Circadian Clocks in Fish—What Have We Learned so far?

{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
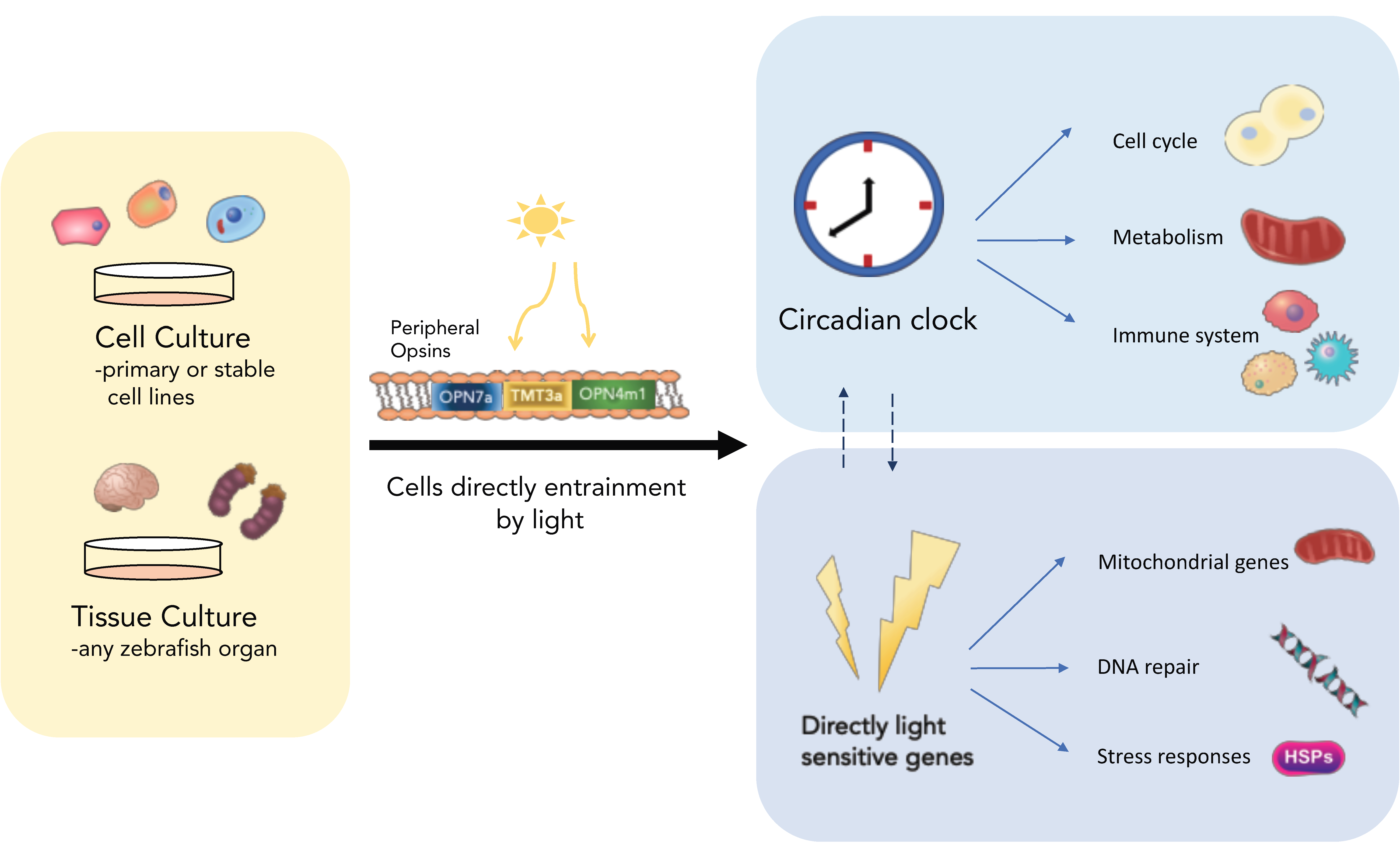
2. Peripheral Clocks and the Emergence of Cell Lines
2.1. Clocks Everywhere
2.2. Global Light Sensitivity
2.3. Development of the Clock and Its Relevance
3. The Importance of Light
3.1. The Photopigments
3.2. Light Input Pathway
3.3. Light Sensitivity in Fish—Light Detection Is Everywhere
3.4. Alternative Fish Models—Cavefish
4. The Outputs
4.1. Sleep and Rhythmic Behaviour
4.2. The Cell Cycle
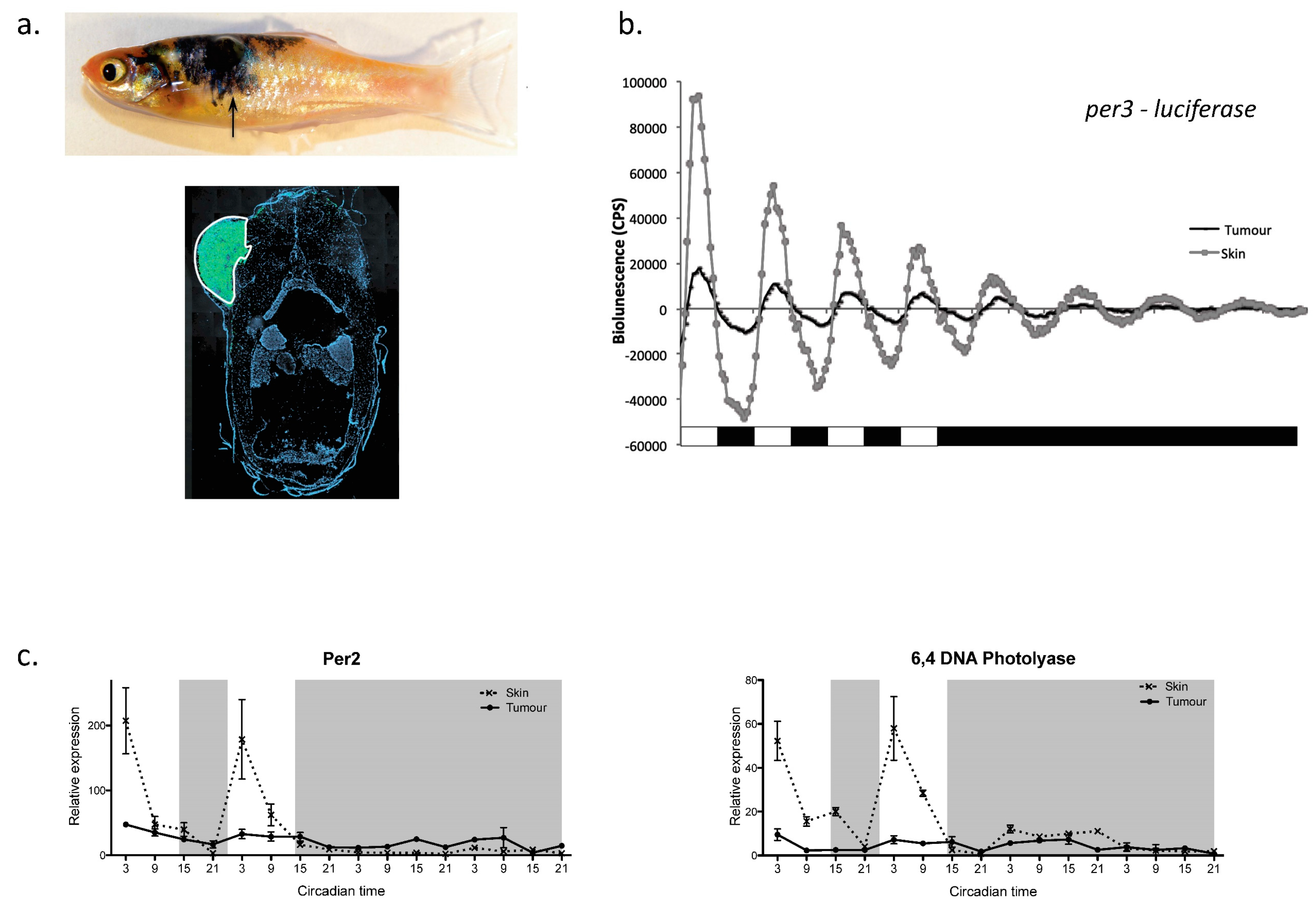
4.3. The Clock and Cancer
5. Conclusions
Funding
Acknowledgments
Conflicts of Interest
References
- Steindal, I.; Beale, A.; Yamamoto, Y.; Whitmore, D. Development of the Astyanax mexicanus circadian clock and non-visual light responses. Dev. Biol. 2018, 441, 345–354. [Google Scholar] [CrossRef]
- Beale, A.; Guibal, C.; Tamai, T.K.; Klotz, L.; Cowen, S.; Peyric, E.; Reynoso, V.H.; Yamamoto, Y.; Whitmore, D. Circadian rhythms in Mexican blind cavefish Astyanax mexicanus in the lab and in the field. Nat. Commun. 2013, 4, 2769. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beale, A.; Whitmore, D.; Moran, D.B. Life in a dark biosphere: A review of circadian physiology in “arrhythmic” environments. J. Comp. Physiol. B 2016, 186, 947–968. [Google Scholar] [CrossRef] [PubMed]
- Cavallari, N.; Frigato, E.; Vallone, D.; Fröhlich, N.; Lopez-Olmeda, J.F.; Foà, A.; Berti, R.; Sánchez-Vázquez, F.J.; Bertolucci, C.; Foulkes, N.S. A blind circadian clock in cavefish reveals that opsins mediate peripheral clock photoreception. PLoS Biol. 2011, 9, e1001142. [Google Scholar] [CrossRef]
- Foulkes, N.; Whitmore, D.; Vallone, D.; Bertolucci, C. Studying the evolution of the vertebrate circadian clock: The power of fish as comparative models. Adv. Genet. 2016, 95, 1–30. [Google Scholar]
- Tosini, G.; Menaker, M. Circadian rhythms in cultured mammalian retina. Science 1996, 272, 419–421. [Google Scholar] [CrossRef] [PubMed]
- Whitmore, D.; Foulkes, N.; Strähle, U.; Sassone-Corsi, P. Zebrafish Clock rhythmic expression reveals independent peripheral circadian oscillators. Nat. Neurosci. 1998, 1, 701–707. [Google Scholar] [CrossRef]
- Whitmore, D.; Foulkes, N.; Sassone-Corsi, P. Light acts directly on organs and cells in culture to set the vertebrate circadian clock. Nature 2000, 404, 87–91. [Google Scholar] [CrossRef]
- Balsalobre, A. Clock genes in mammalian peripheral tissues. Cell Tissue Res. 2002, 309, 193–199. [Google Scholar] [CrossRef] [PubMed]
- Chang, D.C.; Reppert, S.M. The circadian clocks of mice and men. Neuron 2001, 29, 555–558. [Google Scholar] [CrossRef]
- King, D.P.; Zhao, Y.; Sangoram, A.M.; Wilsbacher, L.D.; Tanaka, M.; Antoch, M.P.; Steeves, T.D.; Vitaterna, M.H.; Kornhauser, J.M.; Lowrey, P.L.; et al. Positional cloning of the mouse circadian clock gene. Cell 1997, 89, 641–653. [Google Scholar] [CrossRef]
- Panda, S.; Antoch, M.P.; Miller, B.H.; Su, A.I.; Schook, A.B.; Straume, M.; Schultz, P.G.; Kay, S.A.; Takahashi, J.S.; Hogenesch, J.B. Coordinated transcription of key pathways in the mouse by the circadian clock. Cell 2002, 109, 307–320. [Google Scholar] [CrossRef]
- Balsalobre, A.; Damiola, F.; Schibler, U. A serum shock induces circadian gene expression in mammalian tissue culture cells. Cell 1998, 93, 929–937. [Google Scholar] [CrossRef]
- Sidiropoulos, K.; Viteri, G.; Sevilla, C.; Jupe, S.; Webber, M.; Orlic-Milacic, M.; Jassal, B.; May, B.; Shamovsky, V.; Duenas, C.; et al. Reactome enhanced pathway visualization. Bioinformatics 2017, 33, 3461–3467. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martín-Robles, Á.J.; Whitmore, D.; Sánchez-Vázquez, F.J.; Pendón, C.; Muñoz-Cueto, J.A. Cloning, tissue expression pattern and daily rhythms of Period1, Period2, and Clock transcripts in the flatfish Senegalese sole, Solea senegalensis. J. Comp. Physiol. B Biochem. Syst. Environ. Physiol. 2012, 182, 673–685. [Google Scholar] [CrossRef]
- Martín-Robles, A.J.; Isorna, E.; Whitmore, D.; Muñoz-Cueto, J.A.; Pendón, C. The clock gene Period3 in the nocturnal flatfish Solea senegalensis: Molecular cloning, tissue expression and daily rhythms in central areas. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2011, 159, 7–15. [Google Scholar] [CrossRef] [PubMed]
- Blanco-Vives, B.; Aliaga-Guerrero, M.; Cañavate, J.P.; García-Mateos, G.; Martín-Robles, A.J.; Herrera-Pérez, P.; Muñoz-Cueto, J.A.; Sánchez-Vázquez, F.J. Metamorphosis induces a light-dependent switch in Senegalese sole (Solea senegalensis) from diurnal to nocturnal behavior. J. Biol. Rhythm. 2012, 27, 135–144. [Google Scholar] [CrossRef]
- Mohawk, J.A.; Green, C.B.; Takahashi, J.S. Central and peripheral circadian clocks in mammals. Annu. Rev. Neurosci. 2012, 35, 445–462. [Google Scholar] [CrossRef]
- Chen, S.; Reichert, S.; Singh, C.; Oikonomou, G.; Rihel, J.; Prober, D.A. Light-dependent regulation of sleep and wake states by prokineticin 2 in zebrafish. Neuron 2017, 95, 153–168.e6. [Google Scholar] [CrossRef]
- Gandhi, A.; Mosser, E.; Oikonomou, G.; Neuron, P.D. Melatonin is required for the circadian regulation of sleep. Neuron 2015, 95, 153–168.e6. [Google Scholar] [CrossRef]
- Ben-Moshe Livne, Z.; Alon, S.; Vallone, D.; Bayleyen, Y.; Tovin, A.; Shainer, I.; Nisembaum, L.G.; Aviram, I.; Smadja-Storz, S.; Fuentes, M.; et al. Genetically blocking the zebrafish pineal clock affects circadian behavior. PLoS Genet. 2016, 12, e1006445. [Google Scholar] [CrossRef] [PubMed]
- Dekens, M.P.; Santoriello, C.; Vallone, D.; Grassi, G.; Whitmore, D.; Foulkes, N.S. Light regulates the cell cycle in zebrafish. Curr. Biol. 2003, 13, 2051–2057. [Google Scholar] [CrossRef] [PubMed]
- Ziv, L.; Gothilf, Y. Circadian time-keeping during early stages of development. Proc. Natl. Acad. Sci. USA 2006, 103, 4146–4151. [Google Scholar] [CrossRef] [Green Version]
- Ziv, L.; Levkovitz, S.; Toyama, R.; Falcon, J.; Gothilf, Y. Functional development of the zebrafish pineal gland: Light-induced expression of period2 is required for onset of the circadian clock. J. Neuroendocrinol. 2005, 17, 314–320. [Google Scholar] [CrossRef] [PubMed]
- Kaneko, M.; Cahill, G.M. Light-dependent development of circadian gene expression in transgenic zebrafish. PLoS Biol. 2005, 3, e34. [Google Scholar] [CrossRef]
- Laranjeiro, R.; Whitmore, D. Transcription factors involved in retinogenesis are co-optedby the circadian clockfollowing photoreceptor differentiation. Development 2014, 141, 2644–2656. [Google Scholar] [CrossRef]
- Cheng, P.Y.; Lin, C.C.; Wu, C.S.; Lu, Y.F.; Lin, C.Y.; Chung, C.C.; Chu, C.Y.; Huang, C.J.; Tsai, C.Y.; Korzh, S.; et al. Zebrafish cdx1b regulates expression of downstream factors of Nodal signaling during early endoderm formation. Development 2008, 135, 941–952. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tamai, T.K.; Young, L.C.; Whitmore, D. Light signaling to the zebrafish circadian clock by Cryptochrome 1a. Proc. Natl. Acad. Sci. USA 2007, 104, 14712–14717. [Google Scholar] [CrossRef] [Green Version]
- Goldman, B. Mammalian photoperiodic system: Formal properties and neuroendocrine mechanisms of photoperiodic time measurement. J. Biol. Rhythm. 2001, 16, 283–301. [Google Scholar] [CrossRef]
- Daan, S. Learning and circadian behavior. J. Biol. Rhythm. 2000, 15, 296–299. [Google Scholar] [CrossRef]
- Nishiwaki-Ohkawa, T.; Yoshimura, T. Molecular basis for regulating seasonal reproduction in vertebrates. J. Endocrinol. 2016, 229, R117–R127. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tamai, T.K.; Vardhanabhuti, V.; Foulkes, N.S.; Whitmore, D. Early embryonic light detection improves survival. Curr. Biol. 2004, 14, 446. [Google Scholar] [CrossRef]
- Davies, W.I.; Tamai, T.K.; Zheng, L.; Fu, J.K.; Rihel, J.; Foster, R.G.; Whitmore, D.; Hankins, M.W. An extended family of novel vertebrate photopigments is widely expressed and displays a diversity of function. Genome Res. 2015, 25, 1666–1679. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hirayama, J.; Miyamura, N.; Uchida, Y.; Asaoka, Y.; Honda, R.; Sawanobori, K.; Todo, T.; Yamamoto, T.; Sassone-Corsi, P.; Nishina, H. Common light signaling pathways controlling DNA repair and circadian clock entrainment in zebrafish. Cell Cycle 2009, 8, 2794–2801. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hirayama, J.; Cho, S.; Sassone-Corsi, P. Circadian control by the reduction/oxidation pathway: Catalase represses light-dependent clock gene expression in the zebrafish. Proc. Natl. Acad. Sci. USA 2007, 104, 15747–15752. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cermakian, N.; Pando, M.P.; Thompson, C.L.; Pinchak, A.B.; Selby, C.P.; Gutierrez, L.; Wells, D.E.; Cahill, G.M.; Sancar, A.; Sassone-Corsi, P. Light induction of a vertebrate clock gene involves signaling through blue-light receptors and MAP kinases. Curr. Biol. 2002, 12, 844–848. [Google Scholar] [CrossRef]
- Mracek, P.; Pagano, C.; Frohlich, N.; Idda, L.M.; Cuesta, I.H.; Lopez-Olmeda, J.F.; Sánchez-Vázquez, F.J.; Vallone, D.; Foulkes, N.S. ERK signalling regulates light-induced gene expression via D-Box enhancers in a differential wavelength-dependent manner. PLoS ONE 2013, 8, e67858. [Google Scholar] [CrossRef]
- Pagano, C.; Siauciunaite, R.; Idda, M.L.; Ruggiero, G.; Ceinos, R.M.; Pagano, M.; Frigato, E.; Bertolucci, C.; Foulkes, N.S.; Vallone, D. Evolution shapes the responsiveness of the D-box enhancer element to light and reactive oxygen species in vertebrates. Sci. Rep. 2018, 8, 13180. [Google Scholar] [CrossRef]
- Ramos, B.; Moraes, M.N.; Poletini, M.O.; Lima, L.H.; Castrucci, A.M. From blue light to clock genes in zebrafish ZEM-2S cells. PLoS ONE 2014, 9, e106252. [Google Scholar] [CrossRef]
- Ondrusova, K.; Fatehi, M.; Barr, A.; Czarnecka, Z.; Long, W.; Suzuki, K.; Campbell, S.; Philippaert, K.; Hubert, M.; Tredget, E.; et al. Subcutaneous white adipocytes express a light sensitive signaling pathway mediated via a melanopsin/TRPC channel axis. Sci. Rep. 2017, 7, 16332. [Google Scholar] [CrossRef] [Green Version]
- Rollag, M.D.; Berson, D.M.; Provencio, I. Melanopsin, ganglion-cell photoreceptors, and mammalian photoentrainment. J. Biol. Rhythm. 2003, 18, 227–234. [Google Scholar] [CrossRef]
- Hatori, M.; Panda, S. The emerging roles of melanopsin in behavioral adaptation to light. Trends Mol. Med. 2010, 16, 435–446. [Google Scholar] [CrossRef] [Green Version]
- Buhr, E.D.; Yue, W.W.; Ren, X.; Jiang, Z.; Liao, H.W.; Mei, X.; Vemaraju, S.; Nguyen, M.T.; Reed, R.R.; Lang, R.A.; et al. Neuropsin (OPN5)-mediated photoentrainment of local circadian oscillators in mammalian retina and cornea. Proc. Natl. Acad. Sci. USA 2015, 112, 13093–13098. [Google Scholar] [CrossRef] [Green Version]
- Kojima, D.; Mori, S.; Torii, M.; Wada, A.; Morishita, R.; Fukada, Y. UV-sensitive photoreceptor protein OPN5 in humans and mice. PLoS ONE 2011, 6, e26388. [Google Scholar] [CrossRef]
- Vatine, G.; Vallone, D.; Appelbaum, L.; Mracek, P.; Ben-Moshe, Z.; Lahiri, K.; Gothilf, Y.; Foulkes, N.S. Light directs zebrafish period2 expression via conserved D and E boxes. PLoS Biol. 2009, 7, e1000223. [Google Scholar] [CrossRef]
- Mracek, P.; Santoriello, C.; Idda, M.; Pagano, C.; Ben-Moshe, Z.; Gothilf, Y.; Vallone, D.; Foulkes, N.S. Regulation of per and cry genes reveals a central role for the D-box enhancer in light-dependent gene expression. PLoS ONE 2012, 7, e51278. [Google Scholar] [CrossRef]
- Hirayama, J.; Cardone, L.; Doi, M.; Sassone-Corsi, P. Common pathways in circadian and cell cycle clocks: Light-dependent activation of Fos/AP-1 in zebrafish controls CRY-1a and WEE-1. Proc. Natl. Acad. Sci. USA 2005, 102, 10194–10199. [Google Scholar] [CrossRef] [Green Version]
- Ishikawa, T.; Hirayama, J.; Kobayashi, Y.; Todo, T. Zebrafish CRY represses transcription mediated by CLOCK:BMAL heterodimer without inhibiting its binding to DNA. Genes Cells 2002, 7, 1073–1086. [Google Scholar] [CrossRef]
- Cuesta, I.H.; Lahiri, K.; Lopez-Olmeda, J.F.; Loosli, F.; Foulkes, N.S.; Vallone, D. Differential maturation of rhythmic clock gene expression during early development in medaka (Oryzias latipes). Chronobiol. Int. 2014, 31, 468–478. [Google Scholar] [CrossRef]
- Ben-Moshe, Z.; Alon, S.; Mracek, P.; Faigenbloom, L.; Tovin, A.; Vatine, G.D.; Eisenberg, E.; Foulkes, N.S.; Gothilf, Y. The light-induced transcriptome of the zebrafish pineal gland reveals complex regulation of the circadian clockwork by light. Nucleic Acids Res. 2014, 42, 3750–3767. [Google Scholar] [CrossRef]
- Gavriouchkina, D.; Fischer, S.; Ivacevic, T.; Stolte, J.; Benes, V.; Dekens, M.P. Thyrotroph embryonic factor regulates light-induced transcription of repair genes in zebrafish embryonic cells. PLoS ONE 2010, 5, e12542. [Google Scholar] [CrossRef] [PubMed]
- Weger, B.D.; Sahinbas, M.; Otto, G.W.; Mracek, P.; Armant, O.; Dolle, D.; Lahiri, K.; Vallone, D.; Ettwiller, L.; Geisler, R.; et al. The light responsive transcriptome of the zebrafish: Function and regulation. PLoS ONE 2011, 6, e17080. [Google Scholar] [CrossRef]
- Minchenko, O.; Opentanova, I.; Minchenko, D.; Ogura, T.; Esumi, H. Hypoxia induces transcription of 6-phosphofructo-2-kinase/fructose-2, 6-biphosphatase-4 gene via hypoxia-inducible factor-1α activation. FEBS Lett. 2004, 576, 14–20. [Google Scholar] [CrossRef] [PubMed]
- Fernandes, A.M.; Fero, K.; Arrenberg, A.B.; Bergeron, S.A.; Driever, W.; Burgess, H.A. Deep brain photoreceptors control light-seeking behavior in zebrafish larvae. Curr. Biol. 2012, 22, 2042–2047. [Google Scholar] [CrossRef] [PubMed]
- Colli, L.; Paglianti, A.; Berti, R.; Gandolfi, G.; Tagliavini, J. Molecular phylogeny of the blind cavefish Phreatichthys andruzzii and Garra barreimiae within the family Cyprinidae. Environ. Biol. Fishes 2009, 84, 95–107. [Google Scholar] [CrossRef]
- Ceinos, R.M.; Frigato, E.; Pagano, C.; Fröhlich, N.; Negrini, P.; Cavallari, N.; Vallone, D.; Fuselli, S.; Bertolucci, C.; Foulkes, N.S. Mutations in blind cavefish target the light-regulated circadian clock gene, period 2. Sci. Rep. 2018, 8, 8754. [Google Scholar] [CrossRef] [PubMed]
- Gross, J.B. The complex origin of Astyanax cavefish. BMC Evol. Biol. 2012, 12, 105. [Google Scholar] [CrossRef] [PubMed]
- Bradic, M.; Beerli, P.; García-de León, F.J.; Esquivel-Bobadilla, S.; Borowsky, R.L. Gene flow and population structure in the Mexican blind cavefish complex (Astyanax mexicanus). BMC Evol. Biol. 2012, 12, 9. [Google Scholar] [CrossRef] [PubMed]
- Wlikens, H. Genetic interpretation of regressive evolutionary processes: Studies on hybrid eyes of two Astyanax cave populations (Characidae, Pisces). Evolution 1971, 25, 530–544. [Google Scholar] [CrossRef] [PubMed]
- Oikonomou, G.; Prober, D.A. Attacking sleep from a new angle: Contributions from zebrafish. Curr. Opin. Neurobiol. 2017, 44, 80–88. [Google Scholar] [CrossRef] [PubMed]
- Singh, C.; Oikonomou, G.; Prober, D.A. Norepinephrine is required to promote wakefulness and for hypocretin-induced arousal in zebrafish. Elife 2015, 4, e07000. [Google Scholar] [CrossRef] [PubMed]
- Woods, I.G.; Schoppik, D.; Shi, V.J.; Zimmerman, S.; Coleman, H.A.; Greenwood, J.; Soucy, E.R.; Schier, A.F. Neuropeptidergic signaling partitions arousal behaviors in zebrafish. J. Neurosci. 2014, 34, 3142–3160. [Google Scholar] [CrossRef] [PubMed]
- Appelbaum, L.; Wang, G.X.; Maro, G.S.; Mori, R.; Tovin, A.; Marin, W.; Yokogawa, T.; Kawakami, K.; Smith, S.J.; Gothilf, Y.; et al. Sleep–wake regulation and hypocretin–melatonin interaction in zebrafish. Proc. Natl. Acad. Sci. USA 2009, 106, 21942–21947. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moore, H.; Whitmore, D. Circadian rhythmicity and light sensitivity of the zebrafish brain. PLoS ONE 2014, 9, e86176. [Google Scholar] [CrossRef] [PubMed]
- Laranjeiro, R.; Tamai, T.K.; Peyric, E.; Krusche, P.; Ott, S.; Whitmore, D. Cyclin-dependent kinase inhibitor p20 controls circadian cell-cycle timing. Proc. Natl. Acad. Sci. USA 2013, 110, 6835–6840. [Google Scholar] [CrossRef] [PubMed]
- Tamai, T.; Young, L.C.; Cox, C.A.; Whitmore, D. Light acts on the zebrafish circadian clock to suppress rhythmic mitosis and cell proliferation. J. Biol. Rhythm. 2012, 27, 226–236. [Google Scholar] [CrossRef]
- Bjarnason, G.; Jordan, R.; Sothern, R. Circadian variation in the expression of cell-cycle proteins in human oral epithelium. Am. J. Pathol. 1999, 154, 613–622. [Google Scholar] [CrossRef]
- Bjarnason, G.; Jordan, R.C.; Wood, P.A.; Li, Q.; Lincoln, D.W.; Sothern, R.B.; Hrushesky, W.J.; Ben-David, Y. Circadian expression of clock genes in human oral mucosa and skin: Association with specific cell-cycle phases. Am. J. Pathol. 2001, 158, 1793–1801. [Google Scholar] [CrossRef]
- Johnson, C.H. Circadian clocks and cell division: what’s the pacemaker? Cell Cycle 2010, 9, 3864–3873. [Google Scholar] [CrossRef]
- Feillet, C.; Krusche, P.; Tamanini, F.; Janssens, R.C.; Downey, M.J.; Martin, P.; Teboul, M.; Saito, S.; Lévi, F.A.; Bretschneider, T.; et al. Phase locking and multiple oscillating attractors for the coupled mammalian clock and cell cycle. Proc. Natl. Acad. Sci. USA 2014, 111, 9828–9833. [Google Scholar] [CrossRef] [Green Version]
- Bieler, J.; Cannavo, R.; Gustafson, K.; Gobet, C.; Gatfield, D.; Naef, F. Robust synchronization of coupled circadian and cell cycle oscillators in single mammalian cells. Mol. Syst. Biol. 2014, 10, 739. [Google Scholar] [CrossRef] [PubMed]
- Carr, A.-J.F.; Whitmore, D. Imaging of single light-responsive clock cells reveals fluctuating free-running periods. Nat. Cell Biol. 2005, 7, 319–321. [Google Scholar] [CrossRef] [PubMed]
- Laranjeiro, R.; Tamai, T.; Letton, W.; Hamilton, N.; Whitmore, D. Circadian Clock Synchronization of the Cell Cycle in Zebrafish Occurs through a Gating Mechanism Rather Than a Period-phase Locking Process. J. Biol. Rhythm. 2018, 33, 137–150. [Google Scholar] [CrossRef] [PubMed]
- Filipski, E.; Lévi, F. Circadian disruption in experimental cancer processes. Integr. Cancer Ther. 2009, 4, 298–302. [Google Scholar] [CrossRef] [PubMed]
- Filipski, E.; Li, X.M.; Lévi, F. Disruption of circadian coordination and malignant growth. Cancer Causes Control 2006, 17, 509–514. [Google Scholar] [CrossRef] [PubMed]
- Lahti, T.; Merikanto, I.; Partonen, T. Circadian clock disruptions and the risk of cancer. Ann. Med. 2012, 44, 847–853. [Google Scholar] [CrossRef] [PubMed]
- Hamilton, N.; Diaz-de-Cerio, N.; Whitmore, D. Impaired light detection of the circadian clock in a zebrafish melanoma model. Cell Cycle 2015, 14, 1232–1241. [Google Scholar] [CrossRef] [PubMed] [Green Version]



© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Frøland Steindal, I.A.; Whitmore, D. Circadian Clocks in Fish—What Have We Learned so far? Biology 2019, 8, 17. https://doi.org/10.3390/biology8010017
Frøland Steindal IA, Whitmore D. Circadian Clocks in Fish—What Have We Learned so far? Biology. 2019; 8(1):17. https://doi.org/10.3390/biology8010017
Chicago/Turabian StyleFrøland Steindal, Inga A., and David Whitmore. 2019. "Circadian Clocks in Fish—What Have We Learned so far?" Biology 8, no. 1: 17. https://doi.org/10.3390/biology8010017
APA StyleFrøland Steindal, I. A., & Whitmore, D. (2019). Circadian Clocks in Fish—What Have We Learned so far? Biology, 8(1), 17. https://doi.org/10.3390/biology8010017