Collimated Microbeam Reveals that the Proportion of Non-Damaged Cells in Irradiated Blastoderm Determines the Success of Development in Medaka (Oryzias latipes) Embryos


 ,
,  ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Ethics
2.2. Fish and Embryos
2.3. Irradiation
2.4. Immunohistochemistry
2.5. RNA Isolation, Real-Time qPCR and RNA-Seq Analysis
2.6. Measurement of The Maximum Width of The Optic Tectum (OT)
2.7. Statistical Analysis
3. Results
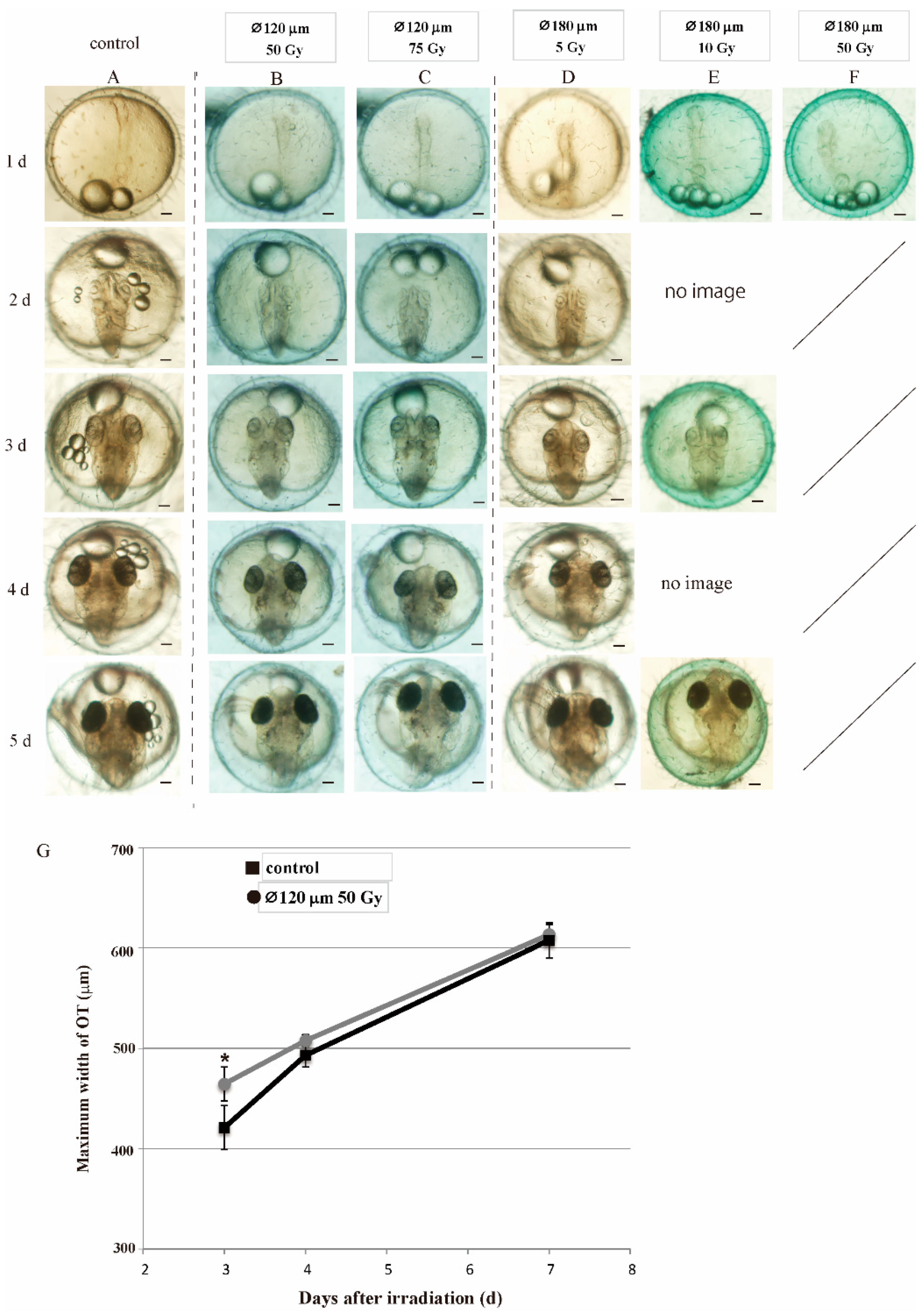
3.1. Susceptibility of Embryos to Irradiation Decreased as the Development Proceeds During Extremely Early Embryogenesis Period
3.2. Embryos Irradiated at Blastula Stage Showed Transient Delay of Brain Development but Normally Hatched
3.3. Normal Embryonic Development Depends on the Proportion of Non-Damaged Cells in Blastoderm of Irradiated Embryo
3.4. Irradiated Blastoderm Cells Did Not Arrest Cell Cycle Progression
3.5. Gamma-Ray Irradiation on the Embryos at Late Embryogenesis Period Resulted in Incomplete Development of Brain
4. Discussion
4.1. The Responses of Medaka Blastula Embryos after Irradiation Were Similar to Those of Pre-Implantation Period Embryos of Mammals
4.2. The Developmental Disturbance in IR Irradiated Embryos Might Be Determined by the Proportion of Severely Injured Cells
4.3. Brain Development Delayed Transiently in Irradiated Blastula Embryos Whereas the Irradiated Embryos during Late Embryogenesis Peiod Resulted in Incoplete Brain Development
4.4. Blastula Embryo Which Was Irradiated with a Lethal Dose IR Can Develop up to Neurula
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Russell, L.B.; Russell, W.L. An analysis of the changing radiation response of the developing mouse embryo. J. Cell. Physiol. Suppl. 1954, 43, 103–149. [Google Scholar] [CrossRef] [PubMed]
- Protection, R. ICRP Publication 26. Annals of the ICRP; Pergamon Press: Oxford, UK, 1977. [Google Scholar]
- De Santis, M.; Cesari, E.; Nobili, E.; Straface, G.; Cavaliere, A.F.; Caruso, A. Radiation effects on development. Birth Defects Res. Part Cembryo Today Rev. 2007, 81, 177–182. [Google Scholar] [CrossRef] [PubMed]
- Yasuda, T.; Aoki, K.; Matsumoto, A.; Maruyama, K.; Hyodo-Taguchi, Y.; Fushiki, S.; Ishikawa, Y. Radiation-induced brain cell death can be observed in living medaka embryos. J. Radiat. Res. 2006, 47, 295–303. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shima, A.; Mitani, H. Medaka as a research organism: Past, present and future. Mech. Dev. 2004, 121, 599–604. [Google Scholar] [CrossRef]
- Hong, Y.; Winkler, C.; Schartl, M. Pluripotency and differentiation of embryonic stem cell lines from the medakafish (Oryzias latipes). Mech. Dev. 1996, 60, 33–44. [Google Scholar] [CrossRef]
- Hong, Y.; Winkler, C.; Schartl, M. Efficiency of cell culture derivation from blastula embryos and of chimera formation in the medaka (Oryzias latipes) depends on donor genotype and passage number. Dev. Genes Evol. 1998, 208, 595–602. [Google Scholar] [CrossRef] [PubMed]
- Hong, N.; He, B.P.; Schartl, M.; Hong, Y. Medaka embryonic stem cells are capable of generating entire organs and embryo-like miniatures. Stem Cells Dev. 2013, 22, 750–757. [Google Scholar] [CrossRef]
- Funayama, T.; Wada, S.; Yokota, Y.; Fukamoto, K.; Sakashita, T.; Taguchi, M.; Kakizaki, T.; Hamada, N.; Suzuki, M.; Furusawa, Y.; et al. Heavy-ion microbeam system at JAEA-Takasaki for microbeam biology. J. Radiat. Res. 2008, 49, 71–82. [Google Scholar] [CrossRef] [Green Version]
- Funayama, T. Heavy-ion microbeams for biological science: Development of system and utilization for biological experiments in QST-Takasaki. Quantum Beam Sci. 2019, 3, 13. [Google Scholar] [CrossRef] [Green Version]
- Furusawa, T.; Fukamoto, K.; Sakashita, T.; Suzuki, E.; Kakizaki, T.; Hamada, N.; Funayama, T.; Suzuki, H.; Ishioka, N.; Wada, S.; et al. Targeted heavy-ion microbeam irradiation of the embryo but not yolk in the diapause-terminated egg of the silkworm, bombyx mori, induces the somatic mutation. J. Radiat. Res. 2009, 50, 371–375. [Google Scholar] [CrossRef]
- Yasuda, T.; Kamahori, M.; Nagata, K.; Watanabe-Asaka, T.; Suzuki, M.; Funayama, T.; Mitani, H.; Oda, S. Abscopal Activation of Microglia in Embryonic Fish Brain Following Targeted Irradiation with Heavy-Ion Microbeam. Int. J. Mol. Sci. 2017, 18, 1428. [Google Scholar] [CrossRef] [Green Version]
- Hyodo-Taguchi, Y.; Egami, N. Establishment of inbred strains of the medaka Oryzias latipes and the usefulness of the strains for biomedical-research. Zool. Sci. 1985, 2, 305–316. [Google Scholar]
- Iwamatsu, T. Stages of normal development in the medaka Oryzias latipes. Mech. Dev. 2004, 121, 605–618. [Google Scholar] [CrossRef] [PubMed]
- Yasuda, T.; Oda, S.; Hibi, Y.; Satoh, S.; Nagata, K.; Hirakawa, K.; Kutsuna, N.; Sagara, H.; Mitani, H. Embryonic Medaka Model of Microglia in the Developing CNS Allowing In Vivo Analysis of Their Spatiotemporal Recruitment in Response to Irradiation. PLoS ONE 2015, 10, e0127325. [Google Scholar] [CrossRef] [Green Version]
- Suzuki, M.; Hattori, Y.; Sakashita, T.; Yokota, Y.; Kobayashi, Y.; Funayama, T. Region-specific irradiation system with heavy-ion microbeam for active individuals of Caenorhabditis elegans. J. Radiat. Res. 2017, 58, 881–886. [Google Scholar] [CrossRef]
- Suzuki, M.; Sakashita, T.; Funayama, T. Immobilization of Live Caenorhabditis elegans Individuals Using an Ultra-thin Polydimethylsiloxane Microfluidic Chip with Water Retention. J. Vis. Exp. JoVE 2019, 145, e59008. [Google Scholar] [CrossRef] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- ICRP. The 2007 Recommendations of the International Commission on Radiological Protection. ICRP Publication 103. Ann. ICRP 2007, 37, 2. [Google Scholar]
- Claveria, C.; Giovinazzo, G.; Sierra, R.; Torres, M. Myc-driven endogenous cell competition in the early mammalian embryo. Nature 2013, 500, 39–44. [Google Scholar] [CrossRef]
- Sancho, M.; Di-Gregorio, A.; George, N.; Pozzi, S.; Sanchez, J.M.; Pernaute, B.; Rodriguez, T.A. Competitive Interactions Eliminate Unfit Embryonic Stem Cells at the Onset of Differentiation. Dev. Cell 2013, 26, 19–30. [Google Scholar] [CrossRef] [Green Version]
- Maruyama, T.; Fujita, Y. Cell competition in mammals—Novel homeostatic machinery for embryonic development and cancer prevention. Curr. Opin. Cell Biol. 2017, 48, 106–112. [Google Scholar] [CrossRef] [Green Version]
- Morata, G.; Ripoll, P. Minutes: Mutants of drosophila autonomously affecting cell division rate. Dev. Biol. 1975, 42, 211–221. [Google Scholar] [CrossRef]
- Paranjpe, S.S.; Veenstra, G.J. Establishing pluripotency in early development. Biochim. Biophys. Acta 2015, 1849, 626–636. [Google Scholar] [CrossRef] [Green Version]
- Nowak, E.; Etienne, O.; Millet, P.; Lages, C.S.; Mathieu, C.; Mouthon, M.A.; Boussin, F.D. Radiation-induced H2AX phosphorylation and neural precursor apoptosis in the developing brain of mice. Radiat. Res. 2006, 165, 155–164. [Google Scholar] [CrossRef]
- Shimada, M.; Matsuzaki, F.; Kato, A.; Kobayashi, J.; Matsumoto, T.; Komatsu, K. Induction of Excess Centrosomes in Neural Progenitor Cells during the Development of Radiation-Induced Microcephaly. PLoS ONE 2016, 11, e0158236. [Google Scholar] [CrossRef]
- Schull, W.J.; Otake, M. Cognitive function and prenatal exposure to ionizing radiation. Teratology 1999, 59, 222–226. [Google Scholar] [CrossRef]
- Kage, T.; Takeda, H.; Yasuda, T.; Maruyama, K.; Yamamoto, N.; Yoshimoto, M.; Araki, K.; Inohaya, K.; Okamoto, H.; Yasumasu, S.; et al. Morphogenesis and regionalization of the medaka embryonic brain. J. Comp. Neurol. 2004, 476, 219–239. [Google Scholar] [CrossRef]
- Aizawa, K.; Mitani, H.; Kogure, N.; Shimada, A.; Hirose, Y.; Sasado, T.; Morinaga, C.; Yasuoka, A.; Yoda, H.; Watanabe, T.; et al. Identification of radiation-sensitive mutants in the Medaka, Oryzias latipes. Mech. Dev. 2004, 121, 895–902. [Google Scholar] [CrossRef]
- Yasuda, T.; The University of Tokyo, Chiba, Japan. Unpublished work. 2015.
- Aizawa, K.; Shimada, A.; Naruse, K.; Mitani, H.; Shima, A. The medaka midblastula transition as revealed by the expression of the paternal genome. Gene Expr. Patterns 2003, 3, 43–47. [Google Scholar] [CrossRef]
- Aizawa, K.; Yori, K.; Kaminaga, C.; Yamashita, T.; Kinoshita, M.; Oda, S.; Mitani, H. Responses of embryonic germ cells of the radiation-sensitive Medaka mutant to gamma-irradiation. J. Radiat. Res. 2007, 48, 121–128. [Google Scholar] [CrossRef]
- Stack, J.H.; Newport, J.W. Developmentally regulated activation of apoptosis early in Xenopus gastrulation results in cyclin A degradation during interphase of the cell cycle. Development 1997, 124, 3185–3195. [Google Scholar]
- Ikegami, R.; Rivera-Bennetts, A.K.; Brooker, D.L.; Yager, T.D. Effect of inhibitors of DNA replication on early zebrafish embryos: Evidence for coordinate activation of multiple intrinsic cell-cycle checkpoints at the mid-blastula transition. Zygote 1997, 5, 153–175. [Google Scholar] [CrossRef]
- Ikegami, R.; Hunter, P.; Yager, T.D. Developmental activation of the capability to undergo checkpoint-induced apoptosis in the early zebrafish embryo. Dev. Biol. 1999, 209, 409–433. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Langley, A.R.; Smith, J.C.; Stemple, D.L.; Harvey, S.A. New insights into the maternal to zygotic transition. Development 2014, 141, 3834–3841. [Google Scholar] [CrossRef] [Green Version]
- Honjo, Y.; Ichinohe, T. Cellular responses to ionizing radiation change quickly over time during early development in zebrafish. Cell Biol. Int. 2019, 43, 516–527. [Google Scholar] [CrossRef] [Green Version]
- el-Deiry, W.S.; Tokino, T.; Velculescu, V.E.; Levy, D.B.; Parsons, R.; Trent, J.M.; Lin, D.; Mercer, W.E.; Kinzler, K.W.; Vogelstein, B. WAF1, a potential mediator of p53 tumor suppression. Cell 1993, 75, 817–825. [Google Scholar] [CrossRef]
- Cayrol, C.; Knibiehler, M.; Ducommun, B. p21 binding to PCNA causes G1 and G2 cell cycle arrest in p53-deficient cells. Oncogene 1998, 16, 311–320. [Google Scholar] [CrossRef] [Green Version]
- te Poele, R.H.; Okorokov, A.L.; Jardine, L.; Cummings, J.; Joel, S.P. DNA damage is able to induce senescence in tumor cells in vitro and in vivo. Cancer Res. 2002, 62, 1876–1883. [Google Scholar]
- Campisi, J.; d’Adda di Fagagna, F. Cellular senescence: When bad things happen to good cells. Nat. Rev. Mol. Cell Biol. 2007, 8, 729–740. [Google Scholar] [CrossRef]
- Purcell, M.; Kruger, A.; Tainsky, M.A. Gene expression profiling of replicative and induced senescence. Cell Cycle 2014, 13, 3927–3937. [Google Scholar] [CrossRef] [Green Version]
- Casella, G.; Munk, R.; Kim, K.M.; Piao, Y.; De, S.; Abdelmohsen, K.; Gorospe, M. Transcriptome signature of cellular senescence. Nucleic Acids Res. 2019, 47, 11476. [Google Scholar] [CrossRef] [PubMed]
- Solozobova, V.; Rolletschek, A.; Blattner, C. Nuclear accumulation and activation of p53 in embryonic stem cells after DNA damage. BMC Cell Biol. 2009, 10, 46. [Google Scholar] [CrossRef] [Green Version]
- Filion, T.M.; Qiao, M.; Ghule, P.N.; Mandeville, M.; van Wijnen, A.J.; Stein, J.L.; Lian, J.B.; Altieri, D.C.; Stein, G.S. Survival responses of human embryonic stem cells to DNA damage. J. Cell. Physiol. 2009, 220, 586–592. [Google Scholar] [CrossRef] [Green Version]
- Itahana, Y.; Zhang, J.; Goke, J.; Vardy, L.A.; Han, R.; Iwamoto, K.; Cukuroglu, E.; Robson, P.; Pouladi, M.A.; Colman, A.; et al. Histone modifications and p53 binding poise the p21 promoter for activation in human embryonic stem cells. Sci. Rep. 2016, 6, 28112. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene ID | Gene Name | Discription | Fold Change IR Embryos/Sham-Cont. |
|---|---|---|---|
| ENSORLG00000016085 | hipk2 | homeodomain interacting protein kinase 2 [Source:ZFIN;Acc:ZDB-GENE-031125-4] | −1.40 |
| ENSORLG00000010268 | msh2 | mutS homolog 2 (E. coli) [Source:ZFIN; Acc:ZDB-GENE-040426-2932] | −1.32 |
| ENSORLG00000016313 | si:ch211-160o17.4 | si:ch211-160o17.4 [Source:ZFIN; Acc:ZDB-GENE-141215-36] | −1.15 |
| ENSORLG00000022951 | si:dkey-74k8.3 | si:dkey-74k8.3 [Source:ZFIN; Acc:ZDB-GENE-141222-20] | −1.07 |
| ENSORLG00000012316 | not defined | homeodomain-interacting protein kinase 3 [Source:NCBI gene;Acc:101162288] | 1.06 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yasuda, T.; Funayama, T.; Nagata, K.; Li, D.; Endo, T.; Jia, Q.; Suzuki, M.; Ishikawa, Y.; Mitani, H.; Oda, S. Collimated Microbeam Reveals that the Proportion of Non-Damaged Cells in Irradiated Blastoderm Determines the Success of Development in Medaka (Oryzias latipes) Embryos. Biology 2020, 9, 447. https://doi.org/10.3390/biology9120447
Yasuda T, Funayama T, Nagata K, Li D, Endo T, Jia Q, Suzuki M, Ishikawa Y, Mitani H, Oda S. Collimated Microbeam Reveals that the Proportion of Non-Damaged Cells in Irradiated Blastoderm Determines the Success of Development in Medaka (Oryzias latipes) Embryos. Biology. 2020; 9(12):447. https://doi.org/10.3390/biology9120447
Chicago/Turabian StyleYasuda, Takako, Tomoo Funayama, Kento Nagata, Duolin Li, Takuya Endo, Qihui Jia, Michiyo Suzuki, Yuji Ishikawa, Hiroshi Mitani, and Shoji Oda. 2020. "Collimated Microbeam Reveals that the Proportion of Non-Damaged Cells in Irradiated Blastoderm Determines the Success of Development in Medaka (Oryzias latipes) Embryos" Biology 9, no. 12: 447. https://doi.org/10.3390/biology9120447
APA StyleYasuda, T., Funayama, T., Nagata, K., Li, D., Endo, T., Jia, Q., Suzuki, M., Ishikawa, Y., Mitani, H., & Oda, S. (2020). Collimated Microbeam Reveals that the Proportion of Non-Damaged Cells in Irradiated Blastoderm Determines the Success of Development in Medaka (Oryzias latipes) Embryos. Biology, 9(12), 447. https://doi.org/10.3390/biology9120447