Uterine Transcriptome: Understanding Physiology and Disease Processes

 ,
,  ,
, {kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Background
2. Microarrays and RNA-Sequencing
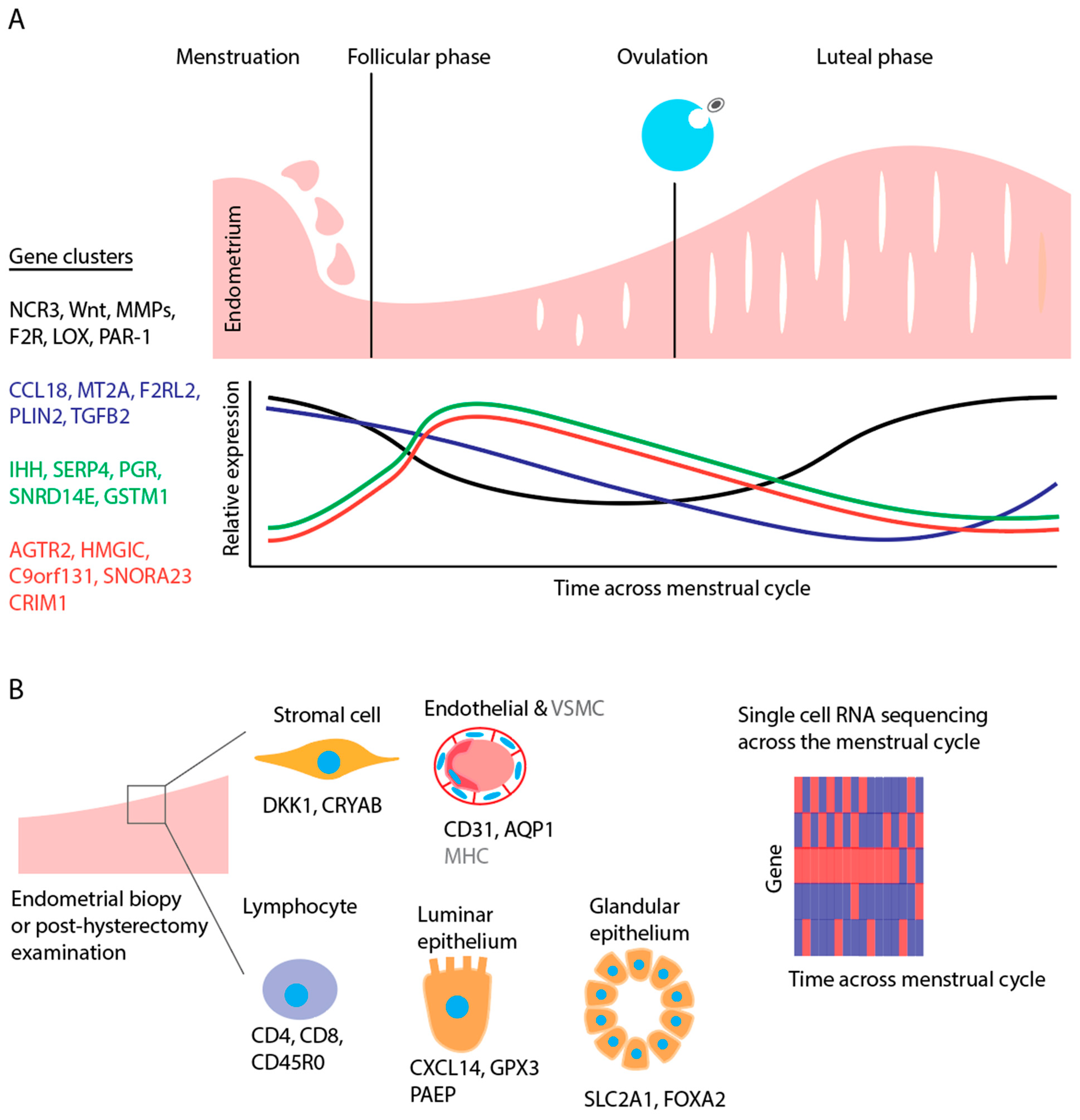
3. Endometrial Transcriptome and the Normal Menstrual Cycle
3.1. Menstruation
3.2. Proliferative and Secretory Phases
3.3. Single-Cell Transcriptomics throughout the Menstrual Cycle
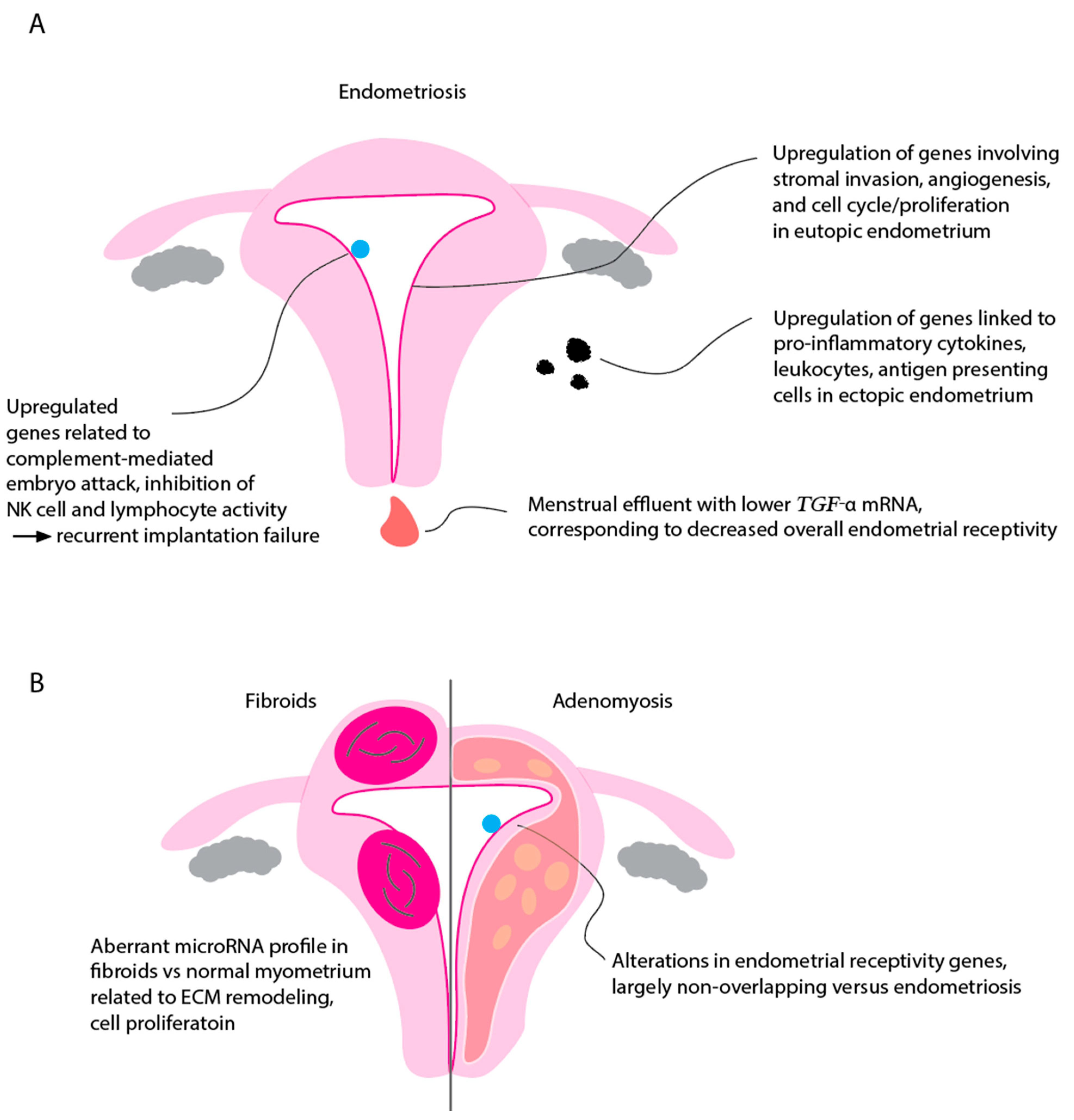
4. Endometrial Transcriptome in Endometriosis
5. Endometrial Transcriptome in Recurrent Implantation Failure
6. Interplay between Endometrium and Immune System Revealed through Transcriptomics
7. Transcriptome Analysis in Other Benign Gynecological Conditions
7.1. Fibroid Transcriptomics
7.2. Adenomyosis Transcriptomics
8. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Salamonsen, L.A.; Hannan, N.J.; Dimitriadis, E. Cytokines and chemokines during human embryo implantation: Roles in implantation and early placentation. Semin. Reprod. Med. 2007, 25, 437–444. [Google Scholar] [CrossRef] [PubMed]
- Diaz-Gimeno, P.; Ruiz-Alonso, M.; Blesa, D.; Simon, C. Transcriptomics of the human endometrium. Int. J. Dev. Biol. 2014, 58, 127–137. [Google Scholar] [CrossRef] [PubMed]
- Rogers, P.A.W. Structure and function of endometrial blood vessels. Hum. Reprod. Update 1996, 2, 57–62. [Google Scholar] [CrossRef] [PubMed]
- Gargett, C.E.; Nguyen, H.P.; Ye, L. Endometrial regeneration and endometrial stem/progenitor cells. Rev. Endocr. Metab. Disord. 2012, 13, 235–251. [Google Scholar] [CrossRef]
- Stadtmauer, D.J.; Wagner, G.P. Single-cell analysis of prostaglandin E2-induced human decidual cell in vitro differentiation: A minimal ancestral deciduogenic signaldagger. Biol. Reprod. 2022, 106, 155–172. [Google Scholar] [CrossRef]
- Fitzgerald, H.C.; Kelleher, A.M.; Ranjit, C.; Schust, D.J.; Spencer, T.E. Basolateral secretions of human endometrial epithelial organoids impact stromal cell decidualization. Mol. Hum. Reprod. 2023, 29, gaad007. [Google Scholar] [CrossRef]
- Chen, L.; Qu, J.; Xiang, C. The multi-functional roles of menstrual blood-derived stem cells in regenerative medicine. Stem Cell Res. Ther. 2019, 10, 1. [Google Scholar] [CrossRef]
- Bozorgmehr, M.; Gurung, S.; Darzi, S.; Nikoo, S.; Kazemnejad, S.; Zarnani, A.H.; Gargett, C.E. Endometrial and Menstrual Blood Mesenchymal Stem/Stromal Cells: Biological Properties and Clinical Application. Front. Cell Dev. Biol. 2020, 8, 497. [Google Scholar] [CrossRef]
- Ponnampalam, A.P.; Weston, G.C.; Trajstman, A.C.; Susil, B.; Rogers, P.A. Molecular classification of human endometrial cycle stages by transcriptional profiling. Mol. Hum. Reprod. 2004, 10, 879–893. [Google Scholar] [CrossRef]
- Arian, S.E.; Hessami, K.; Khatibi, A.; To, A.K.; Shamshirsaz, A.A.; Gibbons, W. Endometrial Receptivity Array Before Frozen Embryo Transfer Cycles: A Systematic Review and Meta-analysis. Fertil. Steril. 2022, 119, 229–238. [Google Scholar] [CrossRef]
- Altmae, S.; Martinez-Conejero, J.A.; Salumets, A.; Simon, C.; Horcajadas, J.A.; Stavreus-Evers, A. Endometrial gene expression analysis at the time of embryo implantation in women with unexplained infertility. Mol. Hum. Reprod. 2010, 16, 178–187. [Google Scholar] [CrossRef] [PubMed]
- Koler, M.; Achache, H.; Tsafrir, A.; Smith, Y.; Revel, A.; Reich, R. Disrupted gene pattern in patients with repeated in vitro fertilization (IVF) failure. Hum. Reprod. 2009, 24, 2541–2548. [Google Scholar] [CrossRef] [PubMed]
- Habermann, J.K.; Bundgen, N.K.; Gemoll, T.; Hautaniemi, S.; Lundgren, C.; Wangsa, D.; Doering, J.; Bruch, H.P.; Nordstroem, B.; Roblick, U.J.; et al. Genomic instability influences the transcriptome and proteome in endometrial cancer subtypes. Mol. Cancer 2011, 10, 132. [Google Scholar] [CrossRef]
- Penariol, L.B.C.; Thome, C.H.; Tozetti, P.A.; Paier, C.R.K.; Buono, F.O.; Peronni, K.C.; Orellana, M.D.; Covas, D.T.; Moraes, M.E.A.; Silva, W.A., Jr.; et al. What Do the Transcriptome and Proteome of Menstrual Blood-Derived Mesenchymal Stem Cells Tell Us about Endometriosis? Int. J. Mol. Sci. 2022, 23, 11515. [Google Scholar] [CrossRef] [PubMed]
- Juarez-Barber, E.; Cozzolino, M.; Corachan, A.; Alecsandru, D.; Pellicer, N.; Pellicer, A.; Ferrero, H. Adjustment of progesterone administration after endometrial transcriptomic analysis does not improve reproductive outcomes in women with adenomyosis. Reprod. Biomed. Online 2023, 46, 99–106. [Google Scholar] [CrossRef]
- Ruiz-Alonso, M.; Blesa, D.; Simon, C. The genomics of the human endometrium. Biochim. Biophys. Acta 2012, 1822, 1931–1942. [Google Scholar] [CrossRef]
- Bellver, J.; Mundi, M.; Esteban, F.J.; Mosquera, S.; Horcajadas, J.A. ‘-omics’ technology and human reproduction: Reproductomics. Expert Rev. Obstet. Gynecol. 2014, 5, 493–506. [Google Scholar] [CrossRef]
- Altmae, S.; Esteban, F.J.; Stavreus-Evers, A.; Simon, C.; Giudice, L.; Lessey, B.A.; Horcajadas, J.A.; Macklon, N.S.; D’Hooghe, T.; Campoy, C.; et al. Guidelines for the design, analysis and interpretation of ‘omics’ data: Focus on human endometrium. Hum. Reprod. Update 2014, 20, 12–28. [Google Scholar] [CrossRef]
- Krjutskov, K.; Katayama, S.; Saare, M.; Vera-Rodriguez, M.; Lubenets, D.; Samuel, K.; Laisk-Podar, T.; Teder, H.; Einarsdottir, E.; Salumets, A.; et al. Single-cell transcriptome analysis of endometrial tissue. Hum. Reprod. 2016, 31, 844–853. [Google Scholar] [CrossRef]
- Buettner, F.; Natarajan, K.N.; Casale, F.P.; Proserpio, V.; Scialdone, A.; Theis, F.J.; Teichmann, S.A.; Marioni, J.C.; Stegle, O. Computational analysis of cell-to-cell heterogeneity in single-cell RNA-sequencing data reveals hidden subpopulations of cells. Nat. Biotechnol. 2015, 33, 155–160. [Google Scholar] [CrossRef]
- Grun, D.; Lyubimova, A.; Kester, L.; Wiebrands, K.; Basak, O.; Sasaki, N.; Clevers, H.; van Oudenaarden, A. Single-cell messenger RNA sequencing reveals rare intestinal cell types. Nature 2015, 525, 251–255. [Google Scholar] [CrossRef] [PubMed]
- Vassena, R.; Eguizabal, C.; Heindryckx, B.; Sermon, K.; Simon, C.; van Pelt, A.M.; Veiga, A.; Zambelli, F.; on behalf of the ESHRE special interest group Stem Cells. Stem cells in reproductive medicine: Ready for the patient? Hum. Reprod. 2015, 30, 2014–2021. [Google Scholar] [CrossRef] [PubMed]
- Retis-Resendiz, A.M.; Gonzalez-Garcia, I.N.; Leon-Juarez, M.; Camacho-Arroyo, I.; Cerbon, M.; Vazquez-Martinez, E.R. The role of epigenetic mechanisms in the regulation of gene expression in the cyclical endometrium. Clin. Epigenet. 2021, 13, 116. [Google Scholar] [CrossRef]
- Critchley, H.O.; Robertson, K.A.; Forster, T.; Henderson, T.A.; Williams, A.R.; Ghazal, P. Gene expression profiling of mid to late secretory phase endometrial biopsies from women with menstrual complaint. Am. J. Obstet. Gynecol. 2006, 195, 406–414.e7. [Google Scholar] [CrossRef]
- Punyadeera, C.; Dassen, H.; Klomp, J.; Dunselman, G.; Kamps, R.; Dijcks, F.; Ederveen, A.; de Goeij, A.; Groothuis, P. Oestrogen-modulated gene expression in the human endometrium. Cell Mol. Life Sci. 2005, 62, 239–250. [Google Scholar] [CrossRef] [PubMed]
- Pende, D.; Parolini, S.; Pessino, A.; Sivori, S.; Augugliaro, R.; Morelli, L.; Marcenaro, E.; Accame, L.; Malaspina, A.; Biassoni, R.; et al. Identification and molecular characterization of NKp30, a novel triggering receptor involved in natural cytotoxicity mediated by human natural killer cells. J. Exp. Med. 1999, 190, 1505–1516. [Google Scholar] [CrossRef] [PubMed]
- Miller, C.; Sassoon, D.A. Wnt-7a maintains appropriate uterine patterning during the development of the mouse female reproductive tract. Development 1998, 125, 3201–3211. [Google Scholar] [CrossRef]
- Salamonsen, L.A.; Woolley, D.E. Matrix metalloproteinases in normal menstruation. Hum. Reprod. 1996, 11 (Suppl. S2), 124–133. [Google Scholar] [CrossRef]
- Petracco, R.G.; Kong, A.; Grechukhina, O.; Krikun, G.; Taylor, H.S. Global gene expression profiling of proliferative phase endometrium reveals distinct functional subdivisions. Reprod. Sci. 2012, 19, 1138–1145. [Google Scholar] [CrossRef]
- Li, X.F.; Ahmed, A. Dual role of angiotensin II in the human endometrium. Hum. Reprod. 1996, 11 (Suppl. S2), 95–108. [Google Scholar] [CrossRef]
- Wang, W.; Vilella, F.; Alama, P.; Moreno, I.; Mignardi, M.; Isakova, A.; Pan, W.; Simon, C.; Quake, S.R. Single-cell transcriptomic atlas of the human endometrium during the menstrual cycle. Nat. Med. 2020, 26, 1644–1653. [Google Scholar] [CrossRef] [PubMed]
- Sigurgeirsson, B.; Amark, H.; Jemt, A.; Ujvari, D.; Westgren, M.; Lundeberg, J.; Gidlof, S. Comprehensive RNA sequencing of healthy human endometrium at two time points of the menstrual cycle. Biol. Reprod. 2017, 96, 24–33. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Alonso, L.; Handfield, L.F.; Roberts, K.; Nikolakopoulou, K.; Fernando, R.C.; Gardner, L.; Woodhams, B.; Arutyunyan, A.; Polanski, K.; Hoo, R.; et al. Mapping the temporal and spatial dynamics of the human endometrium in vivo and in vitro. Nat. Genet. 2021, 53, 1698–1711. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Gu, C.; Ye, M.; Zhang, Z.; Han, W.; Fan, W.; Meng, Y. Identification of global transcriptome abnormalities and potential biomarkers in eutopic endometria of women with endometriosis: A preliminary study. Biomed. Rep. 2017, 6, 654–662. [Google Scholar] [CrossRef] [PubMed]
- May, K.E.; Villar, J.; Kirtley, S.; Kennedy, S.H.; Becker, C.M. Endometrial alterations in endometriosis: A systematic review of putative biomarkers. Hum. Reprod. Update 2011, 17, 637–653. [Google Scholar] [CrossRef]
- AlAshqar, A.; Reschke, L.; Kirschen, G.W.; Borahay, M.A. Role of inflammation in benign gynecologic disorders: From pathogenesis to novel therapiesdagger. Biol. Reprod. 2021, 105, 7–31. [Google Scholar] [CrossRef]
- Ulukus, M.; Ulukus, E.C.; Tavmergen Goker, E.N.; Tavmergen, E.; Zheng, W.; Arici, A. Expression of interleukin-8 and monocyte chemotactic protein 1 in women with endometriosis. Fertil. Steril. 2009, 91, 687–693. [Google Scholar] [CrossRef]
- Kyama, C.M.; Overbergh, L.; Mihalyi, A.; Meuleman, C.; Mwenda, J.M.; Mathieu, C.; D’Hooghe, T.M. Endometrial and peritoneal expression of aromatase, cytokines, and adhesion factors in women with endometriosis. Fertil. Steril. 2008, 89, 301–310. [Google Scholar] [CrossRef]
- Burney, R.O.; Talbi, S.; Hamilton, A.E.; Vo, K.C.; Nyegaard, M.; Nezhat, C.R.; Lessey, B.A.; Giudice, L.C. Gene expression analysis of endometrium reveals progesterone resistance and candidate susceptibility genes in women with endometriosis. Endocrinology 2007, 148, 3814–3826. [Google Scholar] [CrossRef]
- Ahn, S.H.; Khalaj, K.; Young, S.L.; Lessey, B.A.; Koti, M.; Tayade, C. Immune-inflammation gene signatures in endometriosis patients. Fertil. Steril. 2016, 106, 1420–1431.e7. [Google Scholar] [CrossRef]
- Matsuzaki, S.; Canis, M.; Vaurs-Barriere, C.; Boespflug-Tanguy, O.; Dastugue, B.; Mage, G. DNA microarray analysis of gene expression in eutopic endometrium from patients with deep endometriosis using laser capture microdissection. Fertil. Steril. 2005, 84 (Suppl. S2), 1180–1190. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Strawn, E.; Basir, Z.; Wang, Y.; Halverson, G.; Jailwala, P.; Guo, S.W. Genomic alterations in ectopic and eutopic endometria of women with endometriosis. Gynecol. Obstet. Investig. 2006, 62, 148–159. [Google Scholar] [CrossRef] [PubMed]
- Fassbender, A.; Verbeeck, N.; Bornigen, D.; Kyama, C.M.; Bokor, A.; Vodolazkaia, A.; Peeraer, K.; Tomassetti, C.; Meuleman, C.; Gevaert, O.; et al. Combined mRNA microarray and proteomic analysis of eutopic endometrium of women with and without endometriosis. Hum. Reprod. 2012, 27, 2020–2029. [Google Scholar] [CrossRef] [PubMed]
- Tapia, A.; Gangi, L.M.; Zegers-Hochschild, F.; Balmaceda, J.; Pommer, R.; Trejo, L.; Pacheco, I.M.; Salvatierra, A.M.; Henriquez, S.; Quezada, M.; et al. Differences in the endometrial transcript profile during the receptive period between women who were refractory to implantation and those who achieved pregnancy. Hum. Reprod. 2008, 23, 340–351. [Google Scholar] [CrossRef] [PubMed]
- Kao, L.C.; Germeyer, A.; Tulac, S.; Lobo, S.; Yang, J.P.; Taylor, R.N.; Osteen, K.; Lessey, B.A.; Giudice, L.C. Expression profiling of endometrium from women with endometriosis reveals candidate genes for disease-based implantation failure and infertility. Endocrinology 2003, 144, 2870–2881. [Google Scholar] [CrossRef]
- Okamoto, N.; Uchida, A.; Takakura, K.; Kariya, Y.; Kanzaki, H.; Riittinen, L.; Koistinen, R.; Seppala, M.; Mori, T. Suppression by human placental protein 14 of natural killer cell activity. Am. J. Reprod. Immunol. 1991, 26, 137–142. [Google Scholar] [CrossRef]
- Dominguez, F.; Galan, A.; Martin, J.J.; Remohi, J.; Pellicer, A.; Simon, C. Hormonal and embryonic regulation of chemokine receptors CXCR1, CXCR4, CCR5 and CCR2B in the human endometrium and the human blastocyst. Mol. Hum. Reprod. 2003, 9, 189–198. [Google Scholar] [CrossRef]
- Vento-Tormo, R.; Efremova, M.; Botting, R.A.; Turco, M.Y.; Vento-Tormo, M.; Meyer, K.B.; Park, J.E.; Stephenson, E.; Polanski, K.; Goncalves, A.; et al. Single-cell reconstruction of the early maternal-fetal interface in humans. Nature 2018, 563, 347–353. [Google Scholar] [CrossRef]
- Gorivodsky, M.; Torchinsky, A.; Shepshelovich, J.; Savion, S.; Fein, A.; Carp, H.; Toder, V. Colony-stimulating factor-1 (CSF-1) expression in the uteroplacental unit of mice with spontaneous and induced pregnancy loss. Clin. Exp. Immunol. 1999, 117, 540–549. [Google Scholar] [CrossRef]
- Lucas, E.S.; Vrljicak, P.; Muter, J.; Diniz-da-Costa, M.M.; Brighton, P.J.; Kong, C.S.; Lipecki, J.; Fishwick, K.J.; Odendaal, J.; Ewington, L.J.; et al. Recurrent pregnancy loss is associated with a pro-senescent decidual response during the peri-implantation window. Commun. Biol. 2020, 3, 37. [Google Scholar] [CrossRef]
- Rawlings, T.M.; Makwana, K.; Taylor, D.M.; Mole, M.A.; Fishwick, K.J.; Tryfonos, M.; Odendaal, J.; Hawkes, A.; Zernicka-Goetz, M.; Hartshorne, G.M.; et al. Modelling the impact of decidual senescence on embryo implantation in human endometrial assembloids. eLife 2021, 10, e69603. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Jin, N.; Qin, H.; Shi, X.; Liu, Y.; Cheung, W.; Wang, C.C.; Chan, T.F.; Li, T.C. Transcriptomic profiles in peripheral blood between women with unexplained recurrent implantation failure and recurrent miscarriage and the correlation with endometrium: A pilot study. PLoS ONE 2017, 12, e0189159. [Google Scholar] [CrossRef]
- Da Broi, M.G.; Meola, J.; Placa, J.R.; Peronni, K.C.; Rocha, C.V.; Silva, W.A.; Ferriani, R.A.; Navarro, P.A. Is the profile of transcripts altered in the eutopic endometrium of infertile women with endometriosis during the implantation window? Hum. Reprod. 2019, 34, 2381–2390. [Google Scholar] [CrossRef] [PubMed]
- Oertelt-Prigione, S. Immunology and the menstrual cycle. Autoimmun. Rev. 2012, 11, A486–A492. [Google Scholar] [CrossRef]
- Arruvito, L.; Sanz, M.; Banham, A.H.; Fainboim, L. Expansion of CD4 + CD25 + and FOXP3+ regulatory T cells during the follicular phase of the menstrual cycle: Implications for human reproduction. J. Immunol. 2007, 178, 2572–2578. [Google Scholar] [CrossRef] [PubMed]
- Keller, C.C.; Eikmans, M.; van der Hoorn, M.P.; Lashley, L. Recurrent miscarriages and the association with regulatory T cells; A systematic review. J. Reprod. Immunol. 2020, 139, 103105. [Google Scholar] [CrossRef]
- Ma, M.; Wang, M.; Xu, F.; Hao, S. The Imbalance in Th17 and Treg Cells in Polycystic Ovarian Syndrome Patients with Autoimmune Thyroiditis. Immunol. Investig. 2022, 51, 1170–1181. [Google Scholar] [CrossRef]
- Miller, J.E.; Lingegowda, H.; Sisnett, D.J.; Metz, C.N.; Gregersen, P.K.; Koti, M.; Tayade, C. T helper 17 axis and endometrial macrophage disruption in menstrual effluent provides potential insights into the pathogenesis of endometriosis. F S Sci. 2022, 3, 279–287. [Google Scholar] [CrossRef]
- Hansard, L.J.; Healy-Gardner, B.E.; Drapkin, A.T.; Bentley, R.C.; McLachlan, J.A.; Walmer, D.K. Human endometrial transforming growth factor-alpha: A transmembrane, surface epithelial protein that transiently disappears during the midsecretory phase of the menstrual cycle. J. Soc. Gynecol. Investig. 1997, 4, 160–166. [Google Scholar] [CrossRef]
- Tan, Y.; Flynn, W.F.; Sivajothi, S.; Luo, D.; Bozal, S.B.; Dave, M.; Luciano, A.A.; Robson, P.; Luciano, D.E.; Courtois, E.T. Single-cell analysis of endometriosis reveals a coordinated transcriptional programme driving immunotolerance and angiogenesis across eutopic and ectopic tissues. Nat. Cell Biol. 2022, 24, 1306–1318. [Google Scholar] [CrossRef]
- Wang, X.Q.; Li, D.J. The mechanisms by which trophoblast-derived molecules induce maternal-fetal immune tolerance. Cell Mol. Immunol. 2020, 17, 1204–1207. [Google Scholar] [CrossRef] [PubMed]
- Yougbare, I.; Tai, W.S.; Zdravic, D.; Oswald, B.E.; Lang, S.; Zhu, G.; Leong-Poi, H.; Qu, D.; Yu, L.; Dunk, C.; et al. Activated NK cells cause placental dysfunction and miscarriages in fetal alloimmune thrombocytopenia. Nat. Commun. 2017, 8, 224. [Google Scholar] [CrossRef]
- Pan, D.; Liu, Q.; Du, L.; Yang, Y.; Jiang, G. Polarization disorder of decidual NK cells in unexplained recurrent spontaneous abortion revealed by single-cell transcriptome analysis. Reprod. Biol. Endocrinol. 2022, 20, 108. [Google Scholar] [CrossRef]
- Parker, W.H. Etiology, symptomatology, and diagnosis of uterine myomas. Fertil. Steril. 2007, 87, 725–736. [Google Scholar] [CrossRef] [PubMed]
- Divakar, H. Asymptomatic uterine fibroids. Best Pract. Res. Clin. Obstet. Gynaecol. 2008, 22, 643–654. [Google Scholar] [CrossRef]
- Wegienka, G.; Baird, D.D.; Hertz-Picciotto, I.; Harlow, S.D.; Steege, J.F.; Hill, M.C.; Schectman, J.M.; Hartmann, K.E. Self-reported heavy bleeding associated with uterine leiomyomata. Obstet. Gynecol. 2003, 101, 431–437. [Google Scholar] [CrossRef]
- Lippman, S.A.; Warner, M.; Samuels, S.; Olive, D.; Vercellini, P.; Eskenazi, B. Uterine fibroids and gynecologic pain symptoms in a population-based study. Fertil. Steril. 2003, 80, 1488–1494. [Google Scholar] [CrossRef] [PubMed]
- Bochenska, K.; Lewitt, T.; Marsh, E.E.; Pidaparti, M.; Lewicky-Gaupp, C.; Mueller, M.G.; Kenton, K. Fibroids and Urinary Symptoms Study (FUSS). Female Pelvic. Med. Reconstr. Surg. 2021, 27, e481–e483. [Google Scholar] [CrossRef]
- Casini, M.L.; Rossi, F.; Agostini, R.; Unfer, V. Effects of the position of fibroids on fertility. Gynecol. Endocrinol. 2006, 22, 106–109. [Google Scholar] [CrossRef]
- Pritts, E.A.; Parker, W.H.; Olive, D.L. Fibroids and infertility: An updated systematic review of the evidence. Fertil. Steril. 2009, 91, 1215–1223. [Google Scholar] [CrossRef]
- Kim, G.; Jang, G.; Song, J.; Kim, D.; Lee, S.; Joo, J.W.J.; Jang, W. A transcriptome-wide association study of uterine fibroids to identify potential genetic markers and toxic chemicals. PLoS ONE 2022, 17, e0274879. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Lee, J.; Jun, J.H. Identification of differentially expressed microRNAs in outgrowth embryos compared with blastocysts and non-outgrowth embryos in mice. Reprod. Fertil. Dev. 2019, 31, 645–657. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.J.; Kim, Y.Y.; Shin, J.H.; Kim, H.; Ku, S.Y.; Suh, C.S. Variation in MicroRNA Expression Profile of Uterine Leiomyoma with Endometrial Cavity Distortion and Endometrial Cavity Non-Distortion. Int. J. Mol. Sci. 2018, 19, 2524. [Google Scholar] [CrossRef]
- Kim, M.; Kang, D.; Kwon, M.Y.; Lee, H.J.; Kim, M.J. MicroRNAs as potential indicators of the development and progression of uterine leiomyoma. PLoS ONE 2022, 17, e0268793. [Google Scholar] [CrossRef] [PubMed]
- Karmon, A.E.; Cardozo, E.R.; Rueda, B.R.; Styer, A.K. MicroRNAs in the development and pathobiology of uterine leiomyomata: Does evidence support future strategies for clinical intervention? Hum. Reprod. Update 2014, 20, 670–687. [Google Scholar] [CrossRef] [PubMed]
- Lycoudi, A.; Mavreli, D.; Mavrou, A.; Papantoniou, N.; Kolialexi, A. miRNAs in pregnancy-related complications. Expert. Rev. Mol. Diagn. 2015, 15, 999–1010. [Google Scholar] [CrossRef]
- Kim, J.S.; Kurie, J.M.; Ahn, Y.H. BMP4 depletion by miR-200 inhibits tumorigenesis and metastasis of lung adenocarcinoma cells. Mol. Cancer 2015, 14, 173. [Google Scholar] [CrossRef]
- Wu, X.; Ling, J.; Fu, Z.; Ji, C.; Wu, J.; Xu, Q. Effects of miRNA-197 overexpression on proliferation, apoptosis and migration in levonorgestrel treated uterine leiomyoma cells. Biomed. Pharmacother. 2015, 71, 1–6. [Google Scholar] [CrossRef]
- Marsh, E.E.; Steinberg, M.L.; Parker, J.B.; Wu, J.; Chakravarti, D.; Bulun, S.E. Decreased expression of microRNA-29 family in leiomyoma contributes to increased major fibrillar collagen production. Fertil. Steril. 2016, 106, 766–772. [Google Scholar] [CrossRef]
- Tian, R.; Wang, Z.; Shi, Z.; Li, D.; Wang, Y.; Zhu, Y.; Lin, W.; Gui, Y.; Zheng, X.L. Differential expression of G-protein-coupled estrogen receptor-30 in human myometrial and uterine leiomyoma smooth muscle. Fertil. Steril. 2013, 99, 256–263.e253. [Google Scholar] [CrossRef]
- Morikawa, A.; Ohara, N.; Xu, Q.; Nakabayashi, K.; DeManno, D.A.; Chwalisz, K.; Yoshida, S.; Maruo, T. Selective progesterone receptor modulator asoprisnil down-regulates collagen synthesis in cultured human uterine leiomyoma cells through up-regulating extracellular matrix metalloproteinase inducer. Hum. Reprod. 2008, 23, 944–951. [Google Scholar] [CrossRef] [PubMed]
- Wolanska, M.; Sobolewski, K.; Bankowski, E.; Jaworski, S. Matrix metalloproteinases of human leiomyoma in various stages of tumor growth. Gynecol. Obstet. Investig. 2004, 58, 14–18. [Google Scholar] [CrossRef] [PubMed]
- Aghajanova, L.; Houshdaran, S.; Irwin, J.C.; Giudice, L.C. Effects of noncavity-distorting fibroids on endometrial gene expression and function. Biol. Reprod. 2017, 97, 564–576. [Google Scholar] [CrossRef]
- Rackow, B.W.; Taylor, H.S. Submucosal uterine leiomyomas have a global effect on molecular determinants of endometrial receptivity. Fertil. Steril. 2010, 93, 2027–2034. [Google Scholar] [CrossRef]
- Pontis, A.; D’Alterio, M.N.; Pirarba, S.; de Angelis, C.; Tinelli, R.; Angioni, S. Adenomyosis: A systematic review of medical treatment. Gynecol. Endocrinol. 2016, 32, 696–700. [Google Scholar] [CrossRef] [PubMed]
- Bourdon, M.; Santulli, P.; Marcellin, L.; Maignien, C.; Maitrot-Mantelet, L.; Bordonne, C.; Plu Bureau, G.; Chapron, C. Adenomyosis: An update regarding its diagnosis and clinical features. J. Gynecol. Obstet. Hum. Reprod. 2021, 50, 102228. [Google Scholar] [CrossRef]
- Puente, J.M.; Fabris, A.; Patel, J.; Patel, A.; Cerrillo, M.; Requena, A.; Garcia-Velasco, J.A. Adenomyosis in infertile women: Prevalence and the role of 3D ultrasound as a marker of severity of the disease. Reprod. Biol. Endocrinol. 2016, 14, 60. [Google Scholar] [CrossRef]
- Buggio, L.; Dridi, D.; Barbara, G. Adenomyosis: Impact on Fertility and Obstetric Outcomes. Reprod. Sci. 2021, 28, 3081–3084. [Google Scholar] [CrossRef]
- Squillace, A.L.A.; Simonian, D.S.; Allegro, M.C.; Borges, E.J.; Bianchi, P.H.M.; Bibancos, M. Adenomyosis and in vitro fertilization impacts—A literature review. JBRA Assist. Reprod. 2021, 25, 303–309. [Google Scholar] [CrossRef]
- Ruiz-Alonso, M.; Valbuena, D.; Gomez, C.; Cuzzi, J.; Simon, C. Endometrial Receptivity Analysis (ERA): Data versus opinions. Hum. Reprod. Open 2021, 2021, hoab011. [Google Scholar] [CrossRef]
- Liu, Z.; Liu, X.; Wang, M.; Zhao, H.; He, S.; Lai, S.; Qu, Q.; Wang, X.; Zhao, D.; Bao, H. The Clinical Efficacy of Personalized Embryo Transfer Guided by the Endometrial Receptivity Array/Analysis on IVF/ICSI Outcomes: A Systematic Review and Meta-Analysis. Front. Physiol. 2022, 13, 841437. [Google Scholar] [CrossRef] [PubMed]
- Prasnikar, E.; Kunej, T.; Knez, J.; Repnik, K.; Potocnik, U.; Kovacic, B. Determining the Molecular Background of Endometrial Receptivity in Adenomyosis. Biomolecules 2020, 10, 1311. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Sun, Z.; Liu, H.; Niu, W.; Wang, X.; Liang, N.; Wang, X.; Wang, Y.; Shi, Y.; Xu, L.; et al. Single-cell transcriptomic analysis of eutopic endometrium and ectopic lesions of adenomyosis. Cell Biosci. 2021, 11, 51. [Google Scholar] [CrossRef] [PubMed]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kirschen, G.W.; Hessami, K.; AlAshqar, A.; Afrin, S.; Lulseged, B.; Borahay, M. Uterine Transcriptome: Understanding Physiology and Disease Processes. Biology 2023, 12, 634. https://doi.org/10.3390/biology12040634
Kirschen GW, Hessami K, AlAshqar A, Afrin S, Lulseged B, Borahay M. Uterine Transcriptome: Understanding Physiology and Disease Processes. Biology. 2023; 12(4):634. https://doi.org/10.3390/biology12040634
Chicago/Turabian StyleKirschen, Gregory W., Kamran Hessami, Abdelrahman AlAshqar, Sadia Afrin, Bethlehem Lulseged, and Mostafa Borahay. 2023. "Uterine Transcriptome: Understanding Physiology and Disease Processes" Biology 12, no. 4: 634. https://doi.org/10.3390/biology12040634
APA StyleKirschen, G. W., Hessami, K., AlAshqar, A., Afrin, S., Lulseged, B., & Borahay, M. (2023). Uterine Transcriptome: Understanding Physiology and Disease Processes. Biology, 12(4), 634. https://doi.org/10.3390/biology12040634