
Alzheimer’s Disease and Age-Related Changes in the Cu Isotopic Composition of Blood Plasma and Brain Tissues of the APPNL-G-F Murine Model Revealed by Multi-Collector ICP-Mass Spectrometry

 and
and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Reagents
2.2. Samples
2.3. Sample Preparation
2.4. Instrumentation and Measurements
2.5. Statistical Analysis
3. Results
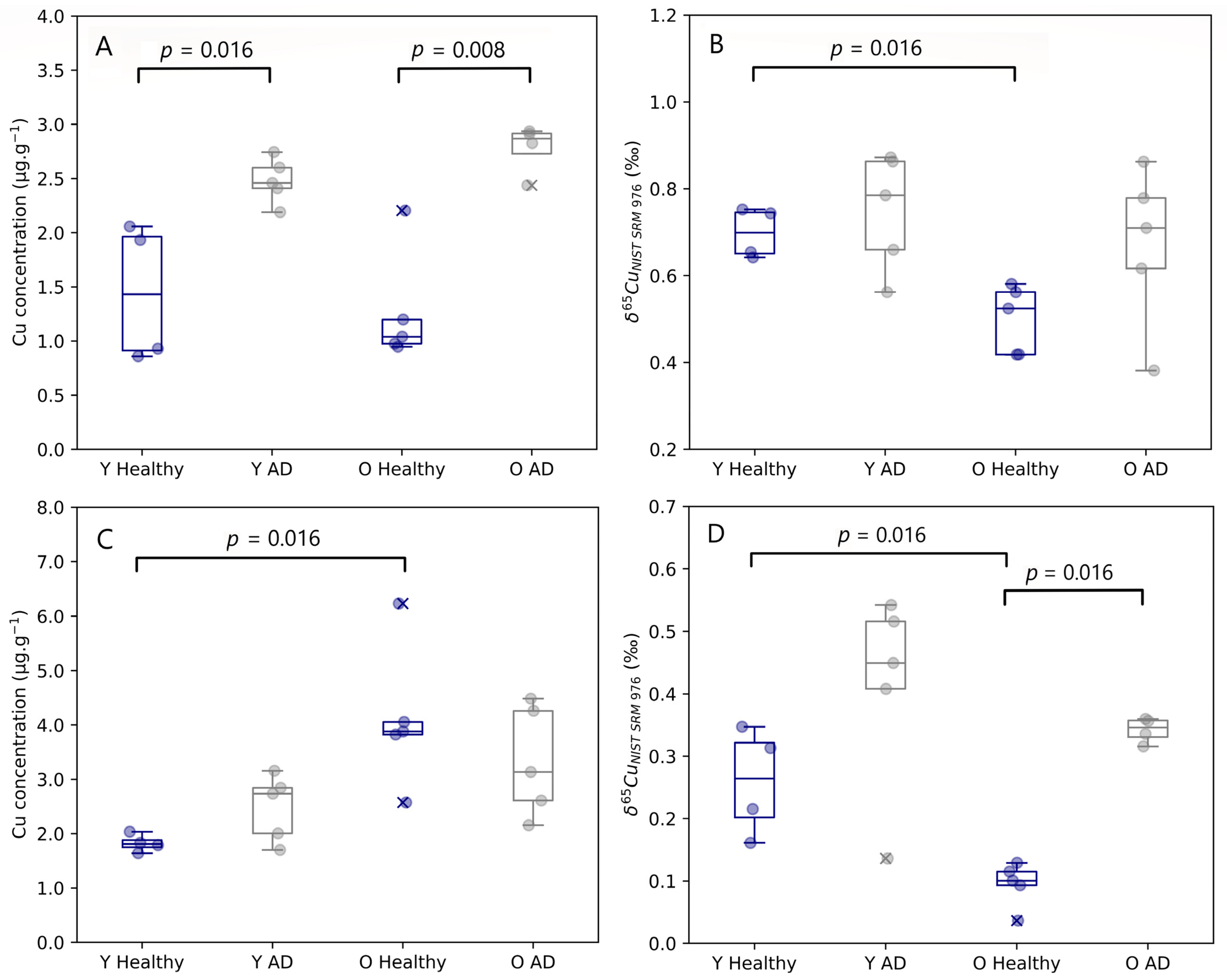
3.1. Blood Plasma
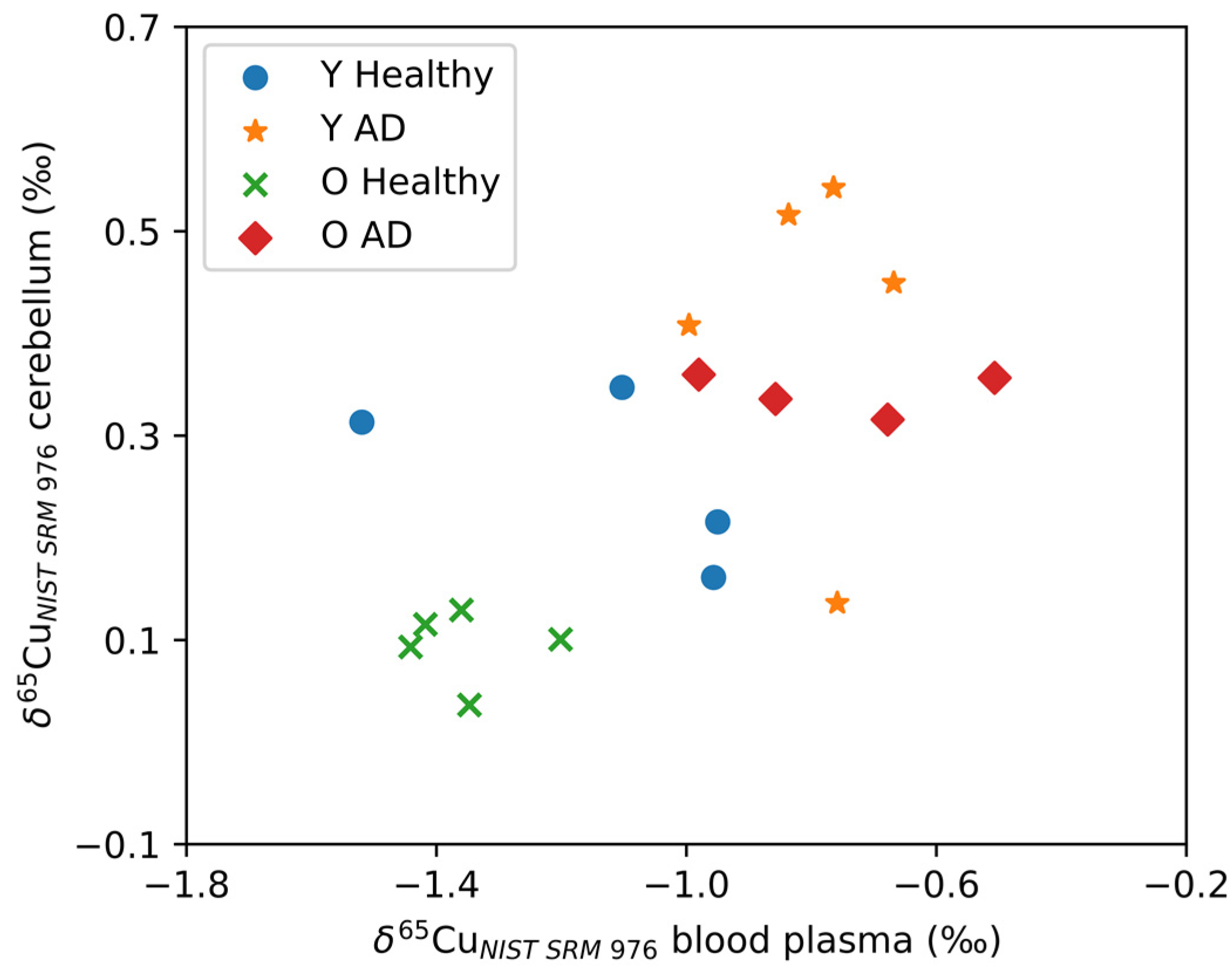
3.2. Copper Alterations in Brain Tissues
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Scheltens, P.; De Strooper, B.; Kivipelto, M.; Holstege, H.; Chételat, G.; Teunissen, C.E.; Cummings, J.; van der Flier, W.M. Alzheimer’s disease. Lancet 2021, 397, 1577–1590. [Google Scholar] [CrossRef] [PubMed]
- Ando, K.; Houben, S.; Homa, M.; de Fisenne, M.-A.; Potier, M.-C.; Erneux, C.; Brion, J.-P.; Leroy, K. Alzheimer’s Disease: Tau Pathology and Dysfunction of Endocytosis. Front. Mol. Neurosci. 2021, 13, 583755. [Google Scholar] [CrossRef] [PubMed]
- Masters, C.L.; Bateman, R.; Blennow, K.; Rowe, C.C.; Sperling, R.A.; Cummings, J.L. Alzheimer’s disease. Nat. Rev. Dis. Primer 2015, 1, 15056. [Google Scholar] [CrossRef] [PubMed]
- Bernstein, S.L.; Dupuis, N.F.; Lazo, N.D.; Wyttenbach, T.; Condron, M.M.; Bitan, G.; Teplow, D.B.; Shea, J.-E.; Ruotolo, B.T.; Robinson, C.V.; et al. Amyloid-β protein oligomerization and the importance of tetramers and dodecamers in the aetiology of Alzheimer’s disease. Nat. Chem. 2009, 1, 326–331. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Eom, K.; Kwon, T. Different Aggregation Pathways and Structures for Aβ40 and Aβ42 Peptides. Biomolecules 2021, 11, 198. [Google Scholar] [CrossRef]
- Patel, R.; Aschner, M. Commonalities between Copper Neurotoxicity and Alzheimer’s Disease. Toxics 2021, 9, 4. [Google Scholar] [CrossRef]
- Donnelly, P.S.; Xiao, Z.; Wedd, A.G. Copper and Alzheimer’s disease. Curr. Opin. Chem. Biol. 2007, 11, 128–133. [Google Scholar] [CrossRef]
- Desai, V.; Kaler, S.G. Role of copper in human neurological disorders. Am. J. Clin. Nutr. 2008, 88, 855S–858S. [Google Scholar] [CrossRef] [Green Version]
- Ejaz, H.W.; Wang, W.; Lang, M. Copper Toxicity Links to Pathogenesis of Alzheimer’s Disease and Therapeutics Approaches. Int. J. Mol. Sci. 2020, 21, 7660. [Google Scholar] [CrossRef]
- Guilloreau, L.; Combalbert, S.; Sournia-Saquet, A.; Mazarguil, H.; Faller, P. Redox Chemistry of Copper–Amyloid-β: The Generation of Hydroxyl Radical in the Presence of Ascorbate is Linked to Redox-Potentials and Aggregation State. ChemBioChem 2007, 8, 1317–1325. [Google Scholar] [CrossRef]
- Murphy, M.P.; Bayir, H.; Belousov, V.; Chang, C.J.; Davies, K.J.A.; Davies, M.J.; Dick, T.P.; Finkel, T.; Forman, H.J.; Janssen-Heininger, Y.; et al. Guidelines for measuring reactive oxygen species and oxidative damage in cells and in vivo. Nat. Metab. 2022, 4, 651–662. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Jiang, Y.; Shi, H.; Peng, Y.; Fan, X.; Li, C. The molecular mechanisms of copper metabolism and its roles in human diseases. Pflüg. Arch.—Eur. J. Physiol. 2020, 472, 1415–1429. [Google Scholar] [CrossRef] [PubMed]
- Kaler, S.G. ATP7A-related copper transport diseases—Emerging concepts and future trends. Nat. Rev. Neurol. 2011, 7, 15–29. [Google Scholar] [CrossRef] [Green Version]
- Cox, D.W.; Moore, S.D.P. Copper Transporting P-Type ATPases and Human Disease. J. Bioenerg. Biomembr. 2002, 34, 333–338. [Google Scholar] [CrossRef]
- Peña, M.M.O.; Lee, J.; Thiele, D.J. A Delicate Balance: Homeostatic Control of Copper Uptake and Distribution. J. Nutr. 1999, 129, 1251–1260. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vanhaecke, F.; Costas-Rodríguez, M. High-precision isotopic analysis of essential mineral elements: Capabilities as a diagnostic/prognostic tool. VIEW 2021, 2, 20200094. [Google Scholar] [CrossRef]
- García-Poyo, M.C.; Bérail, S.; Ronzani, A.L.; Rello, L.; García-González, E.; Nakadi, F.V.; Aramendía, M.; Resano, J.; Resano, M.; Pécheyran, C. Cu fractionation, isotopic analysis, and data processing via machine learning: New approaches for the diagnosis and follow up of Wilson’s disease via ICP-MS. J. Anal. At. Spectrom. 2023, 38, 229–242. [Google Scholar] [CrossRef]
- Costas-Rodríguez, M.; Anoshkina, Y.; Lauwens, S.; Van Vlierberghe, H.; Delanghe, J.; Vanhaecke, F. Isotopic analysis of Cu in blood serum by multi-collector ICP-mass spectrometry: A new approach for the diagnosis and prognosis of liver cirrhosis? Metallomics 2015, 7, 491–498. [Google Scholar] [CrossRef]
- Aramendía, M.; Rello, L.; Resano, M.; Vanhaecke, F. Isotopic analysis of Cu in serum samples for diagnosis of Wilson’s disease: A pilot study. J. Anal. At. Spectrom. 2013, 28, 675. [Google Scholar] [CrossRef]
- Lauwens, S.; Costas-Rodríguez, M.; Van Vlierberghe, H.; Vanhaecke, F. Cu isotopic signature in blood serum of liver transplant patients: A follow-up study. Sci. Rep. 2016, 6, 30683. [Google Scholar] [CrossRef] [Green Version]
- Télouk, P.; Puisieux, A.; Fujii, T.; Balter, V.; Bondanese, V.P.; Morel, A.-P.; Clapisson, G.; Lamboux, A.; Albarede, F. Copper isotope effect in serum of cancer patients. A pilot study. Metallomics 2015, 7, 299–308. [Google Scholar] [CrossRef] [PubMed]
- Hastuti, A.A.M.B.; Costas-Rodríguez, M.; Anoshkina, Y.; Parnall, T.; Madura, J.A.; Vanhaecke, F. High-precision isotopic analysis of serum and whole blood Cu, Fe and Zn to assess possible homeostasis alterations due to bariatric surgery. Anal. Bioanal. Chem. 2020, 412, 727–738. [Google Scholar] [CrossRef] [PubMed]
- Aranaz, M.; Costas-Rodríguez, M.; Lobo, L.; González-Iglesias, H.; Vanhaecke, F.; Pereiro, R. Pilot study of homeostatic alterations of mineral elements in serum of patients with age-related macular degeneration via elemental and isotopic analysis using ICP-mass spectrometry. J. Pharm. Biomed. Anal. 2020, 177, 112857. [Google Scholar] [CrossRef] [PubMed]
- Larner, F.; Sampson, B.; Rehkämper, M.; Weiss, D.J.; Dainty, J.R.; O’Riordan, S.; Panetta, T.; Bain, P.G. High precision isotope measurements reveal poor control of copper metabolism in Parkinsonism. Metallomics 2013, 5, 125. [Google Scholar] [CrossRef]
- Eskici, G.; Axelsen, P.H. Copper and Oxidative Stress in the Pathogenesis of Alzheimer’s Disease. Biochemistry 2012, 51, 6289–6311. [Google Scholar] [CrossRef]
- Vural, H.; Demirin, H.; Kara, Y.; Eren, I.; Delibas, N. Alterations of plasma magnesium, copper, zinc, iron and selenium concentrations and some related erythrocyte antioxidant enzyme activities in patients with Alzheimer’s disease. J. Trace Elem. Med. Biol. 2010, 24, 169–173. [Google Scholar] [CrossRef]
- Koç, E.R.; İLhan, A.; Aytürk, Z.; Acar, B.; Gürler, M.; Altuntaş, A.; KarapİRlİ, M.; Bodur, A.S. A comparison of hair and serum trace elements in patients with Alzheimer disease and healthy participants. Turk. J. Med. Sci. 2015, 45, 1034–1039. [Google Scholar] [CrossRef]
- Brewer, G.J.; Kanzer, S.H.; Zimmerman, E.A.; Celmins, D.F.; Heckman, S.M.; Dick, R. Copper and Ceruloplasmin Abnormalities in Alzheimer’s Disease. Am. J. Alzheimers Dis. Dementiasr 2010, 25, 490–497. [Google Scholar] [CrossRef]
- Pajonk, F.-G.; Kessler, H.; Supprian, T.; Hamzei, P.; Bach, D.; Schweickhardt, J.; Herrmann, W.; Obeid, R.; Simons, A.; Falkai, P.; et al. Cognitive decline correlates with low plasma concentrations of copper in patients with mild to moderate Alzheimer’s disease. J. Alzheimers Dis. 2005, 8, 23–27. [Google Scholar] [CrossRef]
- Squitti, R.; Ventriglia, M.; Barbati, G.; Cassetta, E.; Ferreri, F.; Dal Forno, G.; Ramires, S.; Zappasodi, F.; Rossini, P.M. ‘Free’ copper in serum of Alzheimer’s disease patients correlates with markers of liver function. J. Neural Transm. 2007, 114, 1589–1594. [Google Scholar] [CrossRef]
- Siotto, M.; Simonelli, I.; Pasqualetti, P.; Mariani, S.; Caprara, D.; Bucossi, S.; Ventriglia, M.; Molinario, R.; Antenucci, M.; Rongioletti, M.; et al. Association Between Serum Ceruloplasmin Specific Activity and Risk of Alzheimer’s Disease. J. Alzheimers Dis. 2016, 50, 1181–1189. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.-X.; Tan, L.; Wang, H.-F.; Ma, J.; Liu, J.; Tan, M.-S.; Sun, J.-H.; Zhu, X.-C.; Jiang, T.; Yu, J.-T. Serum Iron, Zinc, and Copper Levels in Patients with Alzheimer’s Disease: A Replication Study and Meta-Analyses. J. Alzheimers Dis. 2015, 47, 565–581. [Google Scholar] [CrossRef] [PubMed]
- Li, D.-D.; Zhang, W.; Wang, Z.-Y.; Zhao, P. Serum Copper, Zinc, and Iron Levels in Patients with Alzheimer’s Disease: A Meta-Analysis of Case-Control Studies. Front. Aging Neurosci. 2017, 9, 300. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bucossi, S.; Ventriglia, M.; Panetta, V.; Salustri, C.; Pasqualetti, P.; Mariani, S.; Siotto, M.; Rossini, P.M.; Squitti, R. Copper in Alzheimer’s Disease: A Meta-Analysis of Serum, Plasma, and Cerebrospinal Fluid Studies. J. Alzheimer’s Dis. 2011, 24, 175–185, Erratum in J. Alzheimers Dis. 2012, 30, 981–984. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Solovyev, N.; El-Khatib, A.H.; Costas-Rodríguez, M.; Schwab, K.; Griffin, E.; Raab, A.; Platt, B.; Theuring, F.; Vogl, J.; Vanhaecke, F. Cu, Fe, and Zn isotope ratios in murine Alzheimer’s disease models suggest specific signatures of amyloidogenesis and tauopathy. J. Biol. Chem. 2021, 296, 100292. [Google Scholar] [CrossRef]
- Moynier, F.; Creech, J.; Dallas, J.; Le Borgne, M. Serum and brain natural copper stable isotopes in a mouse model of Alzheimer’s disease. Sci. Rep. 2019, 9, 11894. [Google Scholar] [CrossRef] [Green Version]
- Moynier, F.; Borgne, M.L.; Lahoud, E.; Mahan, B.; Mouton-Ligier, F.; Hugon, J.; Paquet, C. Copper and zinc isotopic excursions in the human brain affected by Alzheimer’s disease. Alzheimers Dement. Diagn. Assess. Dis. Monit. 2020, 12, e12112. [Google Scholar] [CrossRef]
- Sauzéat, L.; Bernard, E.; Perret-Liaudet, A.; Quadrio, I.; Vighetto, A.; Krolak-Salmon, P.; Broussolle, E.; Leblanc, P.; Balter, V. Isotopic Evidence for Disrupted Copper Metabolism in Amyotrophic Lateral Sclerosis. iScience 2018, 6, 264–271. [Google Scholar] [CrossRef] [Green Version]
- Saito, T.; Matsuba, Y.; Mihira, N.; Takano, J.; Nilsson, P.; Itohara, S.; Iwata, N.; Saido, T.C. Single App knock-in mouse models of Alzheimer’s disease. Nat. Neurosci. 2014, 17, 661–663. [Google Scholar] [CrossRef]
- Maréchal, C.N.; Télouk, P.; Albarède, F. Precise analysis of copper and zinc isotopic compositions by plasma-source mass spectrometry. Chem. Geol. 1999, 156, 251–273. [Google Scholar] [CrossRef]
- Lauwens, S.; Costas-Rodríguez, M.; Vanhaecke, F. Ultra-trace Cu isotope ratio measurements via multi-collector ICP-mass spectrometry using Ga as internal standard: An approach applicable to micro-samples. Anal. Chim. Acta 2018, 1025, 69–79. [Google Scholar] [CrossRef]
- Baxter, D.C.; Rodushkin, I.; Engström, E.; Malinovsky, D. Revised exponential model for mass bias correction using an internal standard for isotope abundance ratio measurements by multi-collector inductively coupled plasma mass spectrometry. J. Anal. At. Spectrom. 2006, 21, 427. [Google Scholar] [CrossRef]
- Woodhead, J. A simple method for obtaining highly accurate Pb isotope data by MC-ICP-MS. J. Anal. At. Spectrom. 2002, 17, 1381–1385. [Google Scholar] [CrossRef]
- Sauzéat, L.; Costas-Rodríguez, M.; Albalat, E.; Mattielli, N.; Vanhaecke, F.; Balter, V. Inter-comparison of stable iron, copper and zinc isotopic compositions in six reference materials of biological origin. Talanta 2021, 221, 121576. [Google Scholar] [CrossRef]
- Hobin, K.; Costas-Rodríguez, M.; Van Wonterghem, E.; Vandenbroucke, R.E.; Vanhaecke, F. High-Precision Isotopic Analysis of Cu and Fe via Multi-Collector Inductively Coupled Plasma-Mass Spectrometry Reveals Lipopolysaccharide-Induced Inflammatory Effects in Blood Plasma and Brain Tissues. Front. Chem. 2022, 10, 896279. [Google Scholar] [CrossRef]
- Malavolta, M.; Piacenza, F.; Basso, A.; Giacconi, R.; Costarelli, L.; Mocchegiani, E. Serum copper to zinc ratio: Relationship with aging and health status. Mech. Ageing Dev. 2015, 151, 93–100. [Google Scholar] [CrossRef] [PubMed]
- Sullivan, J.F.; Blotcky, A.J.; Jetton, M.M.; Hahn, H.K.J.; Burch, R.E. Serum Levels of Selenium, Calcium, Copper Magnesium, Manganese and Zinc in Various Human Diseases. J. Nutr. 1979, 109, 1432–1437. [Google Scholar] [CrossRef] [PubMed]
- Remick, D.G.; Newcomb, D.E.; Bolgos, G.L.; Call, D.R. Comparison of the mortality and inflammatory response of two models of sepsis: Lipopolysaccharide vs. Cecal ligation and puncture. Shock 2000, 13, 110. [Google Scholar] [CrossRef] [PubMed]
- Vassiliev, V.; Harris, Z.L.; Zatta, P. Ceruloplasmin in neurodegenerative diseases. Brain Res. Rev. 2005, 49, 633–640. [Google Scholar] [CrossRef]
- Costas-Rodríguez, M.; Van Campenhout, S.; Hastuti, A.A.M.B.; Devisscher, L.; Van Vlierberghe, H.; Vanhaecke, F. Body distribution of stable copper isotopes during the progression of cholestatic liver disease induced by common bile duct ligation in mice. Metallomics 2019, 11, 1093–1103. [Google Scholar] [CrossRef]
- Squitti, R.; Ghidoni, R.; Simonelli, I.; Ivanova, I.D.; Colabufo, N.A.; Zuin, M.; Benussi, L.; Binetti, G.; Cassetta, E.; Rongioletti, M.; et al. Copper dyshomeostasis in Wilson disease and Alzheimer’s disease as shown by serum and urine copper indicators. J. Trace Elem. Med. Biol. 2018, 45, 181–188. [Google Scholar] [CrossRef] [PubMed]
- Resano, M.; Aramendía, M.; Rello, L.; Calvo, M.L.; Bérail, S.; Pécheyran, C. Direct determination of Cu isotope ratios in dried urine spots by means of fs-LA-MC-ICPMS. Potential to diagnose Wilson’s disease. J Anal Spectrom 2013, 28, 98–106. [Google Scholar] [CrossRef]
- Miller, K.A.; Keenan, C.M.; Martin, G.R.; Jirik, F.R.; Sharkey, K.A.; Wieser, M.E. The expression levels of cellular prion protein affect copper isotopic shifts in the organs of mice. J. Anal. At. Spectrom. 2016, 31, 2015–2022. [Google Scholar] [CrossRef]
- Mahan, B.; Tacail, T.; Lewis, J.; Elliott, T.; Habekost, M.; Turner, S.; Chung, R.; Moynier, F. Exploring the K isotope composition of Göttingen minipig brain regions, and implications for Alzheimer’s disease. Metallomics 2022, 14, mfac090. [Google Scholar] [CrossRef]
- Ji, X.; Wang, H.; Zhu, M.; He, Y.; Zhang, H.; Chen, X.; Gao, W.; Fu, Y.; for the Alzheimer’s Disease Neuroimaging Initiative. Brainstem atrophy in the early stage of Alzheimer’s disease: A voxel-based morphometry study. Brain Imaging Behav. 2021, 15, 49–59. [Google Scholar] [CrossRef]
- Erdő, F.; Denes, L.; de Lange, E. Age-associated physiological and pathological changes at the blood–brain barrier: A review. J. Cereb. Blood Flow Metab. 2017, 37, 4–24. [Google Scholar] [CrossRef] [Green Version]
- Gaier, E.D.; Eipper, B.A.; Mains, R.E. Copper signaling in the mammalian nervous system: Synaptic effects. J. Neurosci. Res. 2012, 91, 2–19. [Google Scholar] [CrossRef] [Green Version]
- Loeffler, D.A.; LeWitt, P.A.; Juneau, P.L.; Sima, A.A.F.; Nguyen, H.-U.; DeMaggio, A.J.; Brickman, C.M.; Brewer, G.J.; Dick, R.D.; Troyer, M.D.; et al. Increased regional brain concentrations of ceruloplasmin in neurodegenerative disorders. Brain Res. 1996, 738, 265–274. [Google Scholar] [CrossRef]
- Schrag, M.; Mueller, C.; Oyoyo, U.; Smith, M.A.; Kirsch, W.M. Iron, zinc and copper in the Alzheimer’s disease brain: A quantitative meta-analysis. Some insight on the influence of citation bias on scientific opinion. Prog. Neurobiol. 2011, 94, 296–306. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Phinney, A.L.; Horne, P.; Yang, J.; Janus, C.; Bergeron, C.; Westaway, D. Mouse models of Alzheimer’s disease: The long and filamentous road. Neurol. Res. 2003, 25, 590–600. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Instrument Settings | Agilent 8800 ICP-MS/MS | Instrument Settings | Neptune Plus MC-ICP-MS |
|---|---|---|---|
| Sample uptake rate (µL.min−1) | 350 | Sample uptake rate (µL.min−1) | 100 |
| Plasma gas flow rate (L.min−1)–Ar | 15 | Plasma gas flow rate (L.min−1)–Ar | 15 |
| Auxiliary gas flow rate (L.min−1)–Ar | 0.9 | Auxiliary gas flow rate (L.min−1)–Ar | 0.7-0.8 |
| Nebulizer gas flow rate (L.min−1)–Ar | 1.12 | Nebulizer gas flow rate (L.min−1)–Ar | 1.0-1.1 |
| Collision gas flow rate (mL.min−1)–He | 1.0 | ||
| Reaction gas flow rate (mL.min−1)–NH3 | 3.0 | ||
| Rf Power (W) | 1550 | Rf Power (W) | 1200 |
| Sampling cone | Ni tip with Cu base | Sampling cone | Ni, Jet cone |
| Skimmer cone | Ni | Skimmer cone | Ni, X-type |
| Data acquisition parameters | Agilent 8800 ICP-MS/MS | Data acquisition Parameters | Neptune Plus MC-ICP-MS |
| Operation mode | NH3/He (10%/90%) mode | Scan type | Static, multicollection |
| Integration time (s) | 1 | Resolution mode | Pseudo-medium |
| Replicates | 10 | Blocks × cycles | 9 × 5 |
| Sweeps | 100 | Integration time (s) | 4.194 |
| Nuclides monitored | 65Cu+ → 65Cu(14N1H3)2+, 71Ga+ (on mass) | Cup configuration—Cu & Ga (IS) | L4: 63Cu, L2: 65Cu, C: 67Zn, H2: 69Ga, H4: 71Ga |
| Groups | Sample Type | Cu Concentration (µg.g−1) | δ65Cu (‰) | ||||
|---|---|---|---|---|---|---|---|
| Median | IQR | N | Median | IQR | N | ||
| Y Healthy | Brain stem | 1.54 | 1.40 | 4 | 0.70 | 0.11 | 4 |
| Cerebellum | 1.81 | 0.31 | 4 | 0.26 | 0.16 | 4 | |
| Cortex | 2.88 | 0.62 | 4 | 0.53 | 0.26 | 4 | |
| Hippocampus | 2.67 | 1.94 | 4 | 0.55 | 0.21 | 4 | |
| Y AD | Brain stem | 2.46 | 0.37 | 5 | 0.78 | 0.25 | 5 |
| Cerebellum | 2.74 | 1.15 | 5 | 0.45 | 0.26 | 5 | |
| Cortex | 3.22 | 0.63 | 5 | 0.63 | 0.29 | 5 | |
| Hippocampus | 3.51 | 1.62 | 5 | 0.63 | 0.18 | 5 | |
| O Healthy | Brain stem | 1.04 | 1.00 | 5 | 0.52 | 0.15 | 5 |
| Cerebellum | 3.88 | 1.94 | 5 | 0.10 | 0.06 | 5 | |
| Cortex | 3.06 | 0.69 | 5 | 0.44 | 0.14 | 5 | |
| Hippocampus | 1.54 | 3.54 | 5 | 0.49 | 0.09 | 5 | |
| O AD | Brain stem | 2.91 | 0.45 | 5 | 0.68 | 0.32 | 5 |
| Cerebellum | 3.70 | 2.03 | 5 | 0.35 | 0.04 | 4 | |
| Cortex | 3.22 | 0.35 | 5 | 0.52 | 0.25 | 5 | |
| Hippocampus | 3.80 | 1.31 | 4 | 0.53 | 0.22 | 5 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hobin, K.; Costas-Rodríguez, M.; Van Wonterghem, E.; Vandenbroucke, R.E.; Vanhaecke, F. Alzheimer’s Disease and Age-Related Changes in the Cu Isotopic Composition of Blood Plasma and Brain Tissues of the APPNL-G-F Murine Model Revealed by Multi-Collector ICP-Mass Spectrometry. Biology 2023, 12, 857. https://doi.org/10.3390/biology12060857
Hobin K, Costas-Rodríguez M, Van Wonterghem E, Vandenbroucke RE, Vanhaecke F. Alzheimer’s Disease and Age-Related Changes in the Cu Isotopic Composition of Blood Plasma and Brain Tissues of the APPNL-G-F Murine Model Revealed by Multi-Collector ICP-Mass Spectrometry. Biology. 2023; 12(6):857. https://doi.org/10.3390/biology12060857
Chicago/Turabian StyleHobin, Kasper, Marta Costas-Rodríguez, Elien Van Wonterghem, Roosmarijn E. Vandenbroucke, and Frank Vanhaecke. 2023. "Alzheimer’s Disease and Age-Related Changes in the Cu Isotopic Composition of Blood Plasma and Brain Tissues of the APPNL-G-F Murine Model Revealed by Multi-Collector ICP-Mass Spectrometry" Biology 12, no. 6: 857. https://doi.org/10.3390/biology12060857
APA StyleHobin, K., Costas-Rodríguez, M., Van Wonterghem, E., Vandenbroucke, R. E., & Vanhaecke, F. (2023). Alzheimer’s Disease and Age-Related Changes in the Cu Isotopic Composition of Blood Plasma and Brain Tissues of the APPNL-G-F Murine Model Revealed by Multi-Collector ICP-Mass Spectrometry. Biology, 12(6), 857. https://doi.org/10.3390/biology12060857