
Feeding on the Fruit Waste Orange Bagasse Modifies Immature Protein Content, Body Weight, Scent Bouquet Composition, and Copula Duration in Males of a Tephritid Frugivorous Fly

 , ,
, ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Anastrepha ludens Artificial Diet-Rearing System
2.2. Anastrepha ludens Orange Bagasse Diet-Rearing System
2.3. Experimental Design
2.4. Experimental Procedures and Estimation of Variables
2.4.1. Number of Larvae Recovered per Rearing Tray
2.4.2. Pupation Percentage
2.4.3. Pupal Yield
2.4.4. Pupal Weight
2.4.5. Adult Emergence
2.4.6. Time to Emergence
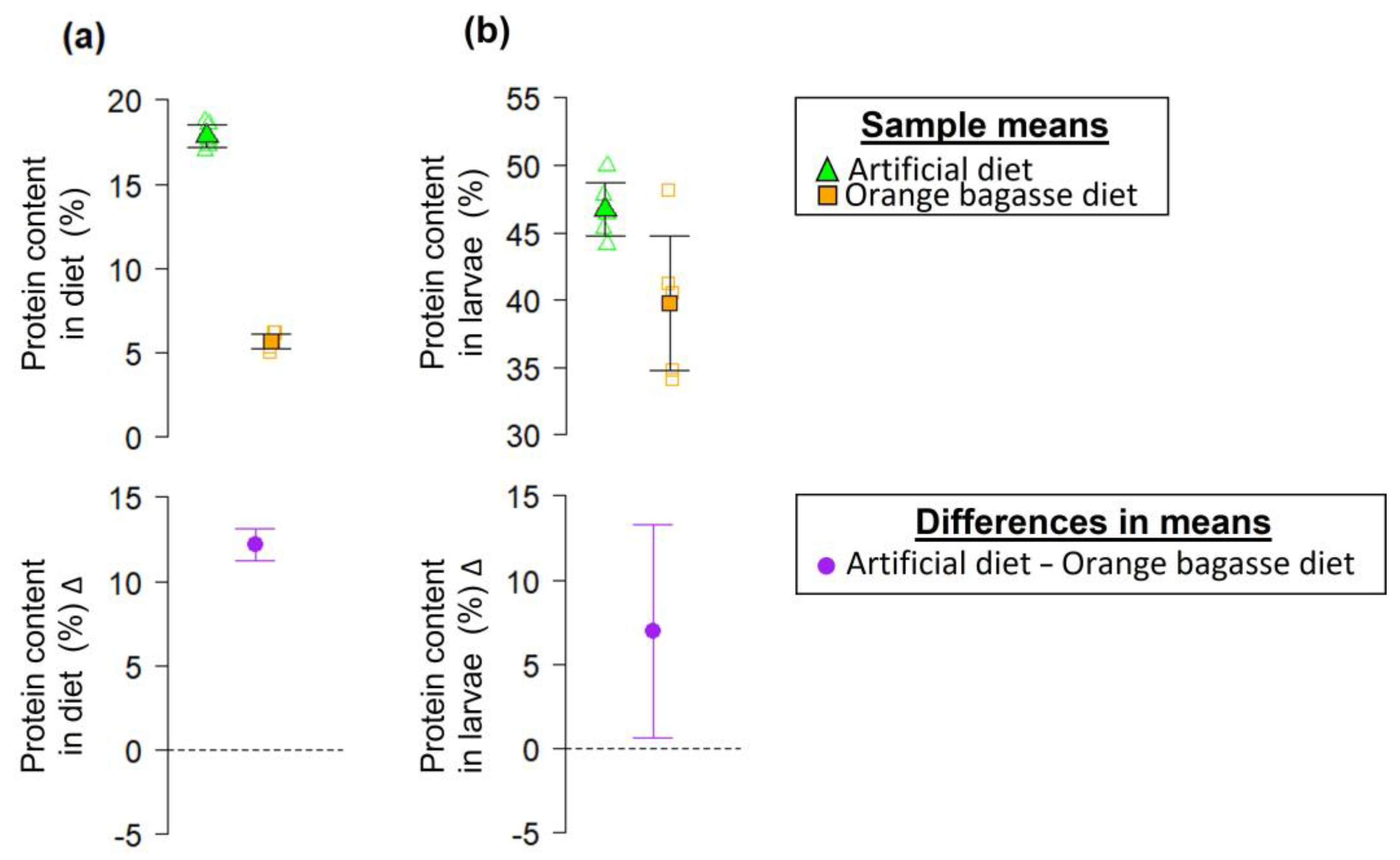
2.4.7. Protein Content in Larvae and Diets
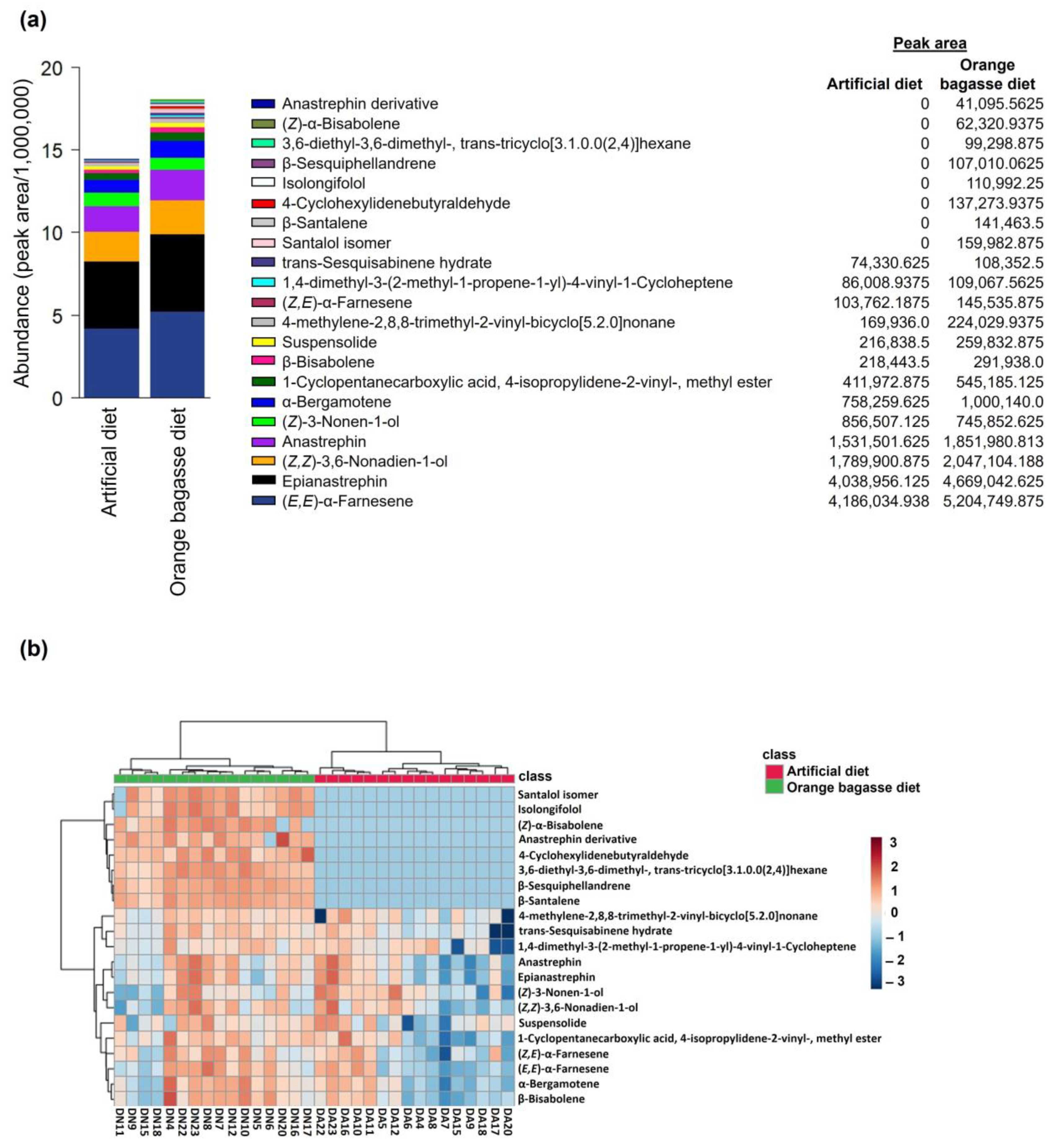
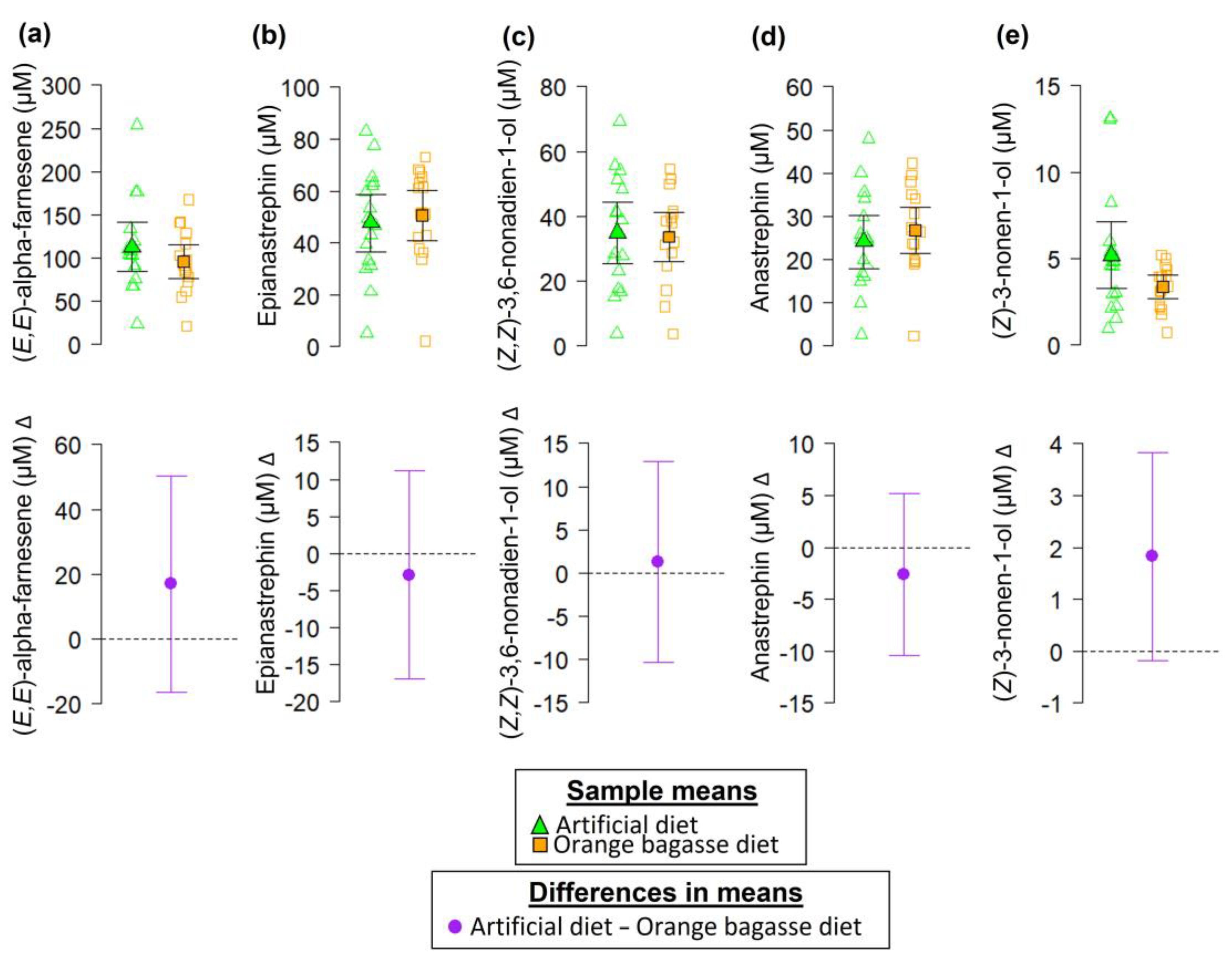
2.4.8. Chemical Compounds Identified in the Volatiles of Sexually Mature Male Flies
- Male Odor Collections
- Chemical Analyses of Male Odors
2.4.9. Sexual Competitiveness and Aspects of Sexual Behavior
- Fly Origin and Handling
- Marking of Males
- Laboratory Sexual Behavior Test
2.5. Statistical Analyses
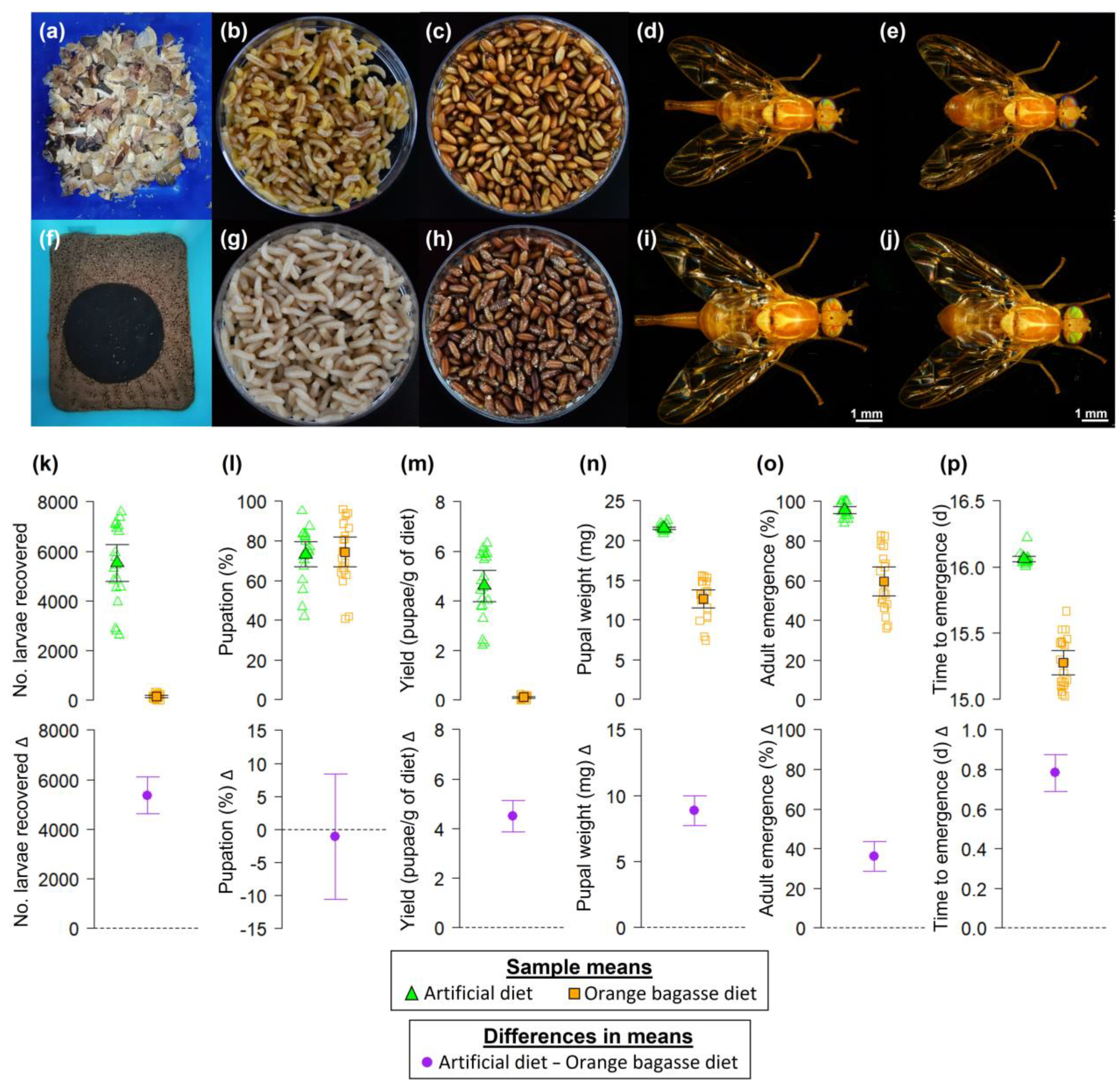
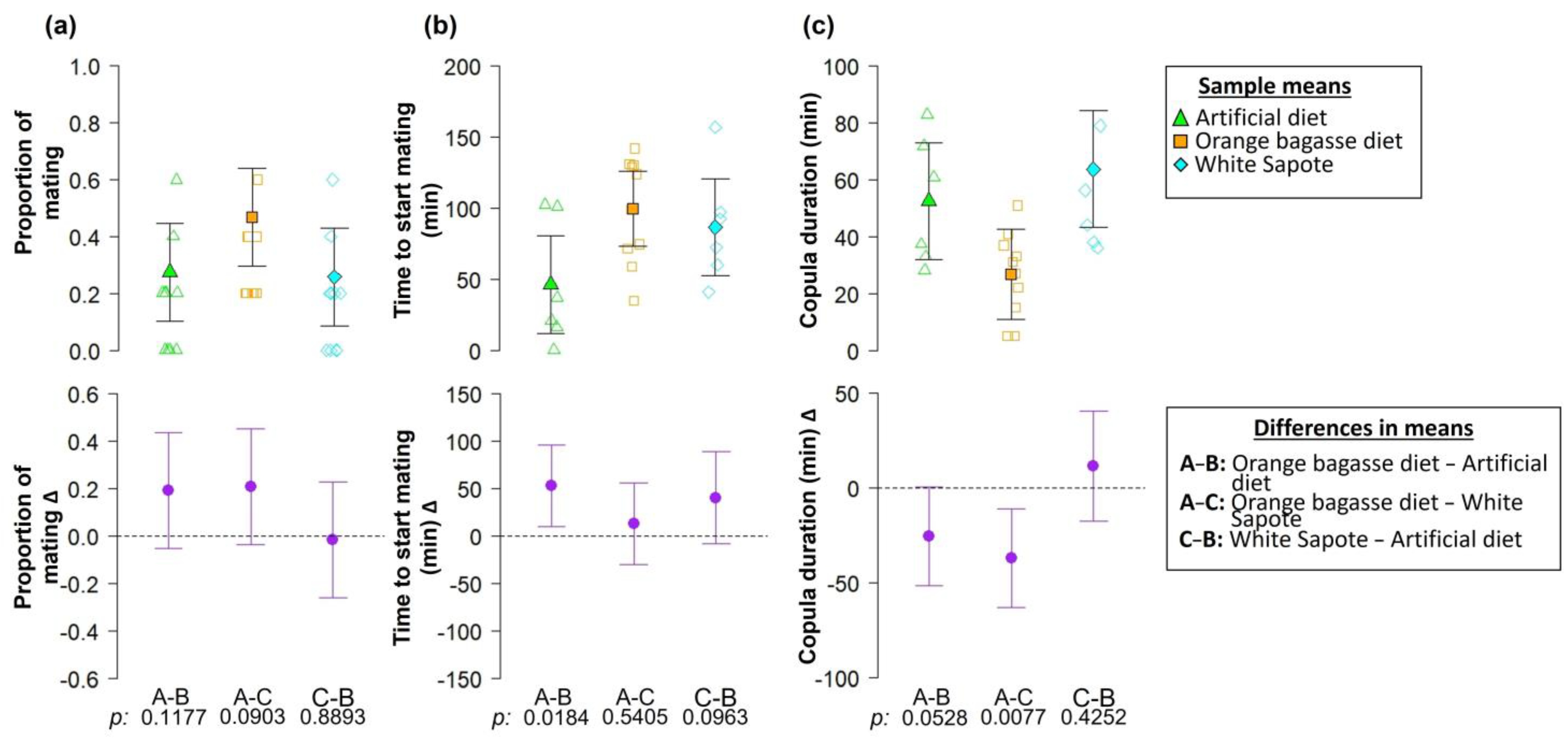
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Volume (mL) of Eggs Inoculated into 1 kg of Diet | Third Instar Larvae Recovered per g of Diet | Pupation (%) | Pupal Weight (mg) | Adult Emergence (%) | Egg-Adult Transformation (%) |
|---|---|---|---|---|---|
| 0.062 | 0.237 (0.081) | 92.4 (11.2) | 17.01 (1.15) | 93.3 (0.58) | 24.03 (9.24) |
| 0.125 | 0.413 (0.112) | 81.6 (1.6) | 14.78 (2.04) | 95.7 (0.58) | 18.85 (5.47) |
| 0.25 | 0.532 (0.252) | 88.7 (8.7) | 13.72 (2.46) | 96.7 (4.04) | 13.36 (6.66) |
References
- Díaz-Fleischer, F.F.; Papaj, D.R.; Prokopy, R.J.; Norrbom, A.L.; Aluja, M. Evolution of fruit fly oviposition behavior. In Fruit Flies (Tephritidae): Phylogeny and Evolution of Behavior; Aluja, M., Norrbom, A., Eds.; CRC Press: Boca Raton, FL, USA, 2000; pp. 811–841. [Google Scholar]
- Aluja, M.; Mangan, R.L. Fruit fly (Diptera: Tephritidae) host status determination: Critical conceptual, methodological, and regulatory considerations. Annu. Rev. Entomol. 2008, 53, 473–502. [Google Scholar] [CrossRef] [PubMed]
- Scolari, F.; Valerio, F.; Benelli, G.; Papadopoulos, N.T.; Vaníčková, L. Tephritid fruit fly semiochemicals: Current knowledge and future perspectives. Insects 2021, 12, 408. [Google Scholar] [CrossRef] [PubMed]
- Aluja, M. Manejo Integrado de la Mosca de la Fruta; Trillas: Mexico City, Mexico, 1993. [Google Scholar]
- Aluja, M.; Piñero, J.I.; Jácome, F.; Díaz, F.; Sivinski, J. Behavior of flies in the genus Anastrepha (Trypetinae: Toxotrypanini). In Fruit Flies (Tephritidae): Phylogeny and Evolution of Behavior; Aluja, M., Norrbom, A., Eds.; CRC Press: Boca Raton, FL, USA, 2000; pp. 375–406. [Google Scholar]
- Birke, A.; Guillén, L.; Midgarden, D.; Aluja, M. Fruit flies, Anastrepha ludens (Loew), A. obliqua (Macquart) and A. grandis (Macquart) (Diptera: Tephritidae): Three pestiferous tropical fruit flies that could potentially expand their range to temperate areas. In Emerging Invasive Pests of Agricultural Crops; Peña, J.E., Wysoki, M., Eds.; CABI International: Boca Raton, FL, USA, 2013; pp. 192–213. [Google Scholar]
- Drew, R.A.; Yuval, B. The evolution of fruit fly feeding behavior. In Fruit Flies (Tephritidae): Phylogeny and Evolution of Behavior; Aluja, M., Norrbom, A., Eds.; CRC Press: Boca Raton, FL, USA, 2000; pp. 731–749. [Google Scholar]
- Aluja, M. Bionomics and management of Anastrepha. Annu. Rev. Entomol. 1994, 39, 155–178. [Google Scholar] [CrossRef]
- Birke, A.; Aluja, M. Do mothers really know best? Complexities in testing the preference-performance hypothesis in polyphagous frugivorous fruit flies. Bull. Entomol. Res. 2018, 108, 674–684. [Google Scholar] [CrossRef] [PubMed]
- Merli, D.; Mannucci, B.; Bassetti, F.; Corana, F.; Falchetto, M.; Malacrida, A.R.; Scolari, F. Larval diet affects male pheromone blend in a laboratory strain of the Medfly, Ceratitis capitata (Diptera: Tephritidae). J. Chem. Ecol. 2018, 44, 339–353. [Google Scholar] [CrossRef]
- Nguyen, B.; Ponton, F.; Than, A.; Taylor, P.W.; Chapman, T.; Morimoto, J. Interactions between ecological factors in the developmental environment modulate pupal and adult traits in a polyphagous fly. Ecol. Evol. 2019, 9, 6342–6352. [Google Scholar] [CrossRef]
- Aluja, M.; Cabagne, G.; Altúzar-Molina, A.; Pascacio-Villafán, C.; Enciso, E.; Guillén, L. Host plant and antibiotic effects on scent bouquet composition of Anastrepha ludens and Anastrepha obliqua calling males, two polyphagous tephritid pests. Insects 2020, 11, 309. [Google Scholar] [CrossRef]
- Morimoto, J.; Than, A.T.; Nguyen, B.; Lundbäck, I.; Dinh, H.; Ponton, F. Density-by-Diet interactions during larval development shape adult life history trait expression and fitness in a polyphagous fly. Am. Nat. 2022, 199, E170–E185. [Google Scholar] [CrossRef]
- Pascacio-Villafán, C.; Righini, N.; Nestel, D.; Birke, A.; Guillén, L.; Aluja, M. Diet quality and conspecific larval density predict functional trait variation and performance in a polyphagous frugivorous fly. Funct. Ecol. 2022, 36, 1163–1176. [Google Scholar] [CrossRef]
- González-López, G.I.; Orozco-Dávila, D.; López-Hernández, P.; Díaz-Fleischer, F.; Alvarado-Castillo, G.; Sánchez-Viveros, G.; Pérez-Staples, D. Effect of limited protein consumption during the larval and adult stage on life—History traits of the Mexican fruit fly. Entomol. Exp. Appl. 2022, 170, 922–932. [Google Scholar] [CrossRef]
- Chen, E.H.; Hou, Q.L.; Wei, D.D.; Jiang, H.B.; Wang, J.J. Phenotypic plasticity, trade-offs and gene expression changes accompanying dietary restriction and switches in Bactrocera dorsalis (Hendel) (Diptera: Tephritidae). Sci. Rep. 2017, 7, 1988. [Google Scholar] [CrossRef] [PubMed]
- Cohen, A.C. Ecology of insect rearing systems: A mini-review of insect rearing papers from 1906–2017. Adv. Entomol. 2018, 6, 86. [Google Scholar] [CrossRef]
- Francuski, L.; Beukeboom, L.W. Insects in production–an introduction. Entomol. Exp. Appl. 2020, 168, 422–431. [Google Scholar] [CrossRef]
- Huynh, M.P.; Shelby, K.S.; Coudron, T.A. Recent advances in insect rearing methodology to promote scientific research and mass production. Insects 2021, 12, 961. [Google Scholar] [CrossRef]
- Bakri, A.; Mehta, K.; Lance, D.R. Sterilizing insects with ionizing radiation. In Sterile Insect Technique. Principles and Practice in Area-Wide Integrated Pest Management, 2nd ed.; Dyck, V.A., Hendrichs, J., Robinson, A.S., Eds.; CRC Press: Boca Raton, FL, USA, 2021; pp. 355–398. [Google Scholar]
- Parker, A.G.; Mamai, W.; Maiga, H. Mass-rearing for the sterile insect technique. In Sterile Insect Technique. Principles and Practice in Area-Wide Integrated Pest Management, 2nd ed.; Dyck, V.A., Hendrichs, J., Robinson, A.S., Eds.; CRC Press: Boca Raton, FL, USA, 2021; pp. 283–316. [Google Scholar]
- Ekesi, S.; Mohammed, S. Mass rearing and quality control parameters for Tephritid fruit flies of economic importance in Africa. In Wide Spectra of Quality Control, 1st ed.; Akyar, I., Ed.; IntechOpen: London, UK, 2011; pp. 387–410. [Google Scholar]
- Cohen, A.C. Design, Operation, and Control of Insect Rearing Systems; CRC Press: Boca Raton, FL, USA, 2021; pp. 1–420. [Google Scholar]
- Maset, B.A.; Demetrio, C.G.B.; Lopes, L.A.; Costa, M.D.L.Z.; Botteon, V.W.; Mastrangelo, T.D.A. Which artificial larval diet is better for Ceratitis capitata (Diptera: Tephritidae) rearing? J. Basic Appl. Zool. 2022, 83, 48. [Google Scholar] [CrossRef]
- Aceituno-Medina, M.; Rincón-Betancurt, O.; Martínez-Salgado, R.T.; Hernández, E. A novel, low-cost coconut fiber larval diet for mass rearing Anastrepha (Diptera: Tephritidae). J. Econ. Entomol. 2019, 112, 1112–1119. [Google Scholar] [CrossRef]
- Vargas, R.; Chang, H.; Williamson, D.L. Evaluation of a sugarcane bagasse larval diet for mass production of the Mediterranean Fruit Fly (Diptera: Tephritidae) in Hawaii. J. Econ. Entomol. 1983, 76, 1360–1362. [Google Scholar] [CrossRef]
- Adhikari, P.; Aryal, N.; Ghimire, A.; Khanal, P. Sustainable biowaste recycling using insects. In Clean Energy and Resources Recovery; Tyagi, V., Aboudi, K., Eds.; Elsevier: Amsterdam, The Netherlands, 2021; Volume 1, pp. 399–420. [Google Scholar]
- Cadinu, L.A.; Barra, P.; Torre, F.; Delogu, F.; Madau, F.A. Insect rearing: Potential, challenges, and circularity. Sustainability 2020, 12, 4567. [Google Scholar] [CrossRef]
- Aluja, M.; Sivinski, J.; Ovruski, S.; Guillen, L.; Lopez, M.; Cancino, J.; Ruíz, L. Colonization and domestication of seven species of native New World hymenopterous larval-prepupal and pupal fruit fly (Diptera: Tephritidae) parasitoids. Biocontrol Sci. Technol. 2009, 19, 49–79. [Google Scholar] [CrossRef]
- Choi, I.S.; Lee, Y.G.; Khanal, S.K.; Park, B.J.; Bae, H.J. A low-energy, cost-effective approach to fruit and citrus peel waste processing for bioethanol production. Appl. Energy 2015, 140, 65–74. [Google Scholar] [CrossRef]
- Al-Saadi, N.H.M.; Ahmad, N.S.; Sa’eed, S.E. Determination of some chemical compounds and the effect of oil extract from orange peel on some pathogens. J. Kerbala Univ. 2009, 7, 2. [Google Scholar]
- Debernardi-Vázquez, T.J.; Aguilar-Rivera, N. Manejo sostenible de subproductos agroindustriales y estrategias tecnológicas de bioconversión. Rev. Colomb. Investig. Agroind. 2020, 7, 22–31. [Google Scholar] [CrossRef]
- Aluja, M.; Ordano, M.; Teal, P.E.; Sivinski, J.; García-Medel, D.; Anzures-Dadda, A. Larval feeding substrate and species significantly influence the effect of a juvenile hormone analog on sexual development/performance in four tropical tephritid flies. J. Insect Physiol. 2009, 55, 231–242. [Google Scholar] [CrossRef] [PubMed]
- Pascacio-Villafán, C.; Guillén, L.; Williams, T.; Aluja, M. Effects of larval density and support substrate in liquid diet on productivity and quality of artificially reared Anastrepha ludens (Diptera: Tephritidae). J. Econ. Entomol. 2018, 111, 2281–2287. [Google Scholar] [CrossRef]
- Storey, R.; Treeby, T. The morphology of epicuticular wax and albedo cells of orange fruit in relation to albedo breakdown. J. Hortic. Sci. Biotechnol. 1994, 69, 329–338. [Google Scholar] [CrossRef]
- Birke, A.; Aluja, M.; Greany, P.; Bigurra, E.; Pérez-Staples, D.; McDonald, R. Long aculeus and behavior of Anastrepha ludens render gibberellic acid ineffective as an agent to reduce ‘Ruby Red’ grapefruit susceptibility to the attack of this pestiferous fruit fly in commercial groves. J. Econ. Entomol. 2006, 99, 1184–1193. [Google Scholar] [CrossRef]
- FAO/IAEA/USDA. Product Quality Control for Sterile Mass-Reared and Released Tephritid Fruit Flies; Version 6.0; International Atomic Energy Agency: Vienna, Austria, 2014. [Google Scholar]
- Pascacio-Villafán, C.; Guillén, L.; Aluja, M. Agar and carrageenan as cost-effective gelling agents in yeast-reduced artificial diets for mass-rearing fruit flies and their parasitoids. Insects 2020, 11, 131. [Google Scholar] [CrossRef]
- Chang, S.K.C.; Zhang, Y. Protein analysis. In Food Analysis, 5th ed.; Nielsen, S.S., Ed.; Food Science Text Series; Springer International Publishing: Cham, Switzerland, 2017; pp. 315–331. [Google Scholar]
- Heath, R.R.; Manukian, A. Development and evaluation of systems to collect volatiles semiochemicals from insects and plants using a charcoal-infused medium for air purification. J. Chem. Ecol. 1992, 18, 1209–1226. [Google Scholar] [CrossRef]
- Aluja, M.; Jácome, I.; Macías, O.R. Effect of adult nutrition on male sexual performance in four Neotropical fruit fly species of the genus Anastrepha (Diptera: Tephritidae). J. Insect Behav. 2001, 14, 759–775. [Google Scholar] [CrossRef]
- Krzywinski, M.; Altman, N. Points of view: Designing comparative experiments. Nat. Methods 2014, 11, 597–598. [Google Scholar] [CrossRef]
- Metaboanalyst 5.0 Bioinformatic Platform. Available online: https://www.metaboanalyst.ca/home.xhtml (accessed on 15 January 2023).
- Dushoff, J.; Kain, M.P.; Bolker, B.M. I can see clearly now: Reinterpreting statistical significance. Methods Ecol. Evol. 2019, 10, 756–759. [Google Scholar] [CrossRef]
- Muff, S.; Nilsen, E.B.; O’Hara, R.B.; Nater, C.R. Rewriting results sections in the language of evidence. Trends Ecol. Evol. 2022, 37, 203–210. [Google Scholar] [CrossRef] [PubMed]
- R Development Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2017. [Google Scholar]
- RStudio Team. RStudio: Integrated Development Environment for R; RStudio, Inc.: Boston, MA, USA, 2021. [Google Scholar]
- Nash, W.J.; Chapman, T. Effect of dietary components on larval life history characteristics in the Medfly (Ceratitis capitata: Diptera, Tephritidae). PLoS ONE 2014, 9, e86029. [Google Scholar] [CrossRef]
- M’hiri, N.; Ioannou, I.; Mihoubi Boudhrioua, N.; Ghoul, M. Effect of different operating conditions on the extraction of phenolic compounds in orange peel. Food Bioprod. Process. 2015, 96, 161–170. [Google Scholar] [CrossRef]
- Ayala, J.R.; Montero, G.; Coronado, M.A.; García, C.; Curiel-Alvarez, M.A.; León, J.A.; Sagaste, C.A.; Montes, D.G. Characterization of orange peel waste and valorization to obtain reducing sugars. Molecules 2021, 26, 1348. [Google Scholar] [CrossRef] [PubMed]
- Abdelazem, R.E.; Hefnawy, H.T.; El-Shorbagy, G.A. Chemical composition and phytochemical screening of Citrus sinensis (orange) peels. Zagazig J. Agric. Res. 2021, 48, 793–804. [Google Scholar] [CrossRef]
- Healthline. Available online: https://www.healthline.com/nutrition/can-you-eat-orange-peels (accessed on 14 February 2023).
- Favela-Hernández, J.M.J.; González-Santiago, O.; Ramírez-Cabrera, M.A.; Esquivel-Ferriño, P.C.; Camacho-Corona, M.D.R. Chemistry and pharmacology of Citrus sinensis. Molecules 2016, 21, 247. [Google Scholar] [CrossRef]
- Hoffmann, A.A.; Ross, P.A. Rates and patterns of laboratory adaptation in (mostly) insects. J. Econ. Entomol. 2018, 111, 501–509. [Google Scholar] [CrossRef]
- Aluja, M.; Zamora-Briseño, J.A.; Pérez-Brocal, V.; Altúzar-Molina, A.; Guillén, L.; Desgarennes, D.; Vásquez-Rosas, M.; Ibarra-Laclette, E.; Alonso-Sánchez, A.G.; Moya, A. Metagenomic survey of the highly polyphagous Anastrepha ludens developing in ancestral and exotic hosts reveals the lack of a stable microbiota in larvae and the strong influence of metamorphosis on adult gut microbiota. Front. Microbiol. 2021, 12, 685937. [Google Scholar] [CrossRef]
- Drew, R.A.I. Amino acid increases in fruit infested by fruit flies of the family Tephritidae. Zool. J. Linn. Soc. 1988, 93, 107–112. [Google Scholar] [CrossRef]
- Deutscher, A.T.; Chapman, T.A.; Shuttleworth, L.A.; Riegler, M.; Reynolds, O.L. Tephritid-microbial interactions to enhance fruit fly performance in sterile insect technique programs. BMC Microbiol. 2019, 19, 287. [Google Scholar] [CrossRef] [PubMed]
- Behar, A.; Yuval, B.; Jurkevitch, E. Enterobacteria—mediated nitrogen fixation in natural populations of the fruit fly Ceratitis capitata. Mol. Ecol. 2005, 14, 2637–2643. [Google Scholar] [CrossRef]
- Behar, A.; Jurkevitch, E.; Yuval, B. Bringing back the fruit into fruit fly–bacteria interactions. Mol. Ecol. 2008, 17, 1375–1386. [Google Scholar] [CrossRef]
- Morimoto, J. Addressing global challenges with unconventional insect ecosystem services: Why should humanity care about insect larvae? People Nat. 2020, 3, 582–595. [Google Scholar] [CrossRef]
- Pérez-Staples, D.; Abraham, S. Postcopulatory behavior of tephritid flies. Annu. Rev. Entomol. 2023, 68, 89–108. [Google Scholar] [CrossRef] [PubMed]
- Bosa, C.F.; Cruz-López, L.; Zepeda-Cisneros, C.S.; Valle-Mora, J.; Guillén-Navarro, K.; Liedo, P. Sexual behavior and male volatile compounds in wild and mass—Reared strains of the Mexican fruit fly Anastrepha ludens (Diptera: Tephritidae) held under different colony management regimes. Insect Sci. 2016, 23, 105–116. [Google Scholar] [CrossRef] [PubMed]
- Russell, W.M.S.; Burch, R.L. The Principles of Humane Experimental Technique, 1st ed.; Methuen: London, UK, 1959; pp. 1–238. [Google Scholar]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pascacio-Villafán, C.; Guillén, L.; Altúzar-Molina, A.; Tellez-Mora, J.A.; Cruz-Hernández, E.; Aluja, M. Feeding on the Fruit Waste Orange Bagasse Modifies Immature Protein Content, Body Weight, Scent Bouquet Composition, and Copula Duration in Males of a Tephritid Frugivorous Fly. Biology 2023, 12, 739. https://doi.org/10.3390/biology12050739
Pascacio-Villafán C, Guillén L, Altúzar-Molina A, Tellez-Mora JA, Cruz-Hernández E, Aluja M. Feeding on the Fruit Waste Orange Bagasse Modifies Immature Protein Content, Body Weight, Scent Bouquet Composition, and Copula Duration in Males of a Tephritid Frugivorous Fly. Biology. 2023; 12(5):739. https://doi.org/10.3390/biology12050739
Chicago/Turabian StylePascacio-Villafán, Carlos, Larissa Guillén, Alma Altúzar-Molina, Julio A. Tellez-Mora, Enedina Cruz-Hernández, and Martín Aluja. 2023. "Feeding on the Fruit Waste Orange Bagasse Modifies Immature Protein Content, Body Weight, Scent Bouquet Composition, and Copula Duration in Males of a Tephritid Frugivorous Fly" Biology 12, no. 5: 739. https://doi.org/10.3390/biology12050739
APA StylePascacio-Villafán, C., Guillén, L., Altúzar-Molina, A., Tellez-Mora, J. A., Cruz-Hernández, E., & Aluja, M. (2023). Feeding on the Fruit Waste Orange Bagasse Modifies Immature Protein Content, Body Weight, Scent Bouquet Composition, and Copula Duration in Males of a Tephritid Frugivorous Fly. Biology, 12(5), 739. https://doi.org/10.3390/biology12050739