak, D.C.; Martin, P.R. Impacts of climate warming on terrestrial ectotherms across latitude. Proc. Natl. Acad. Sci. USA 2008, 105, 6668–6672. [Google Scholar] [CrossRef]
- Hoffmann, A.A.; Chown, S.L.; Clusella-Trullas, S. Upper thermal limits in terrestrial ectotherms: How constrained are they? Funct. Ecol. 2013, 27, 934–949. [Google Scholar] [CrossRef]
- Kingsolver, J.G.; Diamond, S.E.; Buckley, L.B. Heat stress and the fitness consequences of climate change for terrestrial ectotherms. Funct. Ecol. 2013, 27, 1415–1423. [Google Scholar] [CrossRef]
- Paaijmans, K.P.; Heinig, R.L.; Seliga, R.A.; Blanford, J.I.; Blanford, S.; Murdock, C.C.; Thomas, M.B. Temperature variation makes ectotherms more sensitive to climate change. Glob. Chang. Biol. 2013, 19, 2373–2380. [Google Scholar] [CrossRef]
- Colinet, H.; Sinclair, B.J.; Vernon, P.; Renault, D. Insects in Fluctuating Thermal Environments. Annu. Rev. Entomol. 2015, 60, 123–140. [Google Scholar] [CrossRef] [PubMed]
- Cloudsley-Thompson, J.L. Adaptations of Arthropoda to Arid Environments. Annu. Rev. Entomol. 1975, 20, 261–283. [Google Scholar] [CrossRef]
- Bale, J.S. Insect cold hardiness: Freezing and supercooling—An ecophysiological perspective. J. Insect Physiol. 1987, 33, 899–908. [Google Scholar] [CrossRef]
- Willot, Q.; Gueydan, C.; Aron, S. Proteome stability, heat hardening and heat-shock protein expression profiles in Cataglyphis desert ants. J. Exp. Biol. 2017, 220, 1721–1728. [Google Scholar] [CrossRef]
- Halsch, C.A.; Shapiro, A.M.; Fordyce, J.A.; Nice, C.C.; Thorne, J.H.; Waetjen, D.P.; Forister, M.L. Insects and recent climate change. Proc. Natl. Acad. Sci. USA 2021, 118, e2002543117. [Google Scholar] [CrossRef] [PubMed]
- Chapman, J.W.; Reynolds, D.R.; Wilson, K. Long-range seasonal migration in insects: Mechanisms, evolutionary drivers and ecological consequences. Ecol. Lett. 2015, 18, 287–302. [Google Scholar] [CrossRef] [PubMed]
- Bale, J.S.; Hayward, S.A.L. Insect overwintering in a changing climate. J. Exp. Biol. 2010, 213, 980–994. [Google Scholar] [CrossRef]
- Badeck, F.-W.; Bondeau, A.; Böttcher, K.; Doktor, D.; Lucht, W.; Schaber, J.; Sitch, S. Responses of spring phenology to climate change. New Phytol. 2004, 162, 295–309. [Google Scholar] [CrossRef]
- Thuiller, W. Patterns and uncertainties of species’ range shifts under climate change. Glob. Chang. Biol. 2004, 10, 2020–2027. [Google Scholar] [CrossRef]
- Parmesan, C. Ecological and Evolutionary Responses to Recent Climate Change. Annu. Rev. Ecol. Evol. Syst. 2006, 37, 637–669. [Google Scholar] [CrossRef]
- Kelly, A.E.; Goulden, M.L. Rapid shifts in plant distribution with recent climate change. Proc. Natl. Acad. Sci. USA 2008, 105, 11823–11826. [Google Scholar] [CrossRef] [PubMed]
- Tobin, P.C.; Nagarkatti, S.; Loeb, G.; Saunders, M.C. Historical and projected interactions between climate change and insect voltinism in a multivoltine species. Glob. Chang. Biol. 2008, 14, 951–957. [Google Scholar] [CrossRef]
- Chen, S.; Fleischer, S.J.; Tobin, P.C.; Saunders, M.C. Projecting insect voltinism under high and low greenhouse gas emission conditions. Environ. Entomol. 2011, 40, 505–515. [Google Scholar] [CrossRef]
- Meehl, G.A.; Tebaldi, C. More intense, more frequent, and longer lasting heat waves in the 21st century. Science 2004, 305, 994–997. [Google Scholar] [CrossRef] [PubMed]
- Rahmstorf, S.; Coumou, D. Increase of extreme events in a warming world. Proc. Natl. Acad. Sci. USA 2011, 108, 17905–17909. [Google Scholar] [CrossRef]
- Palmer, T. Climate forecasting: Build high-resolution global climate models. Nature 2014, 515, 338–339. [Google Scholar] [CrossRef]
- Mazdiyasni, O.; AghaKouchak, A. Substantial increase in concurrent droughts and heatwaves in the United States. Proc. Natl. Acad. Sci. USA 2015, 112, 11484–11489. [Google Scholar] [CrossRef]
- Christidis, N.; Jones, G.S.; Stott, P.A. Dramatically increasing chance of extremely hot summers since the 2003 European heatwave. Nat. Clim. Chang. 2015, 5, 46–50. [Google Scholar] [CrossRef]
- Christidis, N.; Stott, P.A.; Zwiers, F.W. Fast-track attribution assessments based on pre-computed estimates of changes in the odds of warm extremes. Clim. Dyn. 2015, 45, 1547–1564. [Google Scholar] [CrossRef]
- Reusch, T.B.H.; Ehlers, A.; Hämmerli, A.; Worm, B. Ecosystem recovery after climatic extremes enhanced by genotypic diversity. Proc. Natl. Acad. Sci. USA 2005, 102, 2826–2831. [Google Scholar] [CrossRef] [PubMed]
- Isbell, F.; Craven, D.; Connolly, J.; Loreau, M.; Schmid, B.; Beierkuhnlein, C.; Bezemer, T.M.; Bonin, C.; Bruelheide, H.; de Luca, E.; et al. Biodiversity increases the resistance of ecosystem productivity to climate extremes. Nature 2015, 526, 574–577. [Google Scholar] [CrossRef]
- Harris, R.M.B.; Beaumont, L.J.; Vance, T.R.; Tozer, C.R.; Remenyi, T.A.; Perkins-Kirkpatrick, S.E.; Mitchell, P.J.; Nicotra, A.B.; McGregor, S.; Andrew, N.R.; et al. Biological responses to the press and pulse of climate trends and extreme events. Nat. Clim. Chang. 2018, 8, 579–587. [Google Scholar] [CrossRef]
- Neven, L.G. Physiological responses of insects to heat. Postharvest Biol. Technol. 2000, 21, 103–111. [Google Scholar] [CrossRef]
- González-Tokman, D.; Córdoba-Aguilar, A.; Dáttilo, W.; Lira-Noriega, A.; Sánchez-Guillén, R.A.; Villalobos, F. Insect responses to heat: Physiological mechanisms, evolution and ecological implications in a warming world. Biol. Rev. 2020, 95, 802–821. [Google Scholar] [CrossRef]
- Sales, K.; Vasudeva, R.; Dickinson, M.E.; Godwin, J.L.; Lumley, A.J.; Michalczyk, Ł.; Hebberecht, L.; Thomas, P.; Franco, A.; Gage, M.J.G. Experimental heatwaves compromise sperm function and cause transgenerational damage in a model insect. Nat. Commun. 2018, 9, 4771. [Google Scholar] [CrossRef] [PubMed]
- Parratt, S.R.; Walsh, B.S.; Metelmann, S.; White, N.; Manser, A.; Bretman, A.J.; Hoffmann, A.A.; Snook, R.R.; Price, T.A.R. Temperatures that sterilize males better match global species distributions than lethal temperatures. Nat. Clim. Chang. 2021, 11, 481–484. [Google Scholar] [CrossRef]
- Iossa, G. Sex-specific differences in thermal fertility limits. Trends Ecol. Evol. 2019, 34, 490–492. [Google Scholar] [CrossRef]
- Walsh, B.S.; Parratt, S.R.; Hoffmann, A.A.; Atkinson, D.; Snook, R.R.; Bretman, A.; Price, T.A. The impact of climate change on fertility. Trends Ecol. Evol. 2019, 34, 249–259. [Google Scholar] [CrossRef] [PubMed]
- van Heerwaarden, B.; Sgrò, C.M. Male fertility thermal limits predict vulnerability to climate warming. Nat. Commun. 2021, 12, 2214. [Google Scholar] [CrossRef]
- Walsh, B.S.; Mannion, N.L.M.; Price, T.A.R.; Parratt, S.R. Sex-specific sterility caused by extreme temperatures is likely to create cryptic changes to the operational sex ratio in Drosophila virilis. Curr. Zool. 2021, 67, 341–343. [Google Scholar] [CrossRef]
- Thakur, M.P.; Risch, A.C.; van der Putten, W.H. Biotic responses to climate extremes in terrestrial ecosystems. iScience 2022, 25, 104559. [Google Scholar] [CrossRef] [PubMed]
- Agosta, S.J.; Joshi, K.A.; Kester, K.M. Upper thermal limits differ among and within component species in a tritrophic host-parasitoid-hyperparasitoid system. PLoS ONE 2018, 13, e0198803. [Google Scholar] [CrossRef]
- Riechert, S.E.; Lockley, T. Spiders as biological control agents. Annu. Rev. Entomol. 1984, 29, 299–320. [Google Scholar] [CrossRef]
- Nyffeler, M.; Birkhofer, K. An estimated 400–800 million tons of prey are annually killed by the global spider community. Sci. Nat. 2017, 104, 30. [Google Scholar] [CrossRef]
- Kumschick, S.; Fronzek, S.; Entling, M.H.; Nentwig, W. Rapid spread of the wasp spider Argiope bruennichi across Europe: A consequence of climate change? Clim. Chang. 2011, 109, 319–329. [Google Scholar] [CrossRef]
- Saupe, E.E.; Papes, M.; Selden, P.A.; Vetter, R.S. Tracking a medically important spider: Climate change, ecological niche modeling, and the brown recluse (Loxosceles reclusa). PLoS ONE 2011, 6, e17731. [Google Scholar] [CrossRef]
- Ferretti, N.E.; Arnedo, M.; González, A. Impact of climate change on spider species distribution along theLa Plata River Basin, Southern South America: Projecting future range shifts for the genus Stenoterommata (Araneae, Mygalomorphae, Nemesiidae). Ann. Zool. Fenn. 2018, 55, 123–133. [Google Scholar] [CrossRef]
- Moradmand, M.; Yousefi, M. Ecological niche modelling and climate change in two species groups of huntsman spider genus Eusparassus in the Western Palearctic. Sci. Rep. 2022, 12, 4138. [Google Scholar] [CrossRef]
- Mammola, S.; Goodacre, S.L.; Isaia, M. Climate change may drive cave spiders to extinction. Ecography 2018, 41, 233–243. [Google Scholar] [CrossRef]
- Cardoso, P.; Leather, S.R. Predicting a global insect apocalypse. Insect Conserv. Divers. 2019, 12, 263–267. [Google Scholar] [CrossRef]
- Goulson, D. The insect apocalypse, and why it matters. Curr. Biol. 2019, 29, R967–R971. [Google Scholar] [CrossRef] [PubMed]
- van der Sluijs, J.P. Insect decline, an emerging global environmental risk. Curr. Opin. Environ. Sustain. 2020, 46, 39–42. [Google Scholar] [CrossRef]
- Foelix, R.F. Biology of Spiders; Harvard University Press: Cambridge, MA, USA, 1982. [Google Scholar]
- Head, G. Selection on fecundity and variation in the degree of sexual size dimorphism among spider species (Class Araneae). Evolution 1995, 49, 776–781. [Google Scholar] [CrossRef] [PubMed]
- Kuntner, M.; Coddington, J.A. Sexual size dimorphism: Evolution and perils of extreme phenotypes in spiders. Annu. Rev. Entomol. 2020, 65, 57–80. [Google Scholar] [CrossRef]
- Vollrath, F.; Parker, G.A. Sexual dimorphism and distorted sex ratios in spiders. Nature 1992, 360, 156–159. [Google Scholar] [CrossRef]
- Hormiga, G.; Scharff, N.; Coddington, J.A. The Phylogenetic Basis of Sexual Size Dimorphism in Orb-Weaving Spiders (Araneae, Orbiculariae). Syst. Biol. 2000, 49, 435–462. [Google Scholar] [CrossRef] [PubMed]
- Quiñones-Lebrón, S.G.; Gregorič, M.; Kuntner, M.; Kralj-Fišer, S. Small size does not confer male agility advantages in a sexually-size dimorphic spider. PLoS ONE 2019, 14, e0216036. [Google Scholar] [CrossRef]
- Yu, K.-P.; Kuntner, M.; Cheng, R.-C.; Austin, A.; Yu, K.-P.; Kuntner, M.; Cheng, R.-C.; Austin, A. Phylogenetic evidence for an independent origin of extreme sexual size dimorphism in a genus of araneid spiders (Araneae: Araneidae). Invertebr. Syst. 2022, 36, 48–62. [Google Scholar] [CrossRef]
- Prenter, J.; Elwood, R.W.; Montgomery, W.I. No association between sexual size dimorphism and life histories in spiders. Proc. R. Soc. Lond. B Biol. Sci. 1998, 265, 57–62. [Google Scholar] [CrossRef]
- Suzuki, Y. Sit-and-wait forager or aggressive spider predator: Araneophagic cases of orb-weaving spiders by crab spiders of the genus Thomisus (Araneae: Thomisidae). Invertebr. Zool. 2020, 17, 189–194. [Google Scholar] [CrossRef]
- Walker, S.E.; Marshall, S.D.; Rypstra, A.L.; Taylor, D.H. The effects of hunger on locomotory behaviour in two species of wolf spider (Araneae, Lycosidae). Anim. Behav. 1999, 58, 515–520. [Google Scholar] [CrossRef]
- Willemart, R.H.; Lacava, M. Foraging strategies of cursorial and ambush spiders. In Behaviour and Ecology of Spiders: Contributions from the Neotropical Region; Viera, C., Gonzaga, M.O., Eds.; Springer International Publishing: Cham, Switzerland, 2017; pp. 227–245. ISBN 978-3-319-65717-2. [Google Scholar]
- Moya-Laraño, J.; Halaj, J.; Wise, D.H. Climbing to reach females: Romeo should be small. Evolution 2002, 56, 420–425. [Google Scholar] [CrossRef]
- Brandt, Y.; Andrade, M.C.B. Testing the gravity hypothesis of sexual size dimorphism: Are small males faster climbers? Funct. Ecol. 2007, 21, 379–385. [Google Scholar] [CrossRef]
- Moya-Laraño, J.; Vinkovi&cacute, D.; Allard, C.M.; Foellmer, M.W. Mass-mediated sex differences in climbing patterns support the gravity hypothesis of sexual size dimorphism. Web Ecol. 2007, 7, 106–112. [Google Scholar] [CrossRef]
- Corcobado, G.; Rodríguez-Gironés, M.A.; De Mas, E.; Moya-Laraño, J. Introducing the refined gravity hypothesis of extreme sexual size dimorphism. BMC Evol. Biol. 2010, 10, 236. [Google Scholar] [CrossRef]
- Grossi, B.; Canals, M. Energetics, scaling and sexual size dimorphism of spiders. Acta Biotheor. 2015, 63, 71–81. [Google Scholar] [CrossRef] [PubMed]
- Schmitt, A.; Schuster, M.; Barth, F.G. Daily locomotor activity patterns in three species of Cupiennius (Araneae, Ctenidae): The males are the wandering spiders. J. Arachnol. 1990, 18, 249–255. [Google Scholar]
- Henschel, J.R. Long-distance wandering and mating by the dancing white lady spider (Leucorchestris arenicola) (Araneae, Sparassidae) across namib dunes. J. Arachnol. 2002, 30, 321–330. [Google Scholar] [CrossRef]
- Stellwag, L.M.; Dodson, G.N. Navigation by male crab spiders Misumenoides formosipes (Araneae: Thomisidae): Floral cues may aid in locating potential mates. J. Insect Behav. 2010, 23, 226–235. [Google Scholar] [CrossRef]
- Mas, E.D.; Ribera, C.; Moya-Laraño, J. Resurrecting the differential mortality model of sexual size dimorphism. J. Evol. Biol. 2009, 22, 1739–1749. [Google Scholar] [CrossRef]
- Harvey, J.A. Prey availability affects developmental trade-offs and sexual-size dimorphism in the false widow spider, Steatoda grossa. J. Insect Physiol. 2022, 136, 104267. [Google Scholar] [CrossRef]
- Aisenberg, A.; Costa, F.G.; González, M.; Postiglioni, R.; Pérez-Miles, F. Sexual dimorphism in chelicerae, forelegs and palpal traits in two burrowing wolf spiders (Araneae: Lycosidae) with sex-role reversal. J. Nat. Hist. 2010, 44, 1189–1202. [Google Scholar] [CrossRef]
- Gasnier, T.R.; Azevedo, C.S.D.; Torres-Sanchez, M.P.; Höfer, H. Adult size of eight hunting spider species in central Amazonia: Temporal variations and sexual dimorphisms. J. Arachnol. 2002, 30, 146–154. [Google Scholar] [CrossRef]
- Humphreys, W.F. Respiration studies on Geolycosa godeffroyi (Araneae:Lycosidae) and their relationship to field estimates of metabolic heat loss. Comp. Biochem. Physiol. A Physiol. 1977, 57, 255–263. [Google Scholar] [CrossRef]
- Brandt, E.E.; Roberts, K.T.; Williams, C.M.; Elias, D.O. Low temperatures impact species distributions of jumping spiders across a desert elevational cline. J. Insect Physiol. 2020, 122, 104037. [Google Scholar] [CrossRef] [PubMed]
- Meineke, E.K.; Holmquist, A.J.; Wimp, G.M.; Frank, S.D. Changes in spider community composition are associated with urban temperature, not herbivore abundance. J. Urban Ecol. 2017, 3, juw010. [Google Scholar] [CrossRef]
- Starrett, J.; Waters, E.R. Positive natural selection has driven the evolution of the Hsp70s in Diguetia spiders. Biol. Lett. 2007, 3, 439–444. [Google Scholar] [CrossRef]
- Chen, L.; Li, Z.; Zhou, X.; Xing, X.; Lv, B. Integrated transcriptome and metabolome analysis reveals molecular responses of spider to single and combined high temperature and drought stress. Environ. Pollut. 2023, 317, 120763. [Google Scholar] [CrossRef]
- Høye, T.T.; Hammel, J.U.; Fuchs, T.; Toft, S. Climate change and sexual size dimorphism in an Arctic spider. Biol. Lett. 2009, 5, 542–544. [Google Scholar] [CrossRef]
- Brown, A.; Houghton, D. Effect of laboratory heat stress on mortality and web mass of the common house spider, Parasteatoda tepidariorum (Koch 1841) (Araneae: Theridiidae). Gt. Lakes Entomol. 2020, 52, 8. [Google Scholar]
- Johnson, J.C.; Urcuyo, J.; Moen, C.; Ii, D.R.S. Urban heat island conditions experienced by the Western black widow spider (Latrodectus hesperus): Extreme heat slows development but results in behavioral accommodations. PLoS ONE 2019, 14, e0220153. [Google Scholar] [CrossRef] [PubMed]
- Johnson, J.C.; Garver, E.; Martin, T. Black widows on an urban heat island: Extreme heat affects spider development and behaviour from egg to adulthood. Anim. Behav. 2020, 167, 77–84. [Google Scholar] [CrossRef]
- Hayasaka, D.; Nakamori, T.; Tamaue, K.; Seko, Y.; Hashimoto, K.; Sawahata, T. Dry-heat tolerance of egg sacs of invasive Latrodectus spiders (Araneae: Theridiidae) in Japan: Implications for efficient control/extermination. J. Econ. Entomol. 2021, 114, 2460–2465. [Google Scholar] [CrossRef] [PubMed]
- Cramer, K.L.; Zagar, L.M. Heat tolerance of the brown recluse spider (Araneae: Sicariidae): Potential for pest control. J. Econ. Entomol. 2016, 109, 284–287. [Google Scholar] [CrossRef] [PubMed]
- Pruitt, J.N.; Riechert, S.E.; Jones, T.C. Behavioural syndromes and their fitness consequences in a socially polymorphic spider, Anelosimus studiosus. Anim. Behav. 2008, 76, 871–879. [Google Scholar] [CrossRef]
- de Tranaltes, C.; Dunn, J.; Martin, J.M.; Johnson, J.C. Siblicide in the city: The urban heat island accelerates sibling cannibalism in the black widow spider (Latrodectus hesperus). Urban Ecosyst. 2022, 25, 305–312. [Google Scholar] [CrossRef]
- Brandt, E.E.; Rosenthal, M.F.; Elias, D.O. Complex interactions between temperature, sexual signals and mate choice in a desert-dwelling jumping spider. Anim. Behav. 2020, 170, 81–87. [Google Scholar] [CrossRef]
- Entling, W.; Schmidt-Entling, M.H.; Bacher, S.; Brandl, R.; Nentwig, W. Body size–climate relationships of European spiders. J. Biogeogr. 2010, 37, 477–485. [Google Scholar] [CrossRef]
- Correa-Garhwal, S.M.; Chaw, R.C.; Clarke, T.H.; Ayoub, N.A.; Hayashi, C.Y. Silk gene expression of theridiid spiders: Implications for male-specific silk use. Zoology 2017, 122, 107–114. [Google Scholar] [CrossRef]
- Craig, C.L.; Bernard, G.D.; Coddington, J.A. Evolutionary shifts in the spectral properties of spider silks. Evolution 1994, 48, 287–296. [Google Scholar] [CrossRef]
- Kato, N.; Takasago, M.; Omasa, K.; Miyashita, T. Coadaptive changes in physiological and biophysical traits related to thermal stress in web spiders. Naturwissenschaften 2008, 95, 1149–1153. [Google Scholar] [CrossRef] [PubMed]
- Riechert, S.E.; Tracy, C.R. Thermal balance and prey availability: Bases for a model relating web-site characteristics to spider reproductive success. Ecology 1975, 56, 265–284. [Google Scholar] [CrossRef]
- Tanaka, K. Food consumption and diet composition of the web-building spider Agelena limbata in two habitats. Oecologia 1991, 86, 8–15. [Google Scholar] [CrossRef]
- Blamires, S.J.; Sellers, W.I. Modelling temperature and humidity effects on web performance: Implications for predicting orb-web spider (Argiope spp.) foraging under Australian climate change scenarios. Conserv. Physiol. 2019, 7, coz083. [Google Scholar] [CrossRef] [PubMed]
- Leather, S.R. Size, reproductive potential and fecundity in insects: Things aren’t as simple as they seem. Oikos 1988, 51, 386–389. [Google Scholar] [CrossRef]
- Omkar, M.G. (Ed.) Reproductive Strategies in Insects; CRC Press: Boca Raton, FL, USA, 2022; ISBN 978-1-00-304319-5. [Google Scholar]
- Michalik, P.; Ramírez, M.J. Evolutionary morphology of the male reproductive system, spermatozoa and seminal fluid of spiders (Araneae, Arachnida)—Current knowledge and future directions. Arthropod Struct. Dev. 2014, 43, 291–322. [Google Scholar] [CrossRef]
- Suter, R.B.; Parkhill, V.S. Fitness consequences of prolonged copulation in the bowl and doily spider. Behav. Ecol. Sociobiol. 1990, 26, 369–373. [Google Scholar] [CrossRef]
- Linn, C.D.; Molina, Y.; Difatta, J.; Christenson, T.E. The adaptive advantage of prolonged mating: A test of alternative hypotheses. Anim. Behav. 2007, 74, 481–485. [Google Scholar] [CrossRef]
- Schneider, J.M.; Gilberg, S.; Fromhage, L.; Uhl, G. Sexual conflict over copulation duration in a cannibalistic spider. Anim. Behav. 2006, 71, 781–788. [Google Scholar] [CrossRef]
- Bukowski, T.C.; Christenson, T.E. Determinants of sperm release and storage in a spiny orbweaving spider. Anim. Behav. 1997, 53, 381–395. [Google Scholar] [CrossRef]
- Peñuelas, J.; Sardans, J.; Estiarte, M.; Ogaya, R.; Carnicer, J.; Coll, M.; Barbeta, A.; Rivas-Ubach, A.; Llusià, J.; Garbulsky, M.; et al. Evidence of current impact of climate change on life: A walk from genes to the biosphere. Glob. Chang. Biol. 2013, 19, 2303–2338. [Google Scholar] [CrossRef]
- Harvey, J.A.; Heinen, R.; Gols, R.; Thakur, M.P. Climate change-mediated temperature extremes and insects: From outbreaks to breakdowns. Glob. Chang. Biol. 2020, 26, 6685–6701. [Google Scholar] [CrossRef]
- Pincebourde, S.; Woods, H.A. There is plenty of room at the bottom: Microclimates drive insect vulnerability to climate change. Curr. Opin. Insect Sci. 2020, 41, 63–70. [Google Scholar] [CrossRef]
- Ma, C.-S.; Ma, G.; Pincebourde, S. Survive a Warming Climate: Insect Responses to Extreme High Temperatures. Annu. Rev. Entomol. 2021, 66, 163–184. [Google Scholar] [CrossRef]
- Robinson, S.L.; Lundholm, J.T. Ecosystem services provided by urban spontaneous vegetation. Urban Ecosyst. 2012, 15, 545–557. [Google Scholar] [CrossRef]
- Gols, R.; Ojeda-Prieto, L.M.; Li, K.; van der Putten, W.H.; Harvey, J.A. Within-patch and edge microclimates vary over a growing season and are amplified during a heatwave: Consequences for ectothermic insects. J. Therm. Biol. 2021, 99, 103006. [Google Scholar] [CrossRef]
- Klockmann, M.; Kleinschmidt, F.; Fischer, K. Carried over: Heat stress in the egg stage reduces subsequent performance in a butterfly. PLoS ONE 2017, 12, e0180968. [Google Scholar] [CrossRef] [PubMed]
- Michalko, R.; Pekár, S.; Dul’a, M.; Entling, M.H. Global patterns in the biocontrol efficacy of spiders: A meta-analysis. Glob. Ecol. Biogeogr. 2019, 28, 1366–1378. [Google Scholar] [CrossRef]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Harvey, J.A.; Dong, Y. Climate Change, Extreme Temperatures and Sex-Related Responses in Spiders. Biology 2023, 12, 615. https://doi.org/10.3390/biology12040615
Harvey JA, Dong Y. Climate Change, Extreme Temperatures and Sex-Related Responses in Spiders. Biology. 2023; 12(4):615. https://doi.org/10.3390/biology12040615
Chicago/Turabian StyleHarvey, Jeffrey A., and Yuting Dong. 2023. "Climate Change, Extreme Temperatures and Sex-Related Responses in Spiders" Biology 12, no. 4: 615. https://doi.org/10.3390/biology12040615
APA StyleHarvey, J. A., & Dong, Y. (2023). Climate Change, Extreme Temperatures and Sex-Related Responses in Spiders. Biology, 12(4), 615. https://doi.org/10.3390/biology12040615