
Observation of Histone H2AX Phosphorylation by Radiation-Induced Bystander Response Using Titanium Characteristic X-ray Microbeam

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Cell Culture
2.2. Microbeam Irradiation
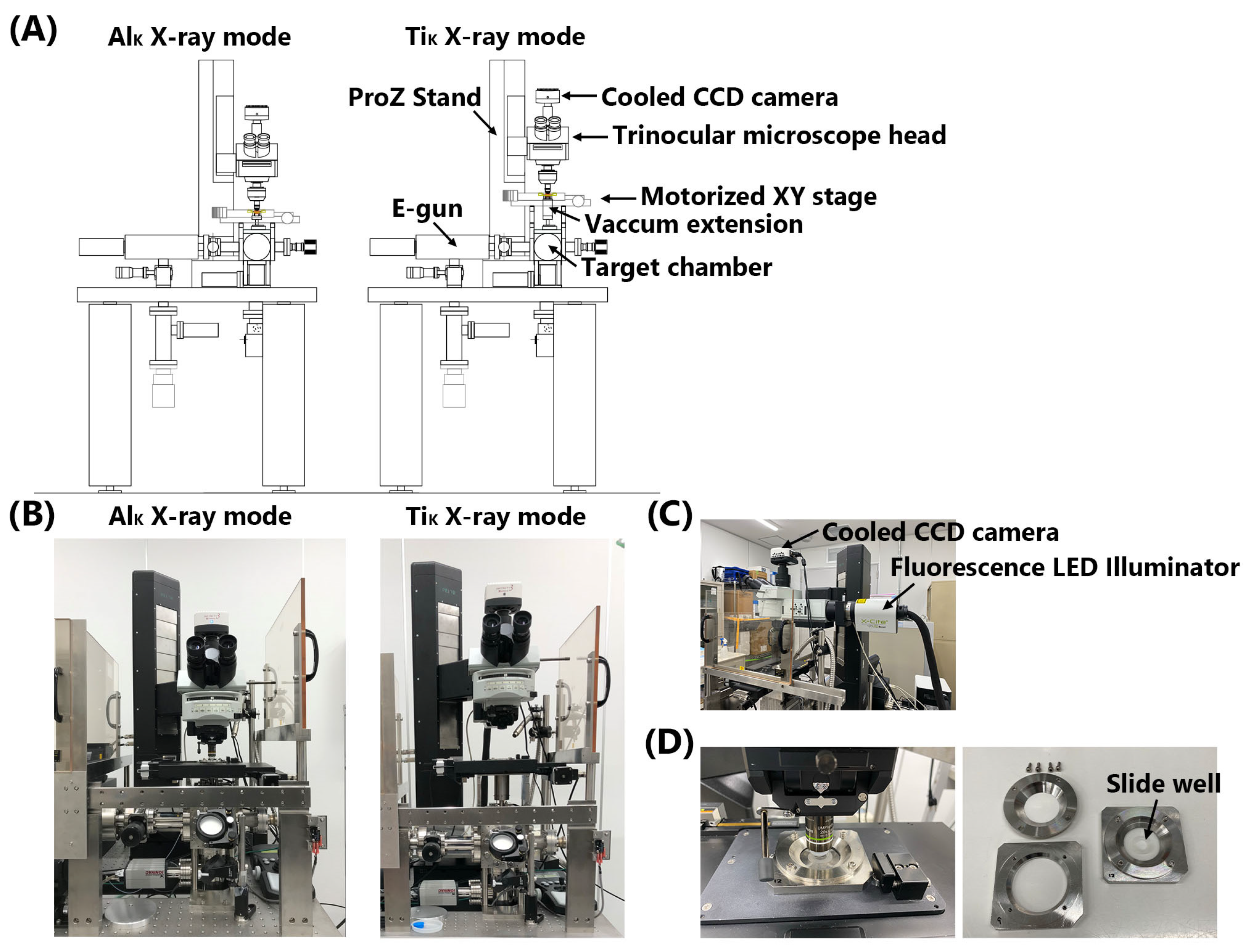
2.2.1. Microbeam X-ray Cell Irradiation System
2.2.2. Microbeam Irradiation
2.3. Immunofluorescence Microscopy
2.4. Statistics
3. Results
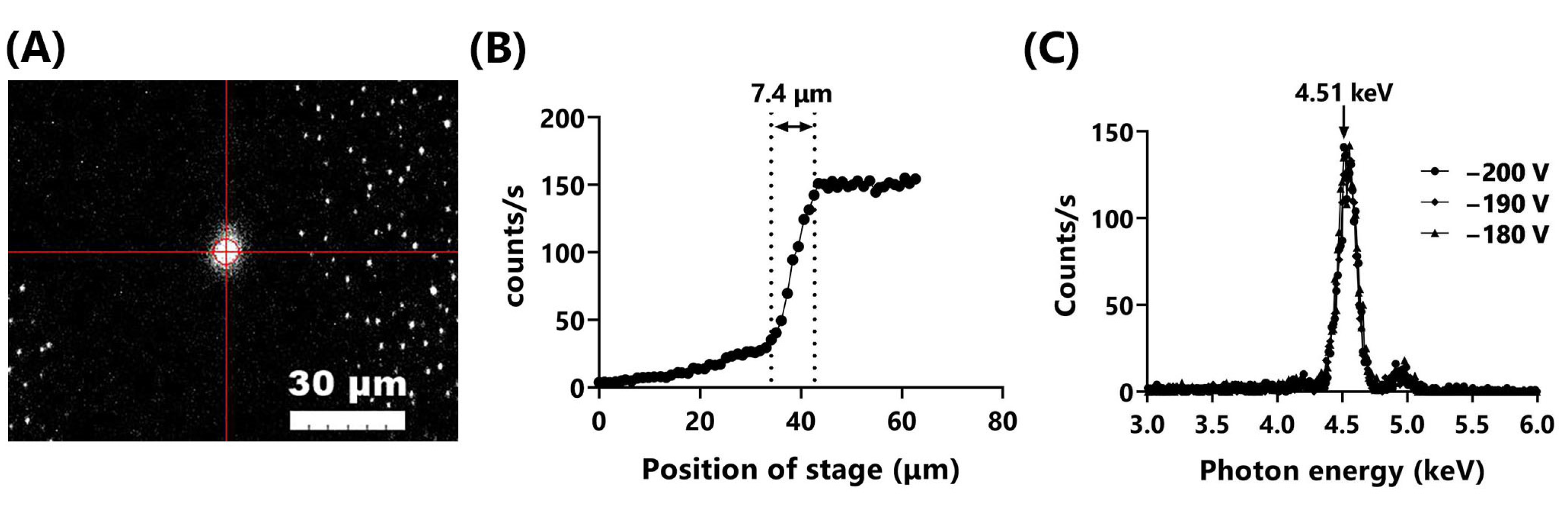
3.1. Upgrade of the Microbeam X-ray Cell Irradiation System
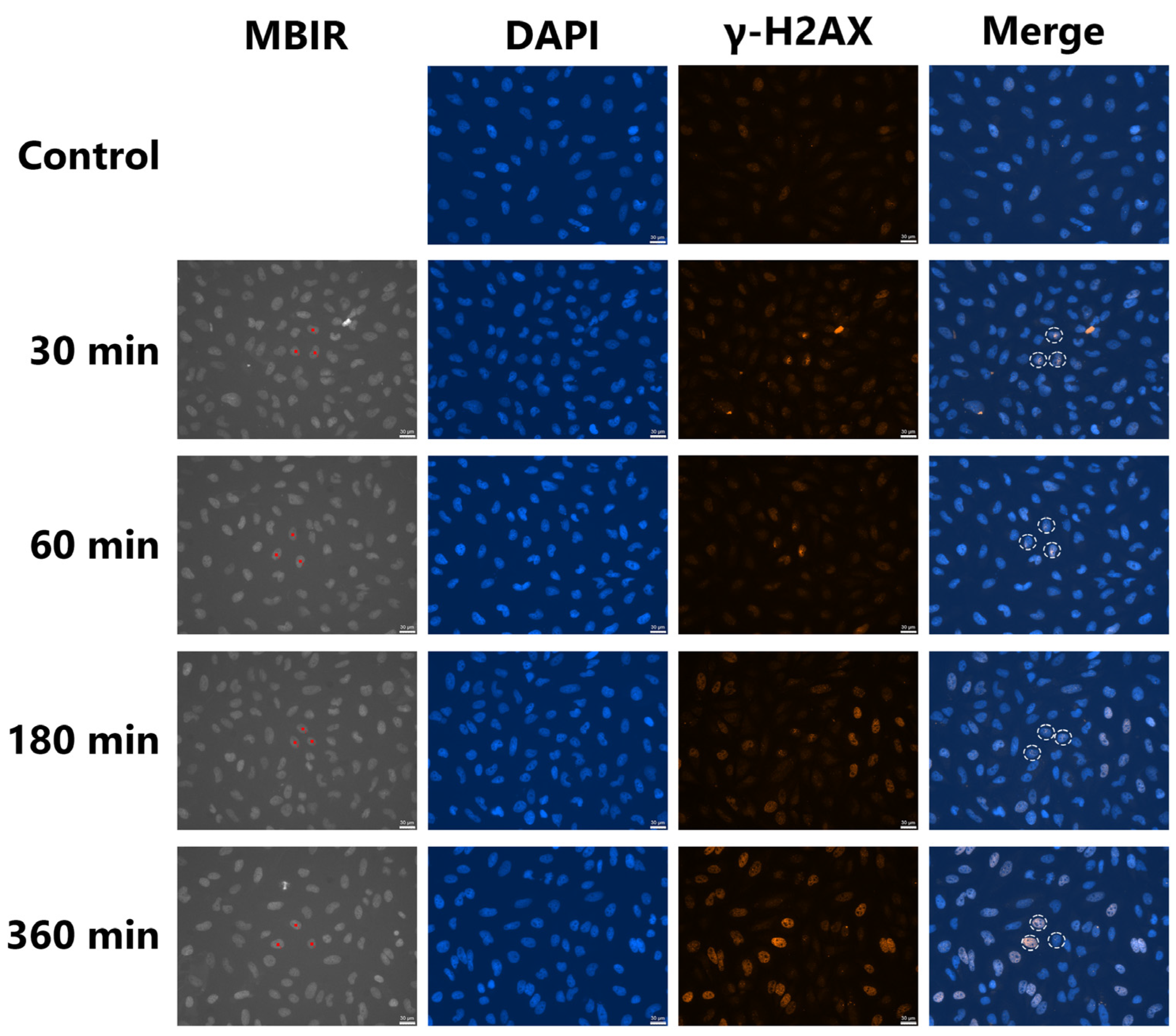
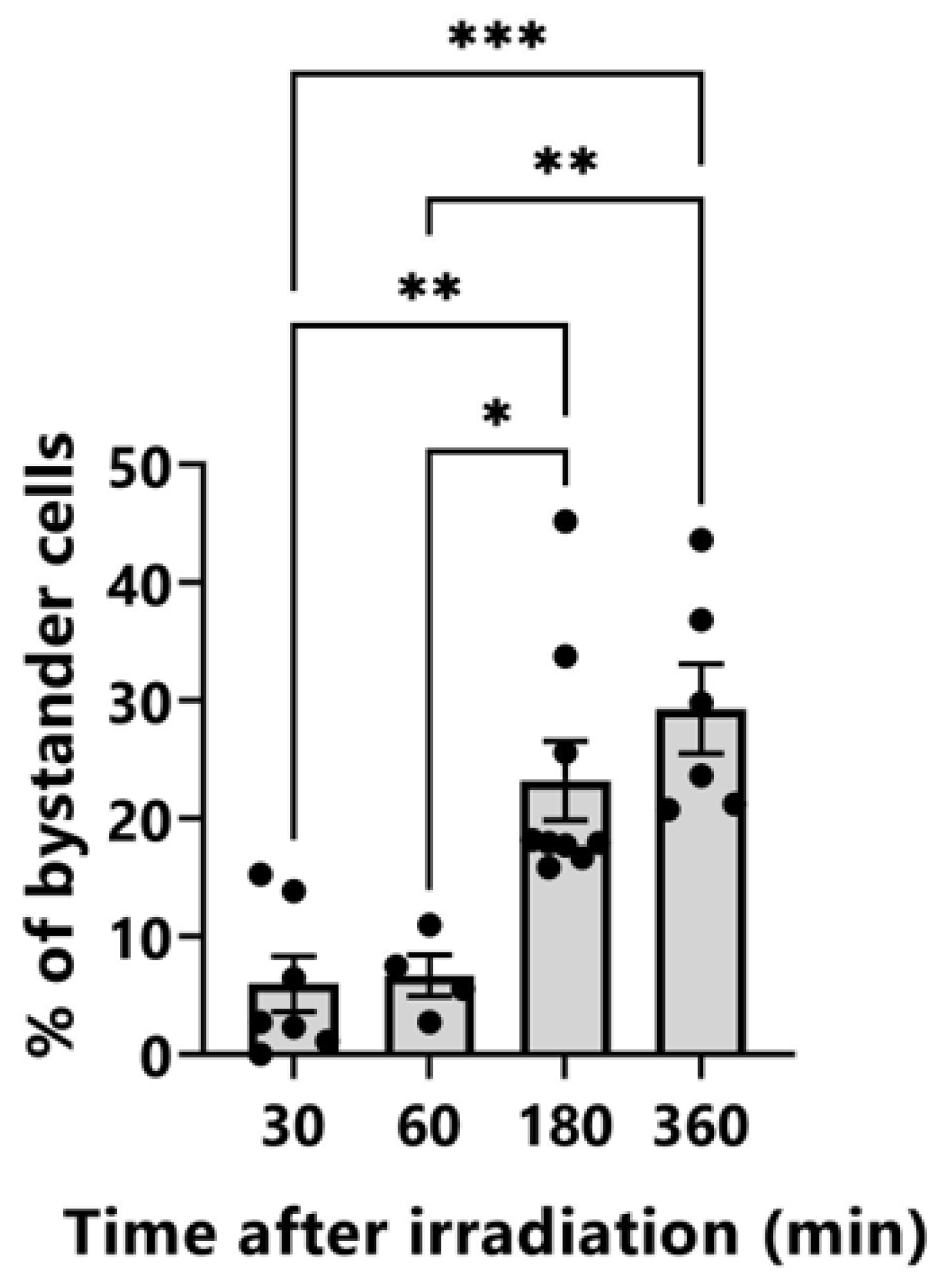
3.2. Induction of γ-H2AX-Positive Bystander Cells
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Tomita, M.; Maeda, M. Mechanisms and biological importance of photon-induced bystander responses: Do they have an impact on low-dose radiation responses. J. Radiat. Res. 2015, 56, 205–219. [Google Scholar] [CrossRef]
- Zhang, Z.; Li, K.; Hong, M. Radiation-induced bystander effect and cytoplasmic irradiation studies with microbeams. Biology 2022, 11, 945. [Google Scholar] [CrossRef] [PubMed]
- Nagasawa, H.; Little, J.B. Induction of sister chromatid exchanges by extremely low doses of α-particles. Cancer Res. 1992, 52, 6394–6396. [Google Scholar] [PubMed]
- Kobayashi, Y.; Funayama, T.; Hamada, N.; Sakashita, T.; Konishi, T.; Imaseki, H.; Yasuda, K.; Hatashita, M.; Takagi, K.; Hatori, S.; et al. Microbeam irradiation facilities for radiobiology in Japan and China. J. Radiat. Res. 2009, 50 (Suppl. A), A29–A47. [Google Scholar] [CrossRef]
- Wu, J.; Hei, T.K. Focus small to find big—The microbeam story. Int. J. Radiat. Biol. 2018, 94, 782–788. [Google Scholar] [CrossRef] [PubMed]
- Hei, T.K.; Zhou, H.; Chai, Y.; Ponnaiya, B.; Ivanov, V.N. Radiation induced non-targeted response: Mechanism and potential clinical implications. Curr. Mol. Pharmacol. 2011, 4, 96–105. [Google Scholar] [CrossRef]
- Tomita, M.; Matsumoto, H.; Funayama, T.; Yokota, Y.; Otsuka, K.; Maeda, M.; Kobayashi, Y. Nitric oxide-mediated bystander signal transduction induced by heavy-ion microbeam irradiation. Life Sci. Space Res. 2015, 6, 36–43. [Google Scholar] [CrossRef]
- Folkard, M.; Schettino, G.; Vojnovic, B.; Gilchrist, S.; Michette, A.G.; Pfauntsch, S.J.; Prise, K.M.; Michael, B.D. A focused ultrasoft X-ray microbeam for targeting cells individually with submicrometer accuracy. Radiat. Res. 2001, 156, 796–804. [Google Scholar] [CrossRef]
- Schettino, G.; Folkard, M.; Prise, K.M.; Vojnovic, B.; Held, K.D.; Michael, B.D. Low-dose studies of bystander cell killing with targeted soft X-rays. Radiat. Res. 2003, 160, 505–511. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, K.; Usami, N.; Hieda, K.; Takakura, K.; Maezawa, H.; Hayashi, T. Development of microbeam irradiation system for radiobiology. Nucl. Instrum. Methods Phys. Res. Sect. A 2001, 467–468, 1329–1332. [Google Scholar] [CrossRef]
- Kobayashi, K.; Usami, N.; Maezawa, H.; Hayashi, T.; Hieda, K.; Takakura, K. Synchrotron X-ray microbeam irradiation system for radiobiology. J. Biomed. Nanotechnol. 2006, 2, 116–119. [Google Scholar] [CrossRef]
- Tomita, M.; Kobayashi, K.; Maeda, M. Microbeam studies of soft X-ray induced bystander cell killing using microbeam X-ray cell irradiation system at CRIEPI. J. Radiat. Res. 2012, 53, 488. [Google Scholar] [CrossRef]
- Tomita, M.; Maeda, M.; Maezawa, H.; Usami, N.; Kobayashi, K. Bystander cell killing in normal human fibroblasts is induced by synchrotron X-ray microbeams. Radiat. Res. 2010, 173, 380–385. [Google Scholar] [CrossRef] [PubMed]
- Tomita, M.; Maeda, M.; Kobayashi, K.; Matsumoto, K. Dose response of soft X-ray-induced bystander cell killing affected by p53 status. Radiat. Res. 2013, 179, 200–207. [Google Scholar] [CrossRef]
- Levayer, R.; Moreno, E. Mechanisms of cell competition: Themes and variations. J. Cell Biol. 2013, 200, 689–698. [Google Scholar] [CrossRef] [PubMed]
- Gullikson, E. X-ray Database. Available online: https://henke.lbl.gov/optical_constants/ (accessed on 20 March 2023).
- Maeda, M.; Tomita, M.; Maeda, M.; Matsumoto, H.; Usami, N.; Kume, K.; Kobayashi, K. Exposure of the cytoplasm to low-dose X-rays modifies ataxia telangiectasia mutated-mediated DNA damage responses. Sci. Rep. 2021, 11, 13113. [Google Scholar] [CrossRef]
- Bodgi, L.; Foray, N. The nucleo-shuttling of the ATM protein as a basis for a novel theory of radiation response: Resolution of the linear-quadratic model. Int. J. Radiat. Biol. 2016, 92, 117–131. [Google Scholar] [CrossRef]
- Prise, K.M.; Folkard, M.; Michael, B.D. Radiation-induced bystander and adaptive responses in cell and tissue models. Dose Response 2006, 4, 263–276. [Google Scholar] [CrossRef] [PubMed]
- Sedelnikova, O.A.; Nakamura, A.; Kovalchuk, O.; Koturbash, I.; Mitchell, S.A.; Marino, S.A.; Brenner, D.J.; Bonner, W.M. DNA double-strand breaks form in bystander cells after microbeam irradiation of three-dimensional human tissue models. Cancer Res. 2007, 67, 4295–4302. [Google Scholar] [CrossRef]
- Kovalchuk, O.; Zemp, F.J.; Filkowski, J.N.; Altamirano, A.M.; Dickey, J.S.; Jenkins-Baker, G.; Marino, S.A.; Brenner, D.J.; Bonner, W.M.; Sedelnikova, O.A. microRNAome changes in bystander three-dimensional human tissue models suggest priming of apoptotic pathways. Carcinogenesis 2010, 31, 1882–1888. [Google Scholar] [CrossRef]
- Fujimichi, Y.; Otsuka, K.; Tomita, M.; Iwasaki, T. Intestinal organoids for studying the effects of low-dose/low-dose-rate radiation. Radiat. Prot. Dosimetry 2022, 198, 1115–1119. [Google Scholar] [CrossRef] [PubMed]
- Sokolov, M.V.; Smilenov, L.B.; Hall, E.J.; Panyutin, I.G.; Bonner, W.M.; Sedelnikova, O.A. Ionizing radiation induces DNA double-strand breaks in bystander primary human fibroblasts. Oncogene 2005, 24, 7257–7265. [Google Scholar] [CrossRef] [PubMed]
- Marti, T.M.; Hefner, E.; Feeney, L.; Natale, V.; Cleaver, J.E. H2AX phosphorylation within the G1 phase after UV irradiation depends on nucleotide excision repair and not DNA double-strand breaks. Proc. Natl. Acad. Sci. USA 2006, 103, 9891–9896. [Google Scholar] [CrossRef]
- Horn, S.; Brady, D.; Prise, K. Alpha particles induce pan-nuclear phosphorylation of H2AX in primary human lymphocytes mediated through ATM. Biochim. Biophys. Acta 2015, 1853, 2199–2206. [Google Scholar] [CrossRef]
- Meyer, B.; Voss, K.O.; Tobias, F.; Jakob, B.; Durante, M.; Taucher-Scholz, G. Clustered DNA damage induces pan-nuclear H2AX phosphorylation mediated by ATM and DNA-PK. Nucleic Acids Res. 2013, 41, 6109–6118. [Google Scholar] [CrossRef] [PubMed]
- Moeglin, E.; Desplancq, D.; Conic, S.; Oulad-Abdelghani, M.; Stoessel, A.; Chiper, M.; Vigneron, M.; Didier, P.; Tora, L.; Weiss, E. Uniform Widespread Nuclear Phosphorylation of Histone H2AX Is an Indicator of Lethal DNA Replication Stress. Cancers 2019, 11, 355. [Google Scholar] [CrossRef]
- Maeda, M.; Tomita, M.; Usami, N.; Kobayashi, K. Bystander cell death is modified by sites of energy deposition within cells irradiated with a synchrotron X-ray microbeam. Radiat. Res. 2010, 174, 37–45. [Google Scholar] [CrossRef]
- Maeda, M.; Kobayashi, K.; Matsumoto, H.; Usami, N.; Tomita, M. X-ray-induced bystander responses reduce spontaneous mutations in V79 cells. J. Radiat. Res. 2013, 54, 1043–1049. [Google Scholar] [CrossRef]
- Han, W.; Wu, L.; Chen, S.; Bao, L.; Zhang, L.; Jiang, E.; Zhao, Y.; Xu, A.; Hei, T.K.; Yu, Z. Constitutive nitric oxide acting as a possible intercellular signaling molecule in the initiation of radiation-induced DNA double strand breaks in non-irradiated bystander cells. Oncogene 2007, 26, 2330–2339. [Google Scholar] [CrossRef]
- Sokolov, M.V.; Dickey, J.S.; Bonner, W.M.; Sedelnikova, O.A. γ-H2AX in bystander cells: Not just a radiation-triggered event, a cellular response to stress mediated by intercellular communication. Cell Cycle 2007, 6, 2210–2212. [Google Scholar] [CrossRef]
- ICRP. ICRP Publication 131: Stem Cell Biology with Respect to Carcinogenesis Aspects of Radiological Protection. Ann. ICRP 2015, 44, 7–357. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| AlK X-rays | TiK X-ray | |
|---|---|---|
| Energy (keV) | 1.49 | 4.51 |
| Attenuation length (1/e) in water (μm) 1 | 7.1 | 171 |
| Beam size (μm) 2 | 1.8 | 7.4 |
| Dose rate at the sample position (Gy/min) 3 | 84 | 0.8 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tomita, M.; Torigata, M.; Ohchi, T.; Ito, A. Observation of Histone H2AX Phosphorylation by Radiation-Induced Bystander Response Using Titanium Characteristic X-ray Microbeam. Biology 2023, 12, 734. https://doi.org/10.3390/biology12050734
Tomita M, Torigata M, Ohchi T, Ito A. Observation of Histone H2AX Phosphorylation by Radiation-Induced Bystander Response Using Titanium Characteristic X-ray Microbeam. Biology. 2023; 12(5):734. https://doi.org/10.3390/biology12050734
Chicago/Turabian StyleTomita, Masanori, Masaya Torigata, Tadayuki Ohchi, and Atsushi Ito. 2023. "Observation of Histone H2AX Phosphorylation by Radiation-Induced Bystander Response Using Titanium Characteristic X-ray Microbeam" Biology 12, no. 5: 734. https://doi.org/10.3390/biology12050734
APA StyleTomita, M., Torigata, M., Ohchi, T., & Ito, A. (2023). Observation of Histone H2AX Phosphorylation by Radiation-Induced Bystander Response Using Titanium Characteristic X-ray Microbeam. Biology, 12(5), 734. https://doi.org/10.3390/biology12050734