
Genetic Analysis as a Tool to Improve the Monitoring of Stranded Cetaceans in Chile

 , , and
, , and
Abstract
:Simple Summary
Abstract
1. Introduction
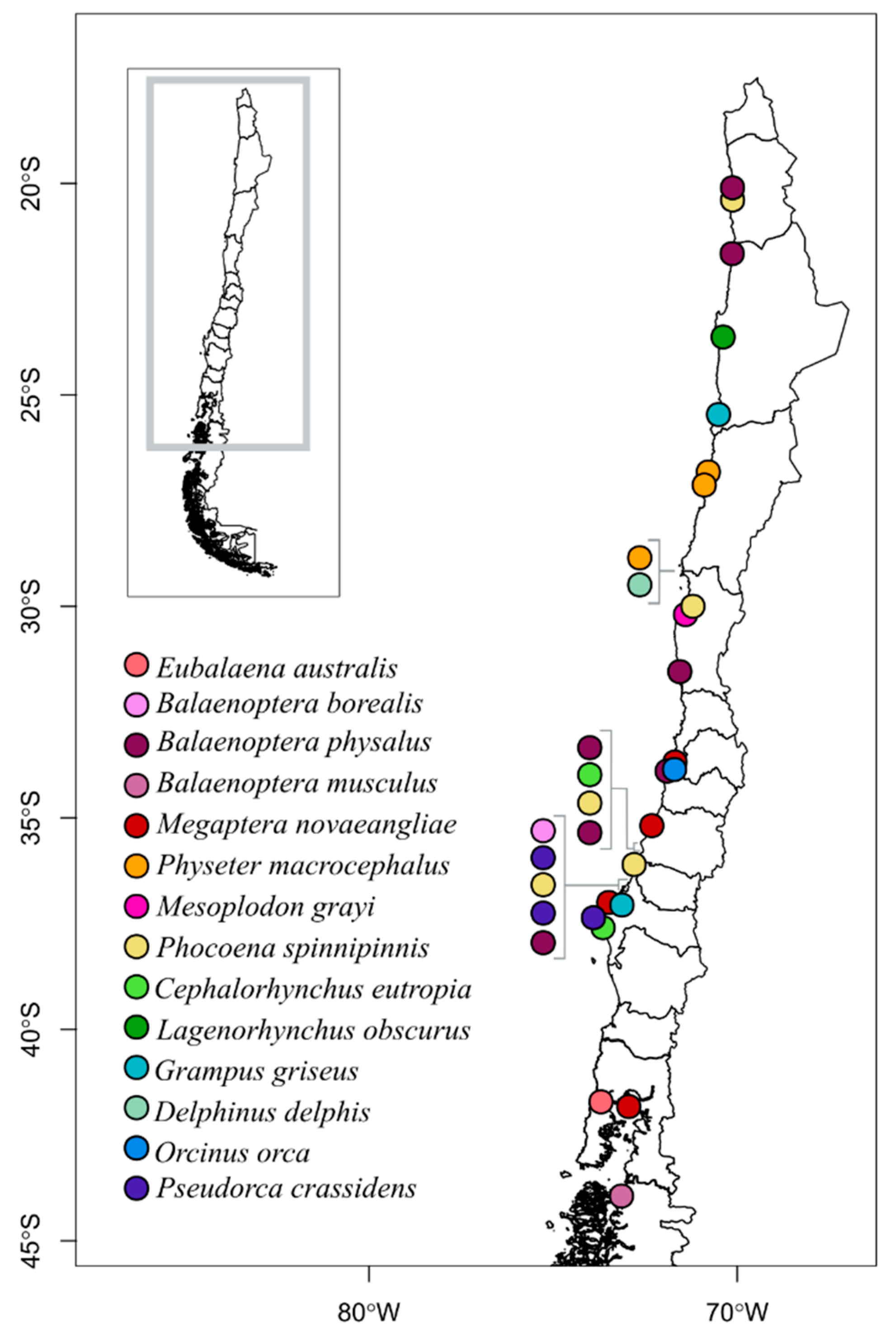
2. Materials and Methods
2.1. Collection of Samples
2.2. Species Identification
2.3. Sex Identification
3. Results
4. Discussion
4.1. Species Identification
4.2. Sex Identification
4.3. Multidisciplinary Approach and Inter-Institutional Collaboration
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Moore, K.M.; Simeone, C.A.; Brownell, R.L., Jr. Strandings. In Encyclopedia of Marine Mammals; Würsig, B.G., Thewissen, J.G.M., Kovacs, K.M., Eds.; Academic Press, An Imprint of Elsevier: London, UK; San Diego, CA, USA, 2018; pp. 945–951. ISBN 978-0-12-804327-1. [Google Scholar]
- Geraci, J.R.; St. Aubin, D.J. Mass Stranding of the Long-Finned Pilot Whale, Globicephala melaena, on Sable Island, Nova Scotia. J. Fish. Res. Board Can. 1977, 34, 2193–2196. [Google Scholar] [CrossRef]
- Bilgmann, K.; Möller, L.M.; Harcourt, R.G.; Kemper, C.M.; Beheregaray, L.B. The Use of Carcasses for the Analysis of Cetacean Population Genetic Structure: A Comparative Study in Two Dolphin Species. PLoS ONE 2011, 6, e20103. [Google Scholar] [CrossRef]
- Costa, A.F.; Siciliano, S.; Emin-Lima, R.; Martins, B.M.L.; Sousa, M.E.M.; Giarrizzo, T.; Silva Júnior, J.D.S.E. Stranding Survey as a Framework to Investigate Rare Cetacean Records of the North and North-Eastern Brazilian Coasts. ZooKeys 2017, 688, 111–134. [Google Scholar] [CrossRef]
- Thompson, K.F.; Millar, C.D.; Scott Baker, C.; Dalebout, M.; Steel, D.; van Helden, A.L.; Constantine, R. A Novel Conservation Approach Provides Insights into the Management of Rare Cetaceans. Biol. Conserv. 2013, 157, 331–340. [Google Scholar] [CrossRef]
- Shotwell, M.; McFee, W.; Slate, E.H. Estimating Gompertz Growth Curves from Marine Mammal Strandings in the Presence of Missing Data. Int. J. Ecol. Econ. Stat. 2010, 19, 32–46. [Google Scholar] [PubMed]
- Turner, J.P.; Clark, L.S.; Haubold, E.M.; Worthy, G.A.J.; Cowan, D.F. Organ Weights and Growth Profiles in Bottlenose Dolphins (Tursiops truncatus) from the Northwestern Gulf of Mexico. Aquat. Mamm. 2006, 32, 46–57. [Google Scholar] [CrossRef]
- McLellan, W.A.; Friedlaender, A.S.; Mead, J.G.; Potter, C.W.; Pabst, D.A. Analysing 25 Years of Bottlenose Dolphin (Tursiops truncatus) Strandings along the Atlantic Coast of the USA: Do Historic Records Support the Coastal Migratory Stock Hypothesis? J. Cetacean Res. Manag. 2002, 4, 297–304. [Google Scholar]
- Alvarado-Rybak, M.; Toro, F.; Escobar-Dodero, J.; Kinsley, A.C.; Sepúlveda, M.A.; Capella, J.; Azat, C.; Cortés-Hinojosa, G.; Zimin-Veselkoff, N.; Mardones, F.O. 50 Years of Cetacean Strandings Reveal a Concerning Rise in Chilean Patagonia. Sci. Rep. 2020, 10, 9511. [Google Scholar] [CrossRef]
- Geraci, J.R.; St. Aubin, D.J. Stranding Workshop Summary Report. Analysis of Marine Mammal Strandings and Recommendations for a Nationwide Stranding Salvage Program. In Biology of Marine Mammals: Insights through Strandings; Final Report to U.S.; Geraci, J.R., St. Aubin, D.J., Eds.; Marine Mammal Commission: Washington, DC, USA, 1979; pp. 1–33. [Google Scholar]
- Wilkinson, D.; Worthy, A.J. Marine Mammal Stranding Networks. In Conservation and Management of Marine Mammals; Twiss, J.R., Reeves, R.R., Eds.; Smithsonian Institution Press: Washington, DC, USA, 1999; pp. 396–411. [Google Scholar]
- Dalebout, M.L.; Baker, C.S.; Mead, J.G.; Cockcroft, V.G.; Yamada, T.K. A Comprehensive and Validated Molecular Taxonomy of Beaked Whales, Family Ziphiidae. J. Hered. 2004, 95, 459–473. [Google Scholar] [CrossRef]
- Kraft, S.; Pérez-Álvarez, M.; Olavarría, C.; Poulin, E. Global Phylogeography and Genetic Diversity of the Long-Finned Pilot Whale Globicephala Melas, with New Data from the Southeastern Pacific. Sci. Rep. 2020, 10, 1769. [Google Scholar] [CrossRef]
- Oremus, M.; Gales, R.; Kettles, H.; Baker, C.S. Genetic Evidence of Multiple Matrilines and Spatial Disruption of Kinship Bonds in Mass Strandings of Long-Finned Pilot Whales, Globicephala melas. J. Hered. 2013, 104, 301–311. [Google Scholar] [CrossRef]
- Oremus, M.; Gales, R.; Dalebout, M.L.; Funahashi, N.; Endo, T.; Kage, T.; Steel, D.; Baker, S.C. Worldwide Mitochondrial DNA Diversity and Phylogeography of Pilot Whales (Globicephala Spp.): WORLDWIDE MtDNA OF PILOT WHALES. Biol. J. Linn. Soc. 2009, 98, 729–744. [Google Scholar] [CrossRef]
- Heyning, J.E. Collecting and Archiving of Cetacean Data and Specimens. NOAA Tech. Rep. NMFS 1991, 98, 69–74. [Google Scholar]
- Sheldrick, M.C. Trends in the Strandings of Cetacea on the British Coasts 1913-72. Mammal Rev. 1976, 6, 15–23. [Google Scholar] [CrossRef]
- Brabyn, M.W. An Analysis of the New Zealand Whale Stranding Record. Sci. Res. Ser. 1991, 29, 1–45. [Google Scholar]
- van der Meij, S.E.T.; Camphuysen, C.J. Distribution and Diversity of Whales and Dolphins (Cetacea) in the Southern North Sea: 1970–2005. Lutra 2006, 49, 3–28. [Google Scholar]
- Li, T.; Wu, H.; Wu, C.; Yang, G.; Chen, B. Molecular Identification of Stranded Cetaceans in Coastal China. Aquat. Mamm. 2019, 45, 525–532. [Google Scholar] [CrossRef]
- Constantine, R.; Carroll, E.; Stewart, R.; Neale, D.; van Helden, A. First Record of True’s Beaked Whale Mesoplodon mirus in New Zealand. Mar. Biodivers. Rec. 2014, 7, e1. [Google Scholar] [CrossRef]
- Sabatier, E.; Pante, E.; Dussud, C.; Van Canneyt, O.; Simon-Bouhet, B.; Viricel, A. Genetic Monitoring of Pilot Whales, Globicephala spp. (Cetacea: Delphinidae), Stranded on French Coasts. Mammalia 2014, 79, 111–114. [Google Scholar] [CrossRef]
- Alfonsi, E.; Méheust, E.; Fuchs, S.; Carpentier, F.-G.; Quillivic, Y.; Viricel, A.; Hassani, S.; Jung, J.-L. The Use of DNA Barcoding to Monitor the Marine Mammal Biodiversity along the French Atlantic Coast. Zookeys 2013, 365, 5–24. [Google Scholar] [CrossRef]
- Fioravanti, T.; Maio, N.; Latini, L.; Splendiani, A.; Guarino, F.M.; Mezzasalma, M.; Petraccioli, A.; Cozzi, B.; Mazzariol, S.; Centelleghe, C.; et al. Nothing Is as It Seems: Genetic Analyses on Stranded Fin Whales Unveil the Presence of a Fin-Blue Whale Hybrid in the Mediterranean Sea (Balaenopteridae). Eur. Zool. J. 2022, 89, 590–600. [Google Scholar] [CrossRef]
- Häussermann, V.; Gutstein, C.S.; Beddington, M.; Cassis, D.; Olavarria, C.; Dale, A.C.; Valenzuela-Toro, A.M.; Perez-Alvarez, M.J.; Sepúlveda, H.H.; McConnell, K.M.; et al. Largest Baleen Whale Mass Mortality during Strong El Niño Event Is Likely Related to Harmful Toxic Algal Bloom. PeerJ 2017, 5, e3123. [Google Scholar] [CrossRef]
- Pérez-Álvarez, M.J.; Rodríguez, F.; Kraft, S.; Segovia, N.; Olavarría, C.; Baker, C.S.; Steel, D.; Funahashi, N.; Häussermann, V.; Ulloa, M.; et al. Phylogeography and Demographic Inference of the Endangered Sei Whale, with Implications for Conservation. Aquat. Conserv. 2021, 31, 3554–3563. [Google Scholar] [CrossRef]
- Haro, D.; Aguayo-Lobo, A.; Blank, O.; Cifuentes, C.; Dougnac, C.; Arredondo, C.; Pardo, C.; Cáceres-Saez, I. Nuevo Varamiento Masivo de Orca Falsa, Pseudorca crassidens, En El Estrecho de Magallanes, Chile. Rev. Biol. Mar. Oceanogr. 2015, 50, 149–155. [Google Scholar] [CrossRef]
- Alvarado-Rybak, M.; Haro, D.; Oyarzún, P.A.; Dougnac, C.; Gutierrez, J.; Toledo, N.; Leiva, N.; Peña, C.; Cifuentes, C.; Muñoz, N.; et al. A Mass Stranding Event of Long-Finned Pilot Whales (Globicephala melas) in Southern Chile. Aquat. Mamm. 2019, 45, 447–455. [Google Scholar] [CrossRef]
- Aljanabi, S.M.; Martinez, I. Universal and Rapid Salt-Extraction of High Quality Genomic DNA for PCR-Based Techniques. Nucleic Acids Res. 1997, 25, 4692–4693. [Google Scholar] [CrossRef]
- Siemann, L.A. Mitochondrial DNA Sequence Variation in North Atlantic Pilot Whales, Globicephala Melas. Ph.D. Thesis, Massachusetts Institute of Technology, Woods Hole Oceanographic Institution, Woods Hole, MA, USA, 1994. [Google Scholar]
- Ross, H.A.; Lento, G.M.; Dalebout, M.L.; Goode, M.; Ewing, G.; McLaren, P.; Rodrigo, A.G.; Lavery, S.; Baker, C.S. DNA Surveillance: Web-Based Molecular Identification of Whales, Dolphins, and Porpoises. J. Hered. 2003, 94, 111–114. [Google Scholar] [CrossRef]
- Dalebout, M.L.; Robertson, K.M.; Frantzis, A.; Engelhaupt, D.; Mignucci-Giannoni, A.A.; Rosario-Delestre, R.J.; Baker, C.S. Worldwide Structure of MtDNA Diversity among Cuvier’s Beaked Whales (Ziphius Cavirostris): Implications for Threatened Populations. Mol. Ecol. 2005, 14, 3353–3371. [Google Scholar] [CrossRef]
- Filatov, D.A. Processing and Population Genetic Analysis of Multigenic Datasets with ProSeq3 Software. Bioinformatics 2009, 25, 3189–3190. [Google Scholar] [CrossRef]
- Aasen, E.; Medrano, J.F. Amplification of the ZFY and ZFX Genes for Sex Identification in Humans, Cattle, Sheep and Goats. Bio/technology 1990, 8, 1279–1281. [Google Scholar] [CrossRef]
- Gilson, A.; Syvanen, M.; Levine, K.; Banks, J. Deer Gender Determination by Polymerase Chain Reaction: Validation Study and Application to Tissues, Bloodstains, and Hair Forensic Samples from California. Calif. Fish Game 1998, 84, 159–169. [Google Scholar]
- IUCN. Balaenoptera Physalus: Cooke, J.G. In The IUCN Red List of Threatened Species 2018: E.T2478A50349982 2018; IUCN: Gland, Switzerland, 2018. [Google Scholar]
- MINSEGPRES. Decreto Supremo N° 23. Aprueba y Oficializa Nómina Para El Cuarto Proceso de Clasificación de Especies Según Su Estado de Conservación; Ministerio de Secretaría General de La Presidencia de La República: Santiago de Chile, Chile, 2009. [Google Scholar]
- García-Cegarra, A.M.; Pacheco, A.S. Collision Risk Areas between Fin and Humpback Whales with Large Cargo Vessels in Mejillones Bay (23°S), Northern Chile. Mar. Policy 2019, 103, 182–186. [Google Scholar] [CrossRef]
- Toro, F.; Gutiérrez, J.; Alvarado-Rybak, M.; Henríquez, A.; Henríquez, J.; Follador, J.; Abarca, P.; Calderón, C.; Peña, C.; Aravena, P.; et al. Report of Two Fin Whale (Balaenoptera Physalus) Stranding Associated with Ship Strike in Central-South Coast of Chile. Lat. Am. J. Aquat. Mamm. 2020, 15, 8–14. [Google Scholar]
- IUCN. Mesoplodon Grayi: Pitman, R.L. & Taylor, B.L. In The IUCN Red List of Threatened Species 2020: E.T13247A50366236 2020; IUCN: Gland, Switzerland, 2020. [Google Scholar]
- Goodall, R.N.P.; Norris, K.S.; Galeazzi, A.R.; Oporto, J.A.; Cameron, I.S. On the Chilean Dolphin, Cephalorhynchus Eutropia (Gray 1846). In Reports of the International Whaling Commission Special Issue 9; International Whaling Commission: Cambridge, UK, 1988; pp. 197–257. [Google Scholar]
- Heinrich, S. Ecology of Chilean Dolphins and Peale’s Dolphins at Isla Chiloe, Southern Chile. Ph.D. Thesis, University of St Andrews, St. Andrews, UK, 2006. [Google Scholar]
- Reyes, J.C.; Oporto, J.A. Gillnet Fisheries and Cetaceans in the Southeast Pacific. In Reports of the International Whaling Commission Special Issue 15; International Whaling Commission: Cambridge, UK, 1994; pp. 467–474. [Google Scholar]
- Pérez-Alvarez, M.J.; Olavarría, C.; Moraga, R.; Baker, C.S.; Hamner, R.M.; Poulin, E. Microsatellite Markers Reveal Strong Genetic Structure in the Endemic Chilean Dolphin. PLoS ONE 2015, 10, e0123956. [Google Scholar] [CrossRef]
- Heinrich, S.; Reeves, R. Cephalorhynchus Eutropia; International Union for Conservation of Nature (IUCN), Ed.; Red List of Threatened Species; International Union for Conservation of Nature: Gland, Switzerland, 2017. [Google Scholar]
- Brownell, R.L., Jr.; Clapham, P.J. Burmeister’s Porpoise Phocoena Spinipinnis Burmeister, 1865. In Handbook of Marine Mammals, Volume 6: The Second Book of Dolphins and the Porpoises; Ridgway, S.H., Harrison, R., Eds.; Academic Press: London, UK, 1999; pp. 393–410. [Google Scholar]
- IUCN. Phocoena Spinipinnis; Félix, F., Alfaro, J., Reyes, J., Mangel, J., Dellabianca, N., Heinrich, S., Crespo, E., Eds.; The IUCN Red List of Threatened Species 2018: E.T17029A50370481 2018; International Union for Conservation of Nature: Gland, Switzerland, 2018. [Google Scholar]
- Weir, C.R.; Rutherford, S. First Record of Burmeister’s Porpoise (Phocoena Spinipinnis) in the Falkland Islands (Malvinas). Mar. Biodivers. Rec. 2019, 12, 19. [Google Scholar] [CrossRef]
- Goodall, R.N.P.; Würsig, B.; Würsig, M.; Harris, G.; Norris, K.S. Sightings of Burmeister’s Porpoise, Phocoena Spinipinnis, off Southern South America. In Annual Reports of the International Whaling Commission; International Whaling Commission: Cambridge, UK, 1995; pp. 297–316. [Google Scholar]
- Rosa, S.; Milinkovitch, M.C.; Van Waerebeek, K.; Berck, J.; Oporto, J.; Alfaro-Shigueto, J.; Van Bressem, M.F.; Goodall, N.; Cassens, I. Population Structure of Nuclear and Mitochondrial DNA Variation among South American Burmeister’s Porpoises (Phocoena Spinipinnis). Conserv. Genet. 2005, 6, 431–443. [Google Scholar] [CrossRef]
- Silva, V.S.; Skueresky, N.; Lopes, F.; Koch, T.K.; Ott, P.H.; Siciliano, S.; Barreto, A.S.; Secchi, E.R.; de Meirelles, A.C.O.; Carvalho, V.L.; et al. Integrating Morphology and DNA Barcoding to Assess Cetacean Diversity in Brazil. Mamm. Res. 2021, 66, 349–369. [Google Scholar] [CrossRef]
- Galletti Vernazzani, B.; Cabrera, E.; Brownell, R.L. Eastern South Pacific Southern Right Whale Photo-Identification Catalog Reveals Behavior and Habitat Use Patterns. Mar. Mammal Sci. 2014, 30, 389–398. [Google Scholar] [CrossRef]
- IUCN. Eubalaena Australis (Chile-Peru Subpopulation): Cooke, J.G. In The IUCN Red List of Threatened Species 2018: E.T133704A50385137 2017; International Union for Conservation of Nature: Gland, Switzerland, 2017. [Google Scholar]
- Carroll, E.L.; Ott, P.H.; McMillan, L.F.; Galletti Vernazzani, B.; Neveceralova, P.; Vermeulen, E.; Gaggiotti, O.E.; Andriolo, A.; Baker, C.S.; Bamford, C.; et al. Genetic Diversity and Connectivity of Southern Right Whales (Eubalaena Australis) Found in the Brazil and Chile–Peru Wintering Grounds and the South Georgia (Islas Georgias Del Sur) Feeding Ground. J. Hered. 2020, 111, 263–276. [Google Scholar] [CrossRef]
- Aguayo-Lobo, A.; Torres Navarro, D.; Acevedo Ramírez, J. Los Mamíferos Marinos de Chile: I. Cetacea. In Serie Científica INACH 47; Instituto Antartico Chileno: Punta Arenas, Chile, 1988. [Google Scholar]
- Dudhat, S.; Pande, A.; Nair, A.; Mondal, I.; Srinivasan, M.; Sivakumar, K. Spatio-Temporal Analysis Identifies Marine Mammal Stranding Hotspots along the Indian Coastline. Sci. Rep. 2022, 12, 4128. [Google Scholar] [CrossRef]
- Cabrera, A.A.; Bérubé, M.; Lopes, X.M.; Louis, M.; Oosting, T.; Rey-Iglesia, A.; Rivera-León, V.E.; Székely, D.; Lorenzen, E.D.; Palsbøll, P.J. A Genetic Perspective on Cetacean Evolution. Annu. Rev. Ecol. Evol. Syst. 2021, 52, 131–151. [Google Scholar] [CrossRef]
- Kraft, S. Varamiento de Calderones de Aleta Larga En La Patagonia Chilena: Contribución a La Estructura Genética, Social y a La Filogeografía de La Especie. Master’s Thesis, Universidad de Chile, Santiago, Chile, 2019. [Google Scholar]
- Rodríguez, F. Diversidad Genética y Fileogeográfica de La Ballena Sei (Balaenoptera Borealis). Caracterización Genética Del Evento de Mortalidad Masiva de Rorcuales Ocurrida En Golfo de Penas, Aysén, Chile. Master’s Thesis, Universidad de Chile, Santiago, Chile, 2020. [Google Scholar]
- Haro, D.; Riccialdelli, L.; Blank, O.; Matus, R.; Sabat, P. Estimating the Isotopic Niche of Males and Females of False Killer Whales (Pseudorca Crassidens) from Magellan Strait, Chile. Mar. Mammal Sci. 2019, 35, 1070–1082. [Google Scholar] [CrossRef]
- Hansen, A.M.K.; Bryan, C.E.; West, K.; Jensen, B.A. Trace Element Concentrations in Liver of 16 Species of Cetaceans Stranded on Pacific Islands from 1997 through 2013. Arch. Environ. Contam. Toxicol. 2016, 70, 75–95. [Google Scholar] [CrossRef]
- Farwig, N.; Ammer, C.; Annighöfer, P.; Baur, B.; Behringer, D.; Diekötter, T.; Hotes, S.; Leyer, I.; Müller, J.; Peter, F.; et al. Bridging Science and Practice in Conservation: Deficits and Challenges from a Research Perspective. Basic Appl. Ecol. 2017, 24, 1–8. [Google Scholar] [CrossRef]
- Gazni, A.; Sugimoto, C.R.; Didegah, F. Mapping World Scientific Collaboration: Authors, Institutions, and Countries. J. Am. Soc. Inf. Sci. 2012, 63, 323–335. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Family | Species Identification | Sex Identification | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Species (molecular ID) | n° | Match | New ID | Mis ID | Match | New ID | Mis ID | Indet | |
| Balaenidae (n = 1) | Eubalaena australis | 1 | 1 | 1M | |||||
| Balaenopteridae (n = 17) | Balaenoptera physalus | 8 | 4 | 2 | 2 | 1F, 4M | 1F, 1M | 1M | |
| Balaenoptera bonaerensis | 1 | 1 | 1M | ||||||
| Balaenoptera borealis | 3 | 1 | 2 | 1M, 1F | 1 | ||||
| Balaenoptera musculus | 1 | 1 | 1 | ||||||
| Megaptera novaeangliae | 4 | 4 | 1M | 1F, 1M | 1M | ||||
| Delphinidae (n = 11) | Cephalorhynchus eutropia | 2 | 2 | 1F | 1F | ||||
| Delphinus delphis | 1 | 1 | 1F | ||||||
| Grampus griseus | 2 | 2 | 1M | 1F | |||||
| Lagenorhynchus australis | 1 | 1 | 1M | ||||||
| Lagenorhynchus obscurus | 1 | 1 | 1M | ||||||
| Orcinus orca | 1 | 1 | 1M | ||||||
| Pseudorca crassidens | 3 | 3 | 2F | 1M | |||||
| Phocoenidae (n = 5) | Phocoena spinipinnis | 5 | 4 | 1 | 1F | 2F, 1M | 1 | ||
| Physeteridae (n = 3) | Physeter macrocephalus | 3 | 3 | 1F | 1F | 1 | |||
| Ziphiidae (n = 2) | Mesoplodon grayi | 1 | 1 | 1F | |||||
| Mesoplodon layardii | 1 | 1 | 1M | ||||||
| Total | 39 | 29 | 7 | 3 | 14 | 17 | 4 | 4 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kraft, S.; Rodríguez, F.; Olavarría, C.; Poulin, E.; Pérez-Álvarez, M.J. Genetic Analysis as a Tool to Improve the Monitoring of Stranded Cetaceans in Chile. Biology 2023, 12, 748. https://doi.org/10.3390/biology12050748
Kraft S, Rodríguez F, Olavarría C, Poulin E, Pérez-Álvarez MJ. Genetic Analysis as a Tool to Improve the Monitoring of Stranded Cetaceans in Chile. Biology. 2023; 12(5):748. https://doi.org/10.3390/biology12050748
Chicago/Turabian StyleKraft, Sebastián, Francisca Rodríguez, Carlos Olavarría, Elie Poulin, and María José Pérez-Álvarez. 2023. "Genetic Analysis as a Tool to Improve the Monitoring of Stranded Cetaceans in Chile" Biology 12, no. 5: 748. https://doi.org/10.3390/biology12050748
APA StyleKraft, S., Rodríguez, F., Olavarría, C., Poulin, E., & Pérez-Álvarez, M. J. (2023). Genetic Analysis as a Tool to Improve the Monitoring of Stranded Cetaceans in Chile. Biology, 12(5), 748. https://doi.org/10.3390/biology12050748