

Whole-Transcriptome Analysis of Repeated Low-Level Sarin-Exposed Rat Hippocampus and Identification of Cerna Networks to Investigate the Mechanism of Sarin-Induced Cognitive Impairment
 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Drugs
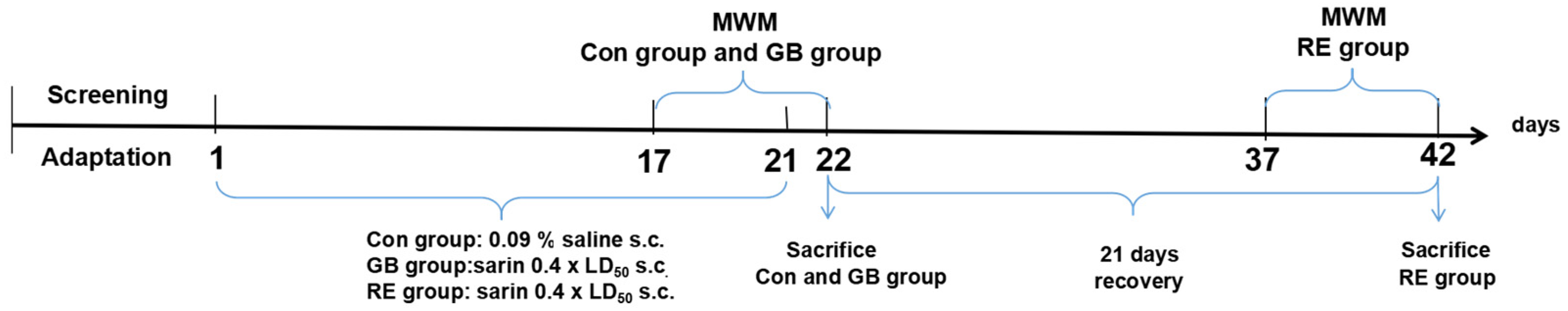
2.3. Administration and Assignment
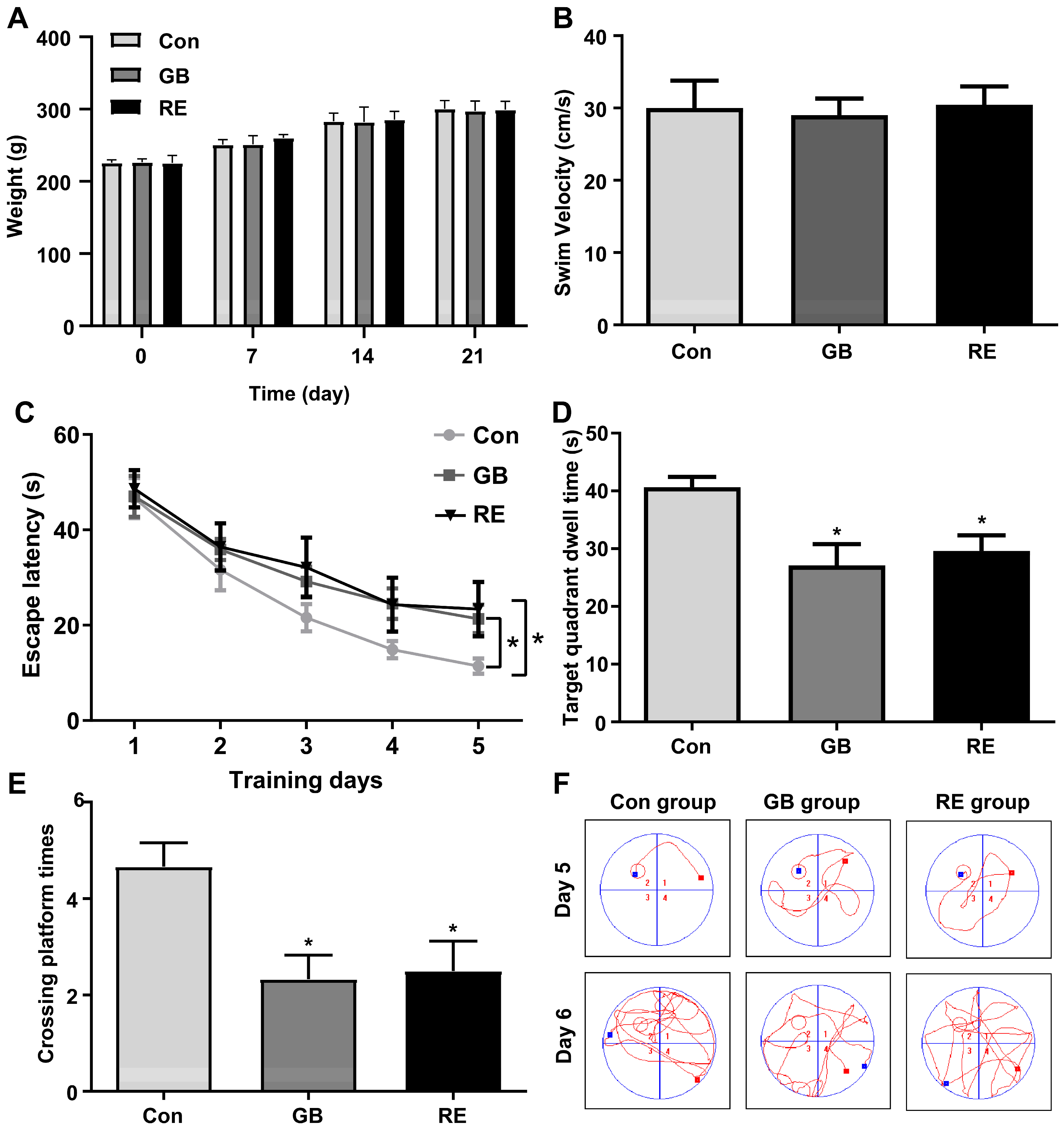
2.4. Morris Water Maze Test
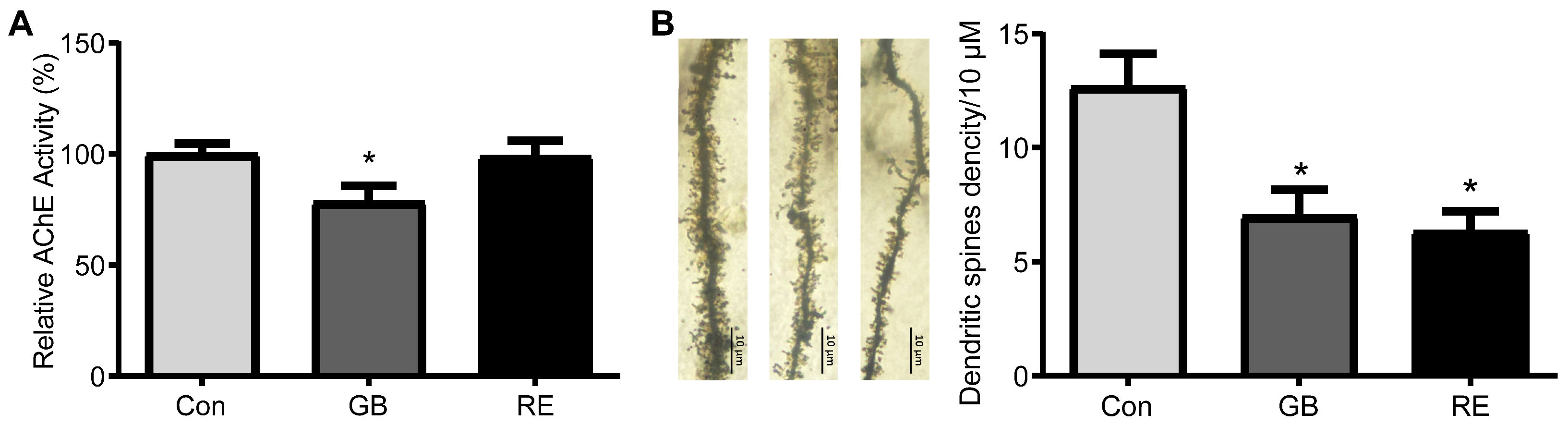
2.5. AChE Activity Assay
2.6. Golgi Staining and Counting
2.7. RNA Isolation, Library Construction, and Sequencing
2.8. Gene Expression Level and Differential Expression Analysis
2.9. Gene Ontology Annotations, Kyoto Encyclopedia of Genes and Genomes Pathway Analysis, and Protein–Protein Interaction Network of DEGs
2.10. ceRNAs Regulatory Network Analysis
2.11. Statistical Analyses
3. Results
3.1. Sarin Irreversibly Impaired the Learning and Memory Functions of Rats
3.2. Effects of Sarin on AChE Activity and Dendritic Spines Density in Rat Hippocampus
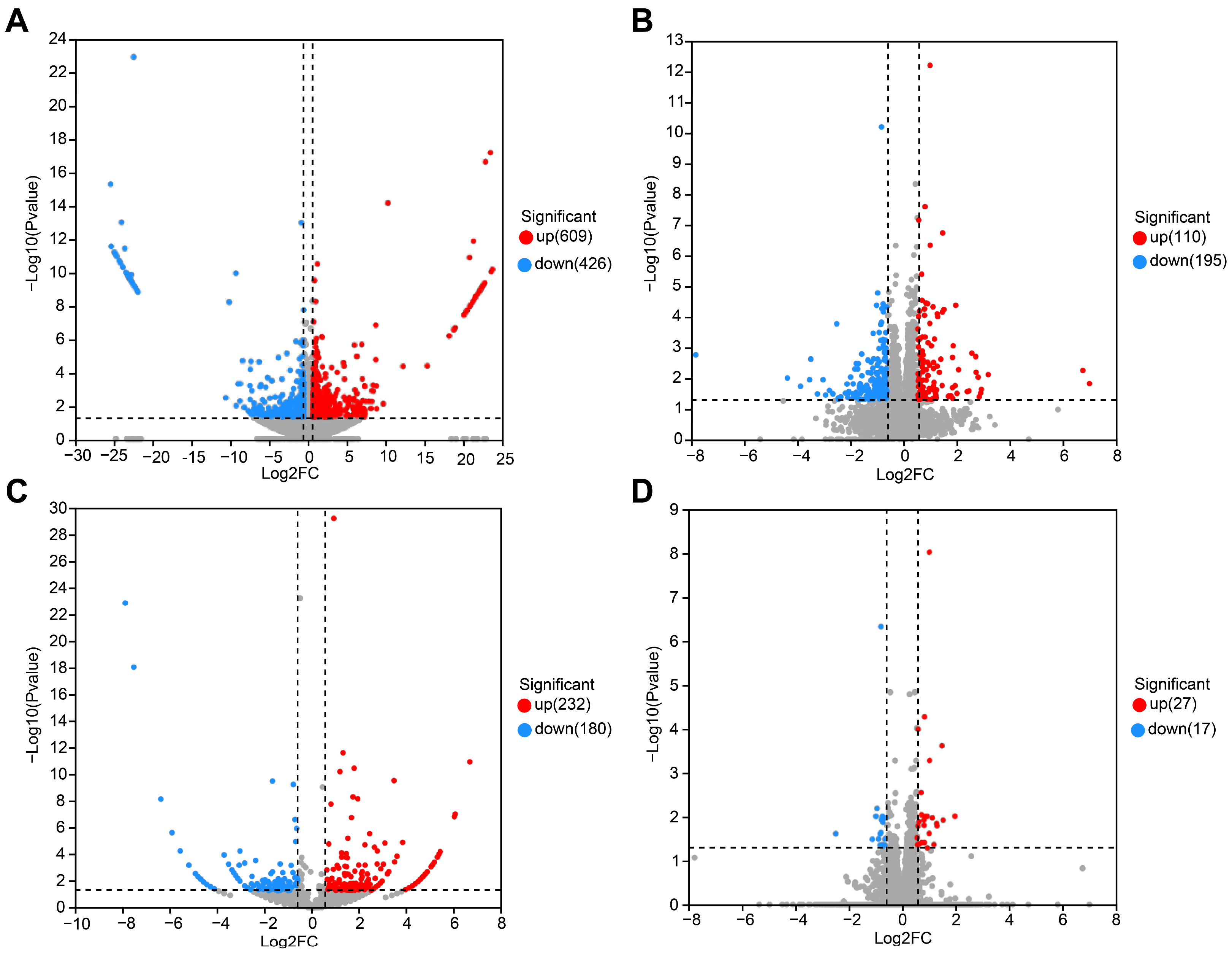
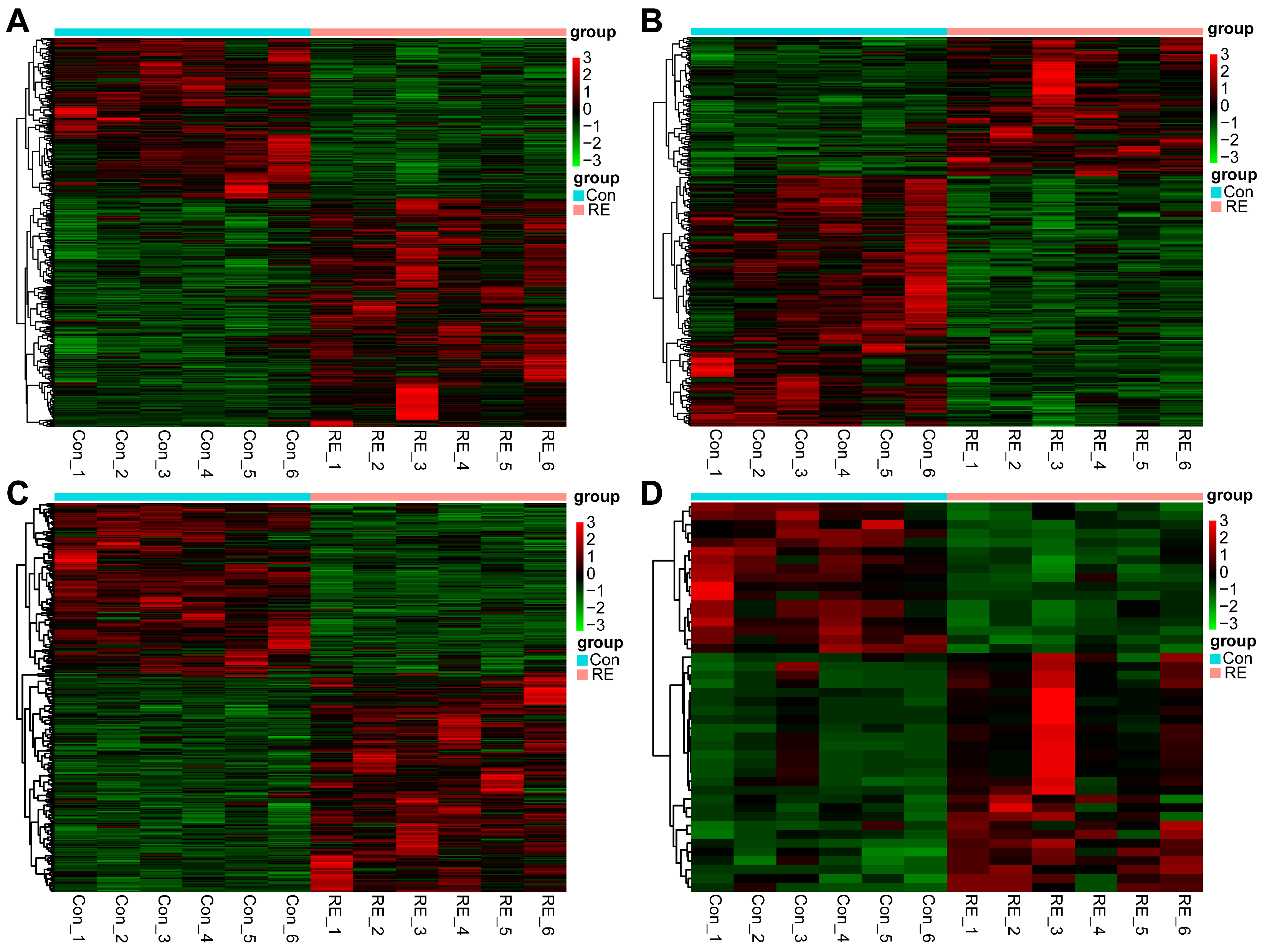
3.3. Differential Expression Analysis
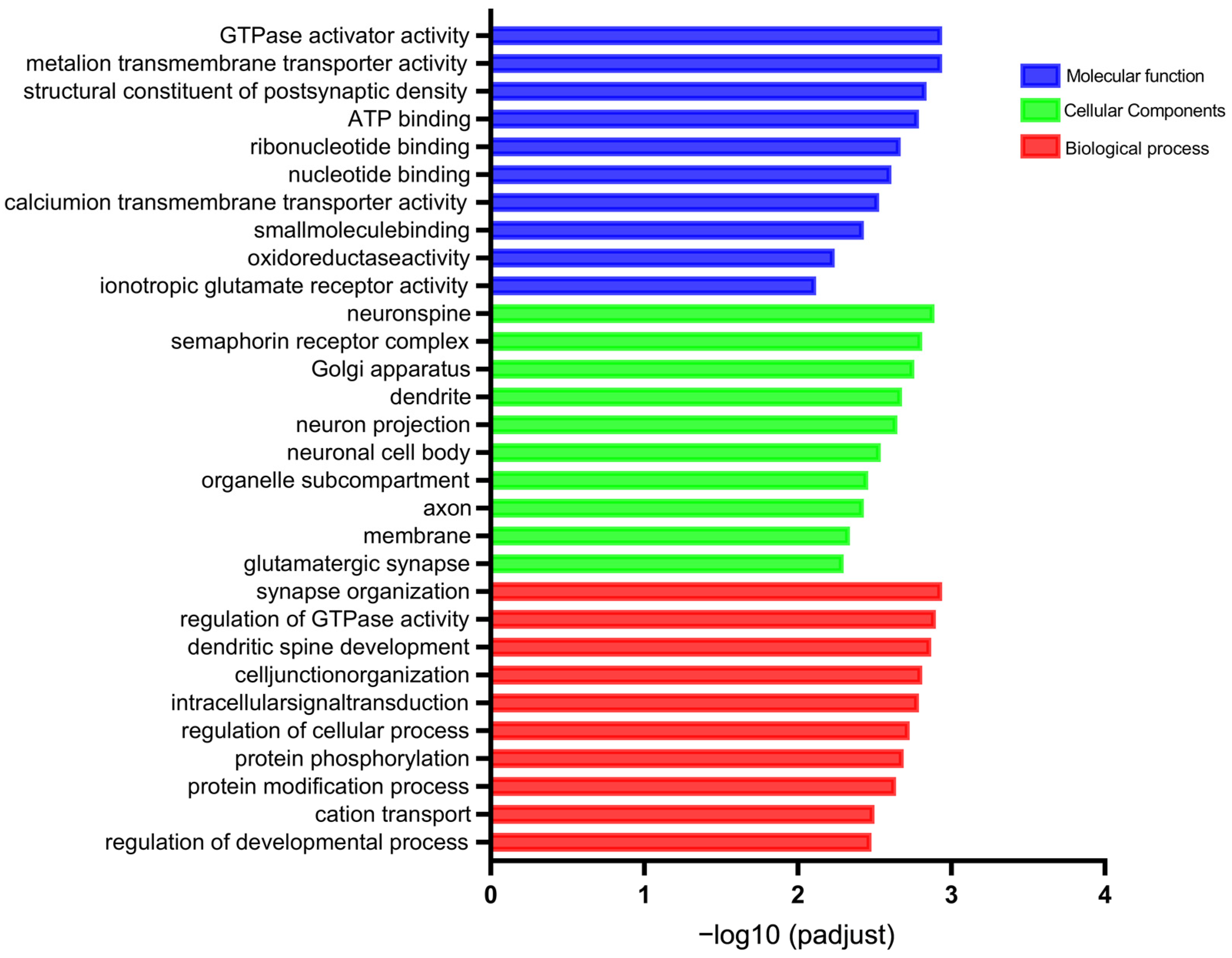
3.4. GO Annotation and KEGG Pathway Analysis
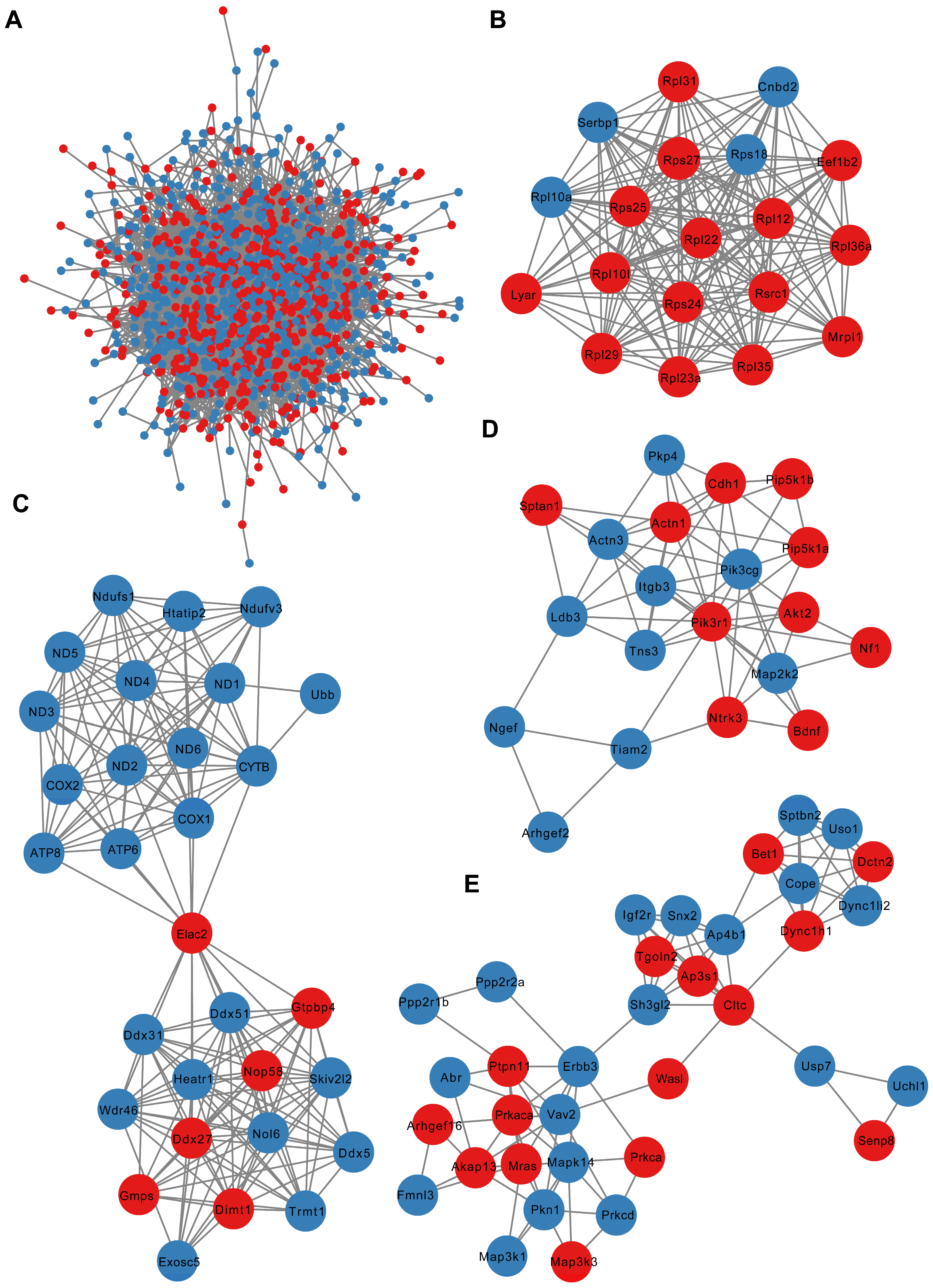
3.5. PPI Network Analysis
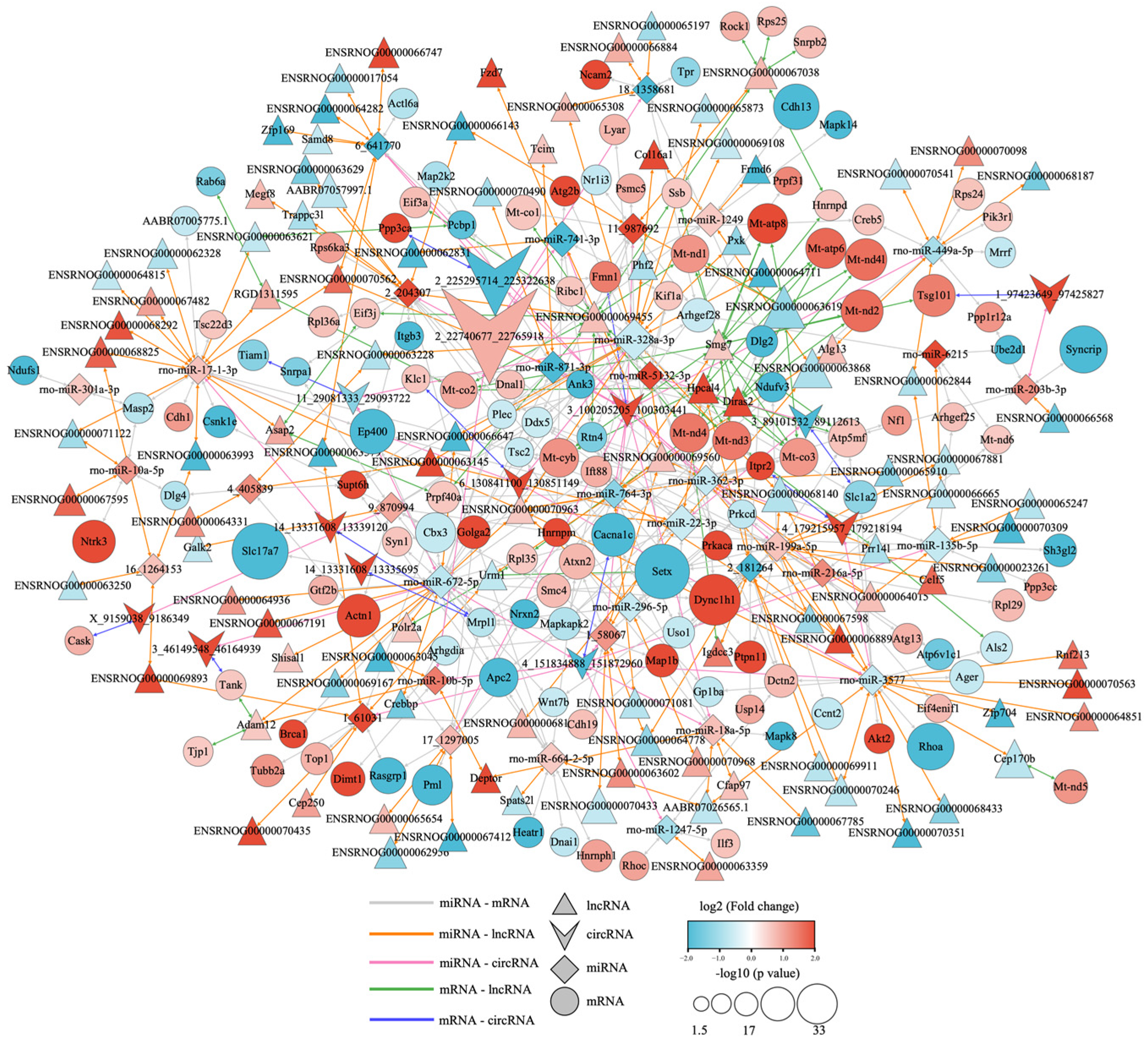
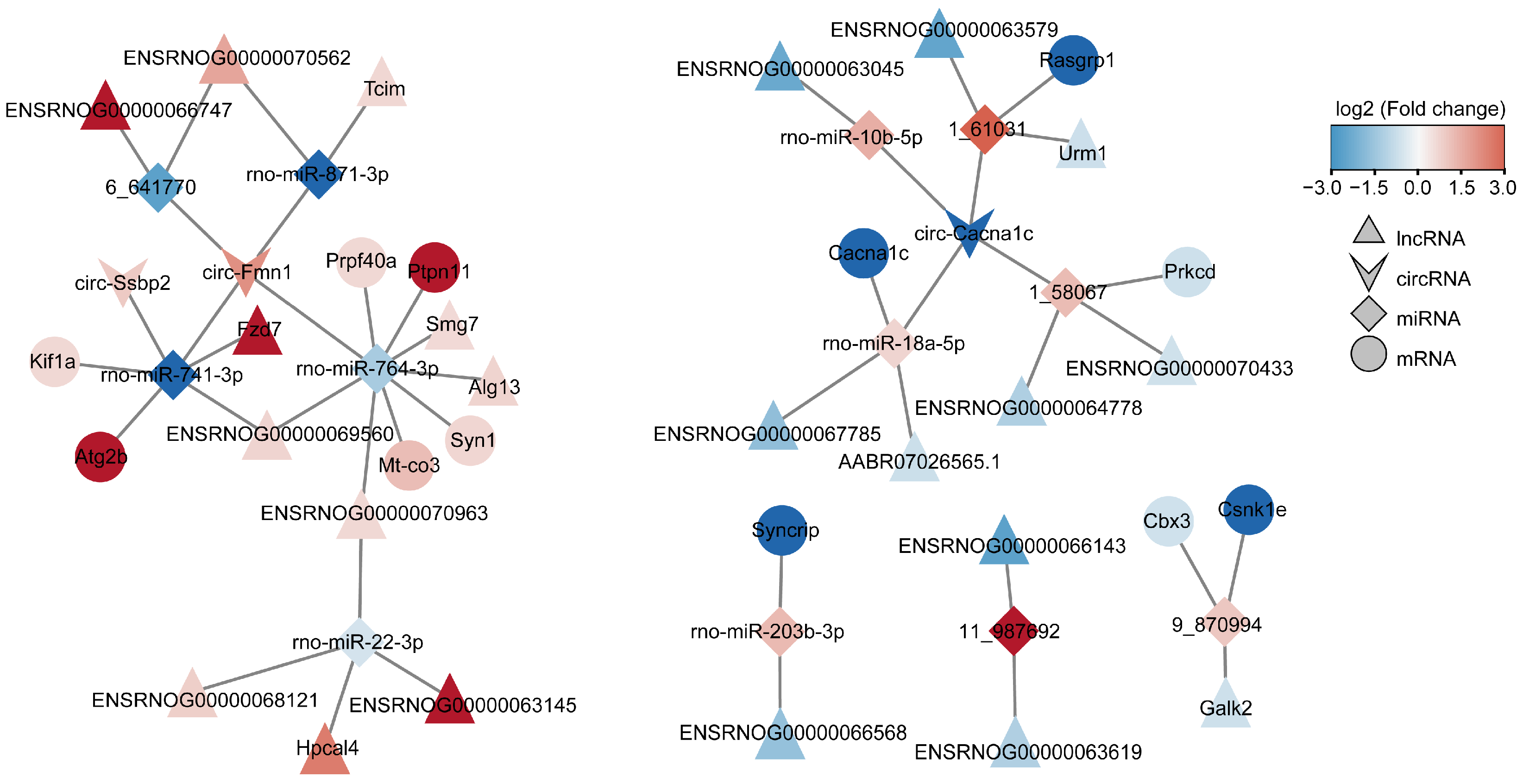
3.6. Competitive Endogenous RNA Network of Different Expression of mRNAs, lncRNAs, circRNAs, and miRNAs
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Khurana, D.; Prabhakar, S. Organophosphorus intoxication. Arch. Neurol. 2000, 57, 600–602. [Google Scholar] [CrossRef] [PubMed]
- Abou-Donia, M.B.; Siracuse, B.; Gupta, N.; Sobel Sokol, A. Sarin (GB, O-isopropyl methylphosphonofluoridate) neurotoxicity: Critical review. Crit. Rev. Toxicol. 2016, 46, 845–875. [Google Scholar] [CrossRef] [PubMed]
- Ben Abraham, R.; Rudick, V.; Weinbroum, A.A. Practical guidelines for acute care of victims of bioterrorism: Conventional injuries and concomitant nerve agent intoxication. Anesthesiology 2002, 97, 989–1004. [Google Scholar] [CrossRef] [PubMed]
- Proctor, S.P.; Heaton, K.J.; Heeren, T.; White, R.F. Effects of sarin and cyclosarin exposure during the 1991 Gulf War on neurobehavioral functioning in US army veterans. Neurotoxicology 2006, 27, 931–939. [Google Scholar] [CrossRef]
- Morita, H.; Yanagisawa, N.; Nakajima, T.; Shimizu, M. Sarin poisoning in Matsumoto, Japan. Lancet 1995, 346, 290–293. [Google Scholar] [CrossRef]
- Masuda, N.; Takatsu, M.; Morinari, H.; Ozawa, T. Sarin poisoning in Tokyo subway. Lancet 1995, 345, 1446. [Google Scholar] [CrossRef]
- Abdel-Rahman, A.; Shetty, A.K.; Abou-Donia, M.B. Acute exposure to sarin increases blood brain barrier permeability and induces neuropathological changes in the rat brain: Dose-response relationships. Neuroscience 2002, 113, 721–741. [Google Scholar] [CrossRef]
- Cruz-Hernandez, A.; Roney, A.; Goswami, D.G.; Tewari-Singh, N. A review of chemical warfare agents linked to respiratory and neurological effects experienced in Gulf War Illness. Inhal. Toxicol. 2022, 34, 412–432. [Google Scholar] [CrossRef]
- Nishiwaki, Y.; Maekawa, K.; Ogawa, Y.; Asukai, N. Effects of sarin on the nervous system in rescue team staff members and police officers 3 years after the Tokyo subway sarin attack. Environ. Health Perspect. 2001, 109, 1169–1173. [Google Scholar] [CrossRef]
- Grauer, E.; Chapman, S.; Rabinovitz, I.; Raveh, L. Single whole-body exposure to sarin vapor in rats: Long-term neuronal and behavioral deficits. Toxicol. Appl. Pharmacol. 2008, 227, 265–274. [Google Scholar] [CrossRef]
- Kassa, J.; Koupilova, M.; Vachek, J. The influence of low-level sarin inhalation exposure on spatial memory in rats. Pharmacol. Biochem. Behav. 2001, 70, 175–179. [Google Scholar] [CrossRef] [PubMed]
- Genovese, R.F.; Oubre, J.L.; Jakubowski, E.M.; Fleming, P.J. Evaluation of cognitive and biochemical effects of low-level exposure to sarin in rhesus and African green monkeys. Toxicology 2007, 231, 11–20. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.S.; Lewis, J.A.; Ippolito, D.L.; Hussainzada, N. Repeated exposure to neurotoxic levels of chlorpyrifos alters hippocampal expression of neurotrophins and neuropeptides. Toxicology 2016, 340, 53–62. [Google Scholar] [CrossRef] [PubMed]
- Farizatto, K.L.G.; Almeida, M.F.; Long, R.T.; Bahr, B.A. Early Synaptic Alterations and Selective Adhesion Signaling in Hippocampal Dendritic Zones Following Organophosphate Exposure. Sci. Rep. 2019, 9, 6532. [Google Scholar] [CrossRef]
- Li, P.; Yin, Y.L.; Zhu, M.L.; Pan, G.P. Chronic administration of isocarbophos induces vascular cognitive impairment in rats. J. Cell. Mol. Med. 2016, 20, 731–739. [Google Scholar] [CrossRef]
- Chebabo, S.R.; Santos, M.D.; Albuquerque, E.X. The organophosphate sarin, at low concentrations, inhibits the evoked release of GABA in rat hippocampal slices. Neurotoxicology 1999, 20, 871–882. [Google Scholar]
- Chao, L.L.; Kriger, S.; Buckley, S.; Ng, P. Effects of low-level sarin and cyclosarin exposure on hippocampal subfields in Gulf War Veterans. Neurotoxicology 2014, 44, 263–269. [Google Scholar] [CrossRef]
- Peng, Z.; Cheng, Y.; Tan, B.C.; Kang, L. Comprehensive analysis of RNA-Seq data reveals extensive RNA editing in a human transcriptome. Nat. Biotechnol. 2012, 30, 253–260. [Google Scholar] [CrossRef]
- Eichler, D.C. Information processing and gene expression. J. Fla. Med. Assoc. 1993, 80, 63–66. [Google Scholar]
- Cai, Y.; Yu, X.; Hu, S.; Yu, J. A brief review on the mechanisms of miRNA regulation. Genom. Proteom. Bioinform. 2009, 7, 147–154. [Google Scholar] [CrossRef]
- Bridges, M.C.; Daulagala, A.C.; Kourtidis, A. LNCcation: lncRNA localization and function. J. Cell Biol. 2021, 220, 202009045. [Google Scholar] [CrossRef] [PubMed]
- Kristensen, L.S.; Andersen, M.S.; Stagsted, L.V.W.; Ebbesen, K.K. The biogenesis, biology and characterization of circular RNAs. Nat. Rev. Genet. 2019, 20, 675–691. [Google Scholar] [CrossRef] [PubMed]
- Thomson, D.W.; Dinger, M.E. Endogenous microRNA sponges: Evidence and controversy. Nat. Rev. Genet. 2016, 17, 272–283. [Google Scholar] [CrossRef] [PubMed]
- Vasamsetti, B.M.K.; Chon, K.; Kim, J.; Oh, J.A. Transcriptome-Based Identification of Genes Responding to the Organophosphate Pesticide Phosmet in Danio rerio. Genes 2021, 12, 1738. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.; Lai, X.; Wang, X.; Ying, J. Long Non-coding RNAs and Circular RNAs: Insights into Microglia and Astrocyte Mediated Neurological Diseases. Front. Mol. Neurosci. 2021, 14, 745–762. [Google Scholar] [CrossRef]
- Ye, J.; Li, J.; Zhao, P. Roles of ncRNAs as ceRNAs in Gastric Cancer. Genes 2021, 12, 1036. [Google Scholar] [CrossRef]
- Qi, X.; Zhang, D.H.; Wu, N.; Xiao, J.H. ceRNA in cancer: Possible functions and clinical implications. J. Med. Genet. 2015, 52, 710–718. [Google Scholar] [CrossRef]
- Moreno-Garcia, L.; Lopez-Royo, T.; Calvo, A.C.; Toivonen, J.M. Competing Endogenous RNA Networks as Biomarkers in Neurodegenerative Diseases. Int. J. Mol. Sci. 2020, 21, 9582. [Google Scholar] [CrossRef]
- Pachiappan, A.; Thwin, M.M.; Weng Keong, L.; Lee, F.K. ETS2 regulating neurodegenerative signaling pathway of human neuronal (SH-SY5Y) cells exposed to single and repeated low-dose sarin (GB). Chem. Res. Toxicol. 2009, 22, 990–996. [Google Scholar] [CrossRef]
- Spradling, K.D.; Lumley, L.A.; Robison, C.L.; Meyerhoff, J.L. Transcriptional responses of the nerve agent-sensitive brain regions amygdala, hippocampus, piriform cortex, septum, and thalamus following exposure to the organophosphonate anticholinesterase sarin. J. Neuroinflamm. 2011, 8, 84. [Google Scholar] [CrossRef]
- Shah, A.; Bhat, R. Fluoxetine and sertraline may be associated with lower risk of suicide death than paroxetine in adults with depression. Evid. Based Ment. Health 2012, 15, 98. [Google Scholar] [CrossRef] [PubMed]
- Shi, M.; Deng, S.; Cui, Y.; Chen, X. Repeated low-dose exposures to sarin disrupted the homeostasis of phospholipid and sphingolipid metabolism in guinea pig hippocampus. Toxicol. Lett. 2021, 338, 32–39. [Google Scholar] [CrossRef] [PubMed]
- Chan, V.T.; Soto, A.; Wagner, J.A.; Watts, B.S. Mechanisms of Organophosphorus (OP) Injury: Sarin-Induced Hippocampal Gene Expression Changes and Pathway Perturbation; 711th Human Performance Wing: Dayton, OH, USA, 2012. [Google Scholar]
- Brimblecombe, R.W.; Green, D.M.; Stratton, J.A.; Thompson, P.B. The protective actions of some anticholinergic drugs in sarin poisoning. Br. J. Pharmacol. 1970, 39, 822–830. [Google Scholar] [CrossRef] [PubMed]
- Vorhees, C.V.; Williams, M.T. Morris water maze: Procedures for assessing spatial and related forms of learning and memory. Nat. Protoc. 2006, 1, 848–858. [Google Scholar] [CrossRef]
- Atchison, C.R.; Sheridan, R.E.; Duniho, S.M.; Shih, T.M. Development of a Guinea pig model for low-dose, long-term exposure to organophosphorus nerve agents. Toxicol. Mech. Methods 2004, 14, 183–194. [Google Scholar] [CrossRef]
- Mach, M.; Grubbs, R.D.; Price, W.A.; Nagaoka, M. Delayed behavioral and endocrine effects of sarin and stress exposure in mice. J. Appl. Toxicol. 2008, 28, 132–139. [Google Scholar] [CrossRef]
- Kassa, J.; Pecka, M.; Tichy, M.; Bajgar, J. Toxic effects of sarin in rats at three months following single or repeated low-level inhalation exposure. Pharmacol. Toxicol. 2001, 88, 209–212. [Google Scholar] [CrossRef]
- De Araujo Furtado, M.; Rossetti, F.; Chanda, S.; Yourick, D. Exposure to nerve agents: From status epilepticus to neuroinflammation, brain damage, neurogenesis and epilepsy. Neurotoxicology 2012, 33, 1476–1490. [Google Scholar] [CrossRef]
- Qujeq, D.; Roushan, T.; Norouzy, A.; Habibi-Rezaei, M. Effects of dichlorvos and carbaryl on the activity of free and immobilized acetylcholinesterase. Toxicol. Ind. Health 2012, 28, 291–295. [Google Scholar] [CrossRef]
- Moser, V.C.; Phillips, P.M.; McDaniel, K.L.; Marshall, R.S. Neurobehavioral effects of chronic dietary and repeated high-level spike exposure to chlorpyrifos in rats. Toxicol. Sci. 2005, 86, 375–386. [Google Scholar] [CrossRef]
- Scremin, O.U.; Shih, T.M.; Huynh, L.; Roch, M. Delayed neurologic and behavioral effects of subtoxic doses of cholinesterase inhibitors. J. Pharmacol. Exp. Ther. 2003, 304, 1111–1119. [Google Scholar] [CrossRef]
- Kasai, H.; Hayama, T.; Ishikawa, M.; Watanabe, S. Learning rules and persistence of dendritic spines. Eur. J. Neurosci. 2010, 32, 241–249. [Google Scholar] [CrossRef]
- Dong, J.; Fu, H.; Fu, Y.; You, M. Maternal Exposure to Di-(2-ethylhexyl) Phthalate Impairs Hippocampal Synaptic Plasticity in Male Offspring: Involvement of Damage to Dendritic Spine Development. ACS Chem. Neurosci. 2021, 12, 311–322. [Google Scholar] [CrossRef]
- Yun, S.H.; Park, K.A.; Sullivan, P.; Pasternak, J.F. Blockade of nicotinic acetylcholine receptors suppresses hippocampal long-term potentiation in wild-type but not ApoE4 targeted replacement mice. J. Neurosci. Res. 2005, 82, 771–777. [Google Scholar] [CrossRef]
- Howard, A.S.; Bucelli, R.; Jett, D.A.; Bruun, D. Chlorpyrifos exerts opposing effects on axonal and dendritic growth in primary neuronal cultures. Toxicol. Appl. Pharmacol. 2005, 207, 112–124. [Google Scholar] [CrossRef]
- Roy, T.S.; Sharma, V.; Seidler, F.J.; Slotkin, T.A. Quantitative morphological assessment reveals neuronal and glial deficits in hippocampus after a brief subtoxic exposure to chlorpyrifos in neonatal rats. Brain Res. Dev. Brain Res. 2005, 155, 71–80. [Google Scholar] [CrossRef]
- Campana, A.D.; Sanchez, F.; Gamboa, C.; Gomez-Villalobos Mde, J. Dendritic morphology on neurons from prefrontal cortex, hippocampus, and nucleus accumbens is altered in adult male mice exposed to repeated low dose of malathion. Synapse 2008, 62, 283–290. [Google Scholar] [CrossRef]
- Papadopoulos, T.; Rhee, H.J.; Subramanian, D.; Paraskevopoulou, F. Endosomal Phosphatidylinositol 3-Phosphate Promotes Gephyrin Clustering and GABAergic Neurotransmission at Inhibitory Postsynapses. J. Biol. Chem. 2017, 292, 1160–1177. [Google Scholar] [CrossRef]
- Sanchez-Santed, F.; Colomina, M.T.; Herrero Hernandez, E. Organophosphate pesticide exposure and neurodegeneration. Cortex 2016, 74, 417–426. [Google Scholar] [CrossRef]
- Myskiw, J.C.; Rossato, J.I.; Bevilaqua, L.R.; Medina, J.H. On the participation of mTOR in recognition memory. Neurobiol. Learn. Mem. 2008, 89, 338–351. [Google Scholar] [CrossRef]
- Fortress, A.M.; Fan, L.; Orr, P.T.; Zhao, Z. Estradiol-induced object recognition memory consolidation is dependent on activation of mTOR signaling in the dorsal hippocampus. Learn. Mem. 2013, 20, 147–155. [Google Scholar] [CrossRef] [PubMed]
- Hoeffer, C.A.; Klann, E. mTOR signaling: At the crossroads of plasticity, memory and disease. Trends Neurosci. 2010, 33, 67–75. [Google Scholar] [CrossRef] [PubMed]
- Bollen, E.; Prickaerts, J. Phosphodiesterases in neurodegenerative disorders. IUBMB Life 2012, 64, 965–970. [Google Scholar] [CrossRef] [PubMed]
- Todorova, V.; Blokland, A. Mitochondria and Synaptic Plasticity in the Mature and Aging Nervous System. Curr. Neuropharmacol. 2017, 15, 166–173. [Google Scholar] [CrossRef]
- Bailey, C.H.; Kandel, E.R.; Harris, K.M. Structural Components of Synaptic Plasticity and Memory Consolidation. Cold Spring Harb. Perspect. Biol. 2015, 7, a021758. [Google Scholar] [CrossRef]
- Perucho, L.; Artero-Castro, A.; Guerrero, S.; Ramon y Cajal, S. RPLP1, a crucial ribosomal protein for embryonic development of the nervous system. PLoS ONE 2014, 9, e99956. [Google Scholar] [CrossRef]
- Kandimalla, R.; Manczak, M.; Yin, X.; Wang, R. Hippocampal phosphorylated tau induced cognitive decline, dendritic spine loss and mitochondrial abnormalities in a mouse model of Alzheimer’s disease. Hum. Mol. Genet. 2018, 27, 30–40. [Google Scholar] [CrossRef]
- Li, H.; Xue, X.; Li, L.; Li, Y. Aluminum-Induced Synaptic Plasticity Impairment via PI3K-Akt-mTOR Signaling Pathway. Neurotox. Res. 2020, 37, 996–1008. [Google Scholar] [CrossRef]
- Falcicchia, C.; Tozzi, F.; Arancio, O.; Watterson, D.M. Involvement of p38 MAPK in Synaptic Function and Dysfunction. Int. J. Mol. Sci. 2020, 21, 5624. [Google Scholar] [CrossRef]
- Ma, N.; Tie, C.; Yu, B.; Zhang, W. Identifying lncRNA-miRNA-mRNA networks to investigate Alzheimer’s disease pathogenesis and therapy strategy. Aging 2020, 12, 2897–2920. [Google Scholar] [CrossRef]
- Bernard, D.; Prasanth, K.V.; Tripathi, V.; Colasse, S. A long nuclear-retained non-coding RNA regulates synaptogenesis by modulating gene expression. EMBO J. 2010, 29, 3082–3093. [Google Scholar] [CrossRef] [PubMed]
- Ku, T.; Li, B.; Gao, R.; Zhang, Y. NF-kappaB-regulated microRNA-574-5p underlies synaptic and cognitive impairment in response to atmospheric PM(2.5) aspiration. Part. Fibre Toxicol. 2017, 14, 34. [Google Scholar] [CrossRef] [PubMed]
- Gast, H.; Gordic, S.; Petrzilka, S.; Lopez, M. Transforming growth factor-beta inhibits the expression of clock genes. Ann. N. Y. Acad. Sci. 2012, 1261, 79–87. [Google Scholar] [CrossRef] [PubMed]
- Schrotter, A.; Oberhaus, A.; Kolbe, K.; Seger, S. LMD proteomics provides evidence for hippocampus field-specific motor protein abundance changes with relevance to Alzheimer’s disease. Biochim. Biophys. Acta Proteins Proteom. 2017, 1865, 703–714. [Google Scholar] [CrossRef]
- Kong, W.; Mou, X.; Zhang, N.; Zeng, W. The construction of common and specific significance subnetworks of Alzheimer’s disease from multiple brain regions. Biomed Res. Int. 2015, 2015, 394260. [Google Scholar] [CrossRef]
- Liu, W.; Liu, C.; Zhu, J.; Shu, P. MicroRNA-16 targets amyloid precursor protein to potentially modulate Alzheimer’s-associated pathogenesis in SAMP8 mice. Neurobiol. Aging 2012, 33, 522–534. [Google Scholar] [CrossRef]
- Simon-Areces, J.; Dopazo, A.; Dettenhofer, M.; Rodriguez-Tebar, A. Formin1 mediates the induction of dendritogenesis and synaptogenesis by neurogenin3 in mouse hippocampal neurons. PLoS ONE 2011, 6, e21825. [Google Scholar] [CrossRef]
- Tigaret, C.M.; Lin, T.E.; Morrell, E.R.; Sykes, L. Neurotrophin receptor activation rescues cognitive and synaptic abnormalities caused by hemizygosity of the psychiatric risk gene Cacna1c. Mol. Psychiatry 2021, 26, 1748–1760. [Google Scholar] [CrossRef] [PubMed]
- Zhao, K.; Zeng, L.; Cai, Z.; Liu, M. RNA sequencing-based identification of the regulatory mechanism of microRNAs, transcription factors, and corresponding target genes involved in vascular dementia. Front. Neurosci. 2022, 16, 917489. [Google Scholar] [CrossRef]
- Tian, T.; Zhang, Y.; Wu, T.; Yang, L. miRNA profiling in the hippocampus of attention-deficit/hyperactivity disorder rats. J. Cell Biochem. 2019, 120, 3621–3629. [Google Scholar] [CrossRef]
- Zurawek, D.; Gruca, P.; Antkiewicz-Michaluk, L.; Dziedzicka-Wasylewska, M. Resilient Phenotype in Chronic Mild Stress Paradigm Is Associated with Altered Expression Levels of miR-18a-5p and Serotonin 5-HT(1a) Receptor in Dorsal Part of the Hippocampus. Mol. Neurobiol. 2019, 56, 7680–7693. [Google Scholar] [CrossRef] [PubMed]
- Ke, X.; Huang, Y.; Fu, Q.; Lane, R.H. Adverse Maternal Environment Alters MicroRNA-10b-5p Expression and Its Epigenetic Profile Concurrently with Impaired Hippocampal Neurogenesis in Male Mouse Hippocampus. Dev. Neurosci. 2021, 43, 95–105. [Google Scholar] [CrossRef]
- Wang, Z.B.; Qu, J.; Xie, P.; Yang, Z.Q. Integrative analysis of expression profile indicates the ECM receptor and LTP dysfunction in the glioma-related epilepsy. BMC Genom. 2022, 23, 430. [Google Scholar] [CrossRef] [PubMed]
- Meseguer, S.; Mudduluru, G.; Escamilla, J.M.; Allgayer, H. MicroRNAs-10a and -10b contribute to retinoic acid-induced differentiation of neuroblastoma cells and target the alternative splicing regulatory factor SFRS1 (SF2/ASF). J. Biol. Chem. 2011, 286, 4150–4164. [Google Scholar] [CrossRef]
- Izadi, F.; Soheilifar, M.H. Exploring Potential Biomarkers Underlying Pathogenesis of Alzheimer’s Disease by Differential Co-expression Analysis. Avicenna J. Med. Biotechnol. 2018, 10, 233–241. [Google Scholar] [PubMed]
- McLeod, F.; Bossio, A.; Marzo, A.; Ciani, L. Wnt Signaling Mediates LTP-Dependent Spine Plasticity and AMPAR Localization through Frizzled-7 Receptors. Cell Rep. 2018, 23, 1060–1071. [Google Scholar] [CrossRef]
- Zhou, Y.; Li, Z.X.; Liu, Y.T.; Xu, Z.C. N-linoleyltyrosine protects neurons against Abeta(1-40)-induced cell toxicity via autophagy involving the CB(2)/AMPK/mTOR/ULK1 pathway. Brain Res. Bull. 2022, 188, 203–213. [Google Scholar] [CrossRef]
- Lo, K.Y.; Kuzmin, A.; Unger, S.M.; Petersen, J.D. KIF1A is the primary anterograde motor protein required for the axonal transport of dense-core vesicles in cultured hippocampal neurons. Neurosci. Lett. 2011, 491, 168–173. [Google Scholar] [CrossRef]
- Kusakari, S.; Saitow, F.; Ago, Y.; Shibasaki, K. Shp2 in forebrain neurons regulates synaptic plasticity, locomotion, and memory formation in mice. Mol. Cell Biol. 2015, 35, 1557–1572. [Google Scholar] [CrossRef]
- Tao-Cheng, J.H. Immunogold labeling of synaptic vesicle proteins in developing hippocampal neurons. Mol. Brain. 2020, 13, 9. [Google Scholar] [CrossRef]
- Swerdlow, R.H. Mitochondria and cell bioenergetics: Increasingly recognized components and a possible etiologic cause of Alzheimer’s disease. Antioxid. Redox Signal. 2012, 16, 1434–1455. [Google Scholar] [CrossRef] [PubMed]
- Ho, A.M.; Peyton, M.P.; Scaletty, S.J.; Trapp, S. Chronic Intermittent Ethanol Exposure Alters Behavioral Flexibility in Aged Rats Compared to Adult Rats and Modifies Protein and Protein Pathways Related to Alzheimer’s Disease. ACS Omega 2022, 7, 46260–46276. [Google Scholar] [CrossRef] [PubMed]
- Kim, B.; Tag, S.H.; Nam, E.; Ham, S. SYNCRIP controls miR-137 and striatal learning in animal models of methamphetamine abstinence. Acta Pharm. Sin. B 2022, 12, 3281–3297. [Google Scholar] [CrossRef] [PubMed]
- Levran, O.; Peles, E.; Randesi, M.; Correa da Rosa, J. Synaptic Plasticity and Signal Transduction Gene Polymorphisms and Vulnerability to Drug Addictions in Populations of European or African Ancestry. CNS Neurosci. Ther. 2015, 21, 898–904. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.; Su, T.; Xue, Y.; Cheng, C. Cbx3 maintains lineage specificity during neural differentiation. Genes Dev. 2017, 31, 241–246. [Google Scholar] [CrossRef] [PubMed]
- Eshraghi, M.; Ramirez-Jarquin, U.N.; Shahani, N.; Nuzzo, T. RasGRP1 is a causal factor in the development of l-DOPA-induced dyskinesia in Parkinson’s disease. Sci. Adv. 2020, 6, eaaz7001. [Google Scholar] [CrossRef]










Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shi, J.; Liu, D.; Jin, Q.; Chen, X.; Zhang, R.; Shi, T.; Zhu, S.; Zhang, Y.; Zong, X.; Wang, C.; et al. Whole-Transcriptome Analysis of Repeated Low-Level Sarin-Exposed Rat Hippocampus and Identification of Cerna Networks to Investigate the Mechanism of Sarin-Induced Cognitive Impairment. Biology 2023, 12, 627. https://doi.org/10.3390/biology12040627
Shi J, Liu D, Jin Q, Chen X, Zhang R, Shi T, Zhu S, Zhang Y, Zong X, Wang C, et al. Whole-Transcriptome Analysis of Repeated Low-Level Sarin-Exposed Rat Hippocampus and Identification of Cerna Networks to Investigate the Mechanism of Sarin-Induced Cognitive Impairment. Biology. 2023; 12(4):627. https://doi.org/10.3390/biology12040627
Chicago/Turabian StyleShi, Jingjing, Dongxin Liu, Qian Jin, Xuejun Chen, Ruihua Zhang, Tong Shi, Siqing Zhu, Yi Zhang, Xingxing Zong, Chen Wang, and et al. 2023. "Whole-Transcriptome Analysis of Repeated Low-Level Sarin-Exposed Rat Hippocampus and Identification of Cerna Networks to Investigate the Mechanism of Sarin-Induced Cognitive Impairment" Biology 12, no. 4: 627. https://doi.org/10.3390/biology12040627
APA StyleShi, J., Liu, D., Jin, Q., Chen, X., Zhang, R., Shi, T., Zhu, S., Zhang, Y., Zong, X., Wang, C., & Li, L. (2023). Whole-Transcriptome Analysis of Repeated Low-Level Sarin-Exposed Rat Hippocampus and Identification of Cerna Networks to Investigate the Mechanism of Sarin-Induced Cognitive Impairment. Biology, 12(4), 627. https://doi.org/10.3390/biology12040627