Inhibitory Effects of Pulse Bioactive Compounds on Cancer Development Pathways

 , ,
, ,  and
and
Abstract
:
1. Introduction
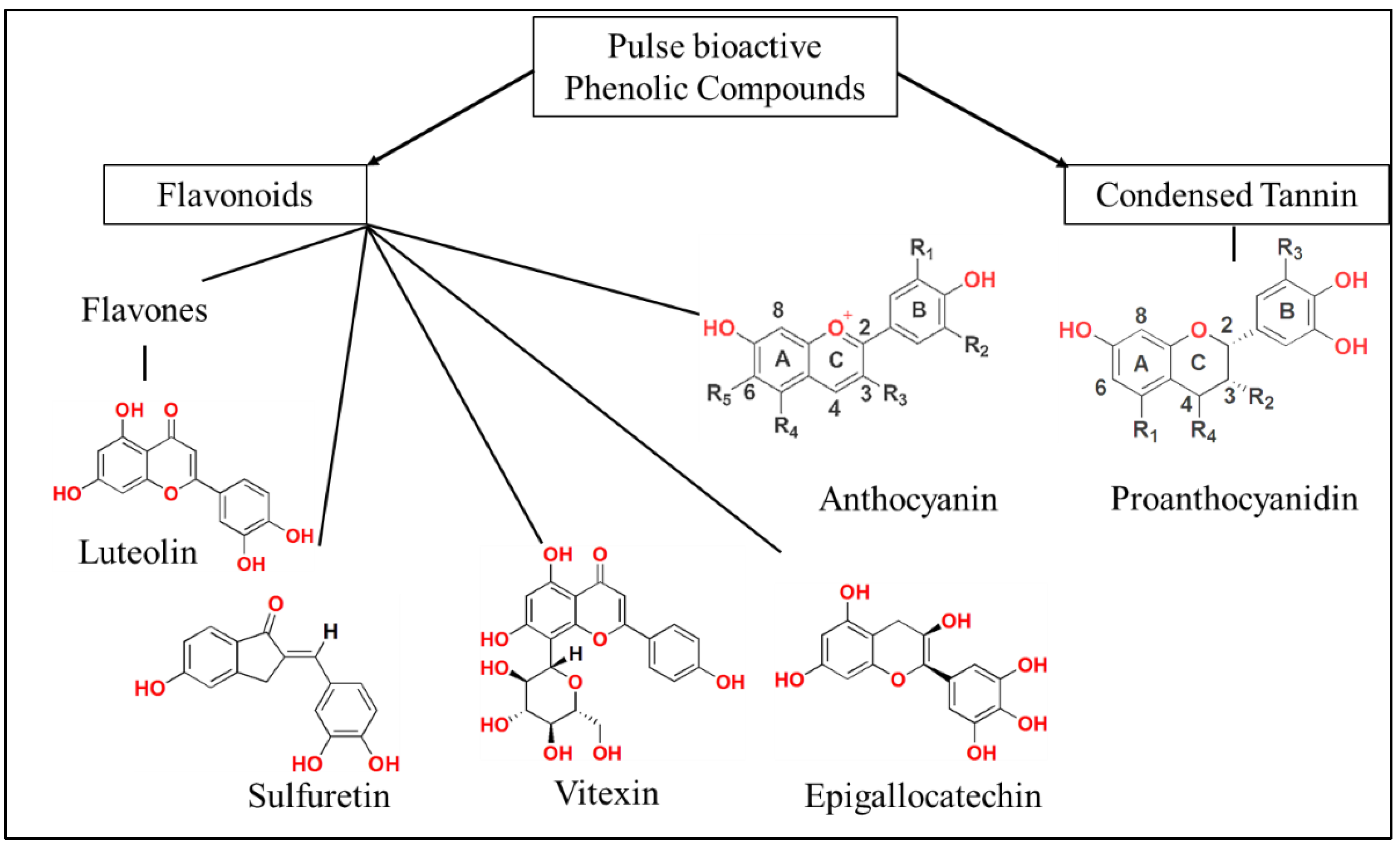
2. Bioactive Compounds in Pulses
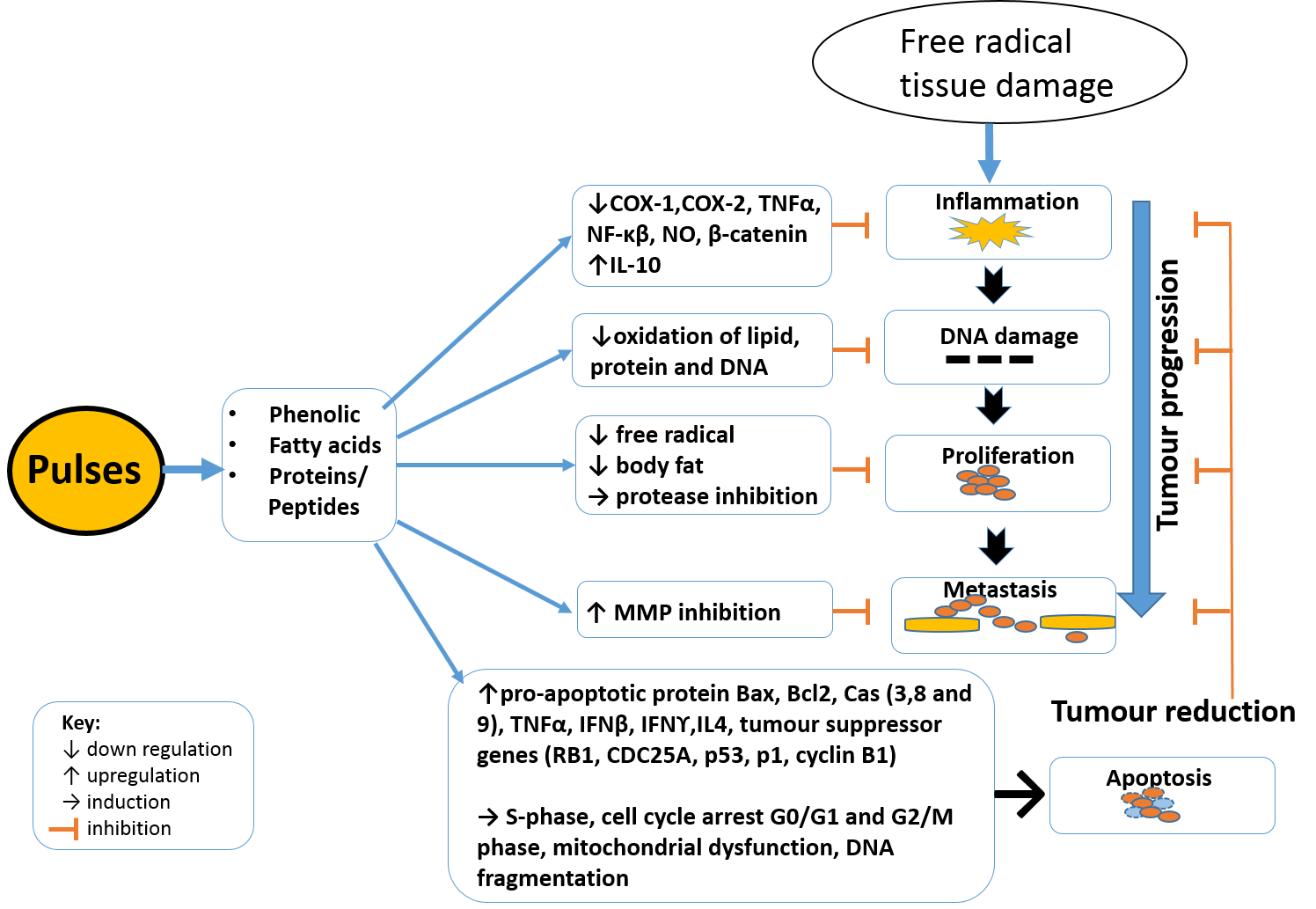
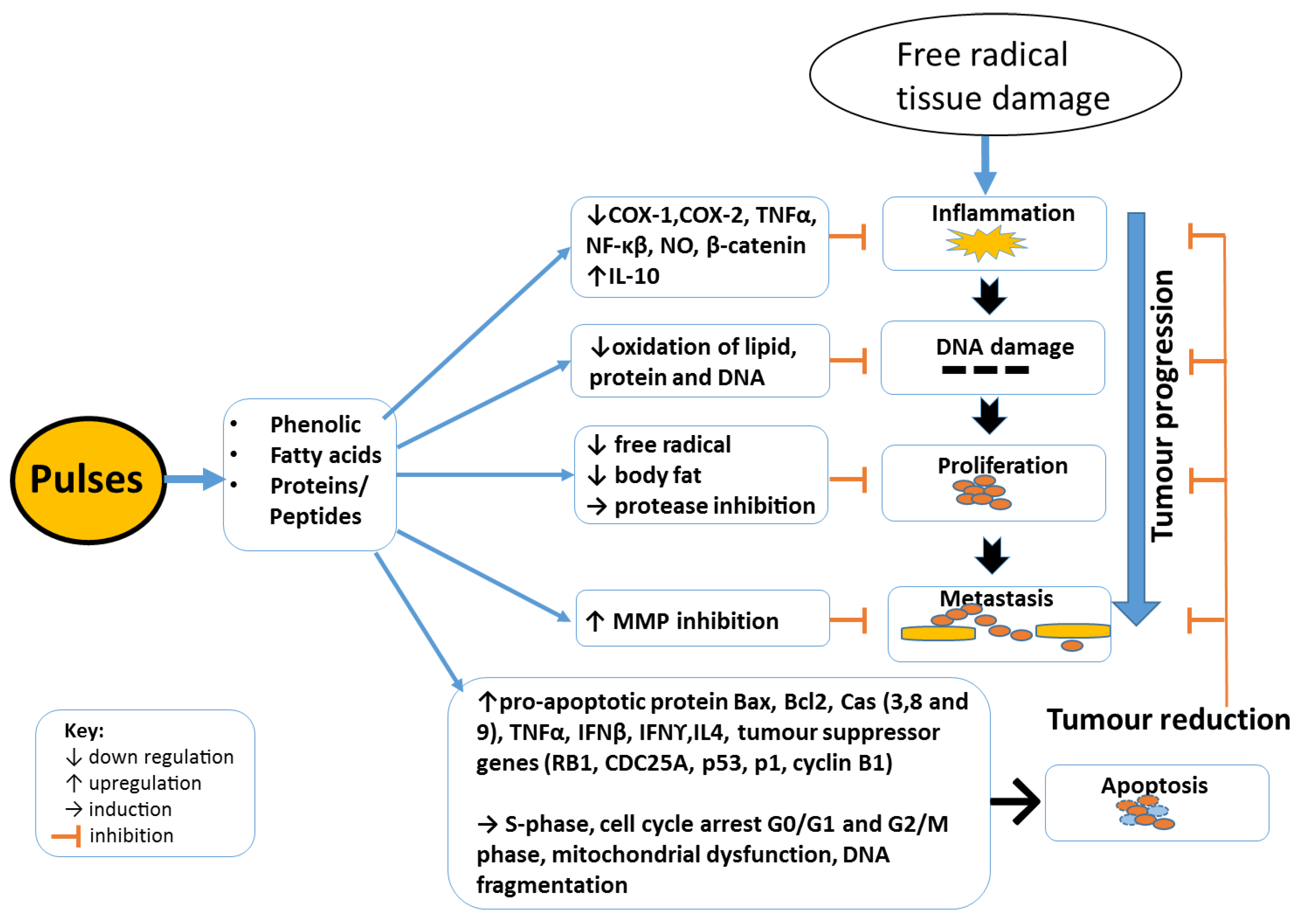
3. Cancer Development and Mechanisms of Inhibition
3.1. Anti-Inflammation
3.1.1. Role of Pulse Phenolic Compounds
3.1.2. Role of Pulse Bioactive Peptides and Amino Acids
3.2. Anti-Proliferation
3.2.1. Role of Pulse Phenolic Compounds
3.2.2. Role of Pulse Bioactive Proteins
3.2.3. Role of Short-Chain Fatty Acids
3.3. Pro-Apoptotic Effects of Pulses
3.3.1. Pro-Apoptotic Bioactive Phenolic Compounds
3.3.2. Pro-Apoptotic Bioactive Peptides
3.3.3. Pro-Apoptotic Pulse Short-Chain Fatty Acids
3.4. Anti-Metastatic Effects of Pulses
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Food and Agriculture Organisation. Pulses and Derived Products. Available online: http://www.fao.org/es/faodef/fdef04e.htm (accessed on 30 June 2018).
- Yao, Y.; Yang, X.; Tian, J.; Liu, C.; Cheng, X.; Ren, G. Antioxidant and Antidiabetic Activities of Black Mung Bean (Vigna radiata L.). J. Agric. Food Chem. 2013, 61, 8104–8109. [Google Scholar] [CrossRef] [PubMed]
- Reverri, E.J.; Randolph, J.M.; Kappagoda, C.T.; Park, E.; Edirisinghe, I.; Burton-Freeman, B.M. Assessing beans as a source of intrinsic fiber on satiety in men and women with metabolic syndrome. Appetite 2017, 118, 75–81. [Google Scholar] [CrossRef] [PubMed]
- Millán-Linares, M.d.C.; Bermúdez, B.; Yust, M.d.M.; Millán, F.; Pedroche, J. Anti-inflammatory activity of lupine (Lupinus angustifolius L.) protein hydrolysates in THP-1-derived macrophages. J. Funct. Foods 2014, 8, 224–233. [Google Scholar] [CrossRef]
- Jang, D.S.; Park, E.J.; Hawthorne, M.E.; Vigo, J.S.; Graham, J.G.; Cabieses, F.; Santarsiero, B.D.; Mesecar, A.D.; Fong, H.H.S.; Mehta, R.G.; et al. Potential Cancer Chemopreventive Constituents of the Seeds of Dipteryx odorata (Tonka Bean). J. Nat. Prod. 2003, 66, 583–587. [Google Scholar] [CrossRef] [PubMed]
- Siah, S.D.; Konczak, I.; Agboola, S.; Wood, J.A.; Blanchard, C.L. In vitro investigations of the potential health benefits of Australian-grown faba beans (Vicia faba L.): Chemopreventative capacity and inhibitory effects on the angiotensin-converting enzyme, α-glucosidase and lipase. Br. J. Nutr. 2012, 108, S123–S134. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Chino, X.; Jiménez-Martínez, C.; Dávila-Ortiz, G.; Álvarez-González, I.; Madrigal-Bujaidar, E. Nutrient and Nonnutrient Components of Legumes, and Its Chemopreventive Activity: A Review. Nutr. Cancer 2015, 67, 401–410. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Shang, P.; Qin, F.; Zhou, Q.; Gao, B.; Huang, H.; Yang, H.; Shi, H.; Yu, L. Chemical composition and antioxidative and anti-inflammatory properties of ten commercial mung bean samples. LWT—Food Sci. Technol. 2013, 54, 171–178. [Google Scholar] [CrossRef]
- Dong, M.; He, X.; Liu, R.H. Phytochemicals of Black Bean Seed Coats: Isolation, Structure Elucidation, and Their Antiproliferative and Antioxidative Activities. J. Agric. Food Chem. 2007, 55, 6044–6051. [Google Scholar] [CrossRef] [PubMed]
- Yun-Kyoung, L.; Jin-Taek, H.; Mee-Sook, L.; Young-Min, K.; Jin, P.O. Kidney Bean Husk Extracts Exert Antitumor Effect by Inducing Apoptosis Involving AMP-Activated Protein Kinase Signaling Pathway. Ann. N. Y. Acad. Sci. 2009, 1171, 484–488. [Google Scholar]
- Cruz-Bravo, R.K.; Guevara-Gonzalez, R.; Ramos-Gomez, M.; Garcia-Gasca, T.; Campos-Vega, R.; Oomah, B.D.; Loarca-Piña, G. Fermented Nondigestible Fraction from Common Bean (Phaseolus vulgaris L.) Cultivar Negro 8025 Modulates HT-29 Cell Behavior. J. Food Sci. 2011, 76, T41–T47. [Google Scholar] [CrossRef] [PubMed]
- Sosulski, F.W.; Dabrowski, K.J. Composition of Free and Hydrolyzable Phenolic-Acids in the Flours and Hulls of 10 Legume Species. J. Agric. Food Chem. 1984, 32, 131–133. [Google Scholar] [CrossRef]
- Giusti, F.; Caprioli, G.; Ricciutelli, M.; Vittori, S.; Sagratini, G. Determination of fourteen polyphenols in pulses by high performance liquid chromatography-diode array detection (HPLC-DAD) and correlation study with antioxidant activity and colour. Food Chem. 2017, 221, 689–697. [Google Scholar] [CrossRef] [PubMed]
- Boudjou, S.; Oomah, B.D.; Zaidi, F.; Hosseinian, F. Phenolics content and antioxidant and anti-inflammatory activities of legume fractions. Food Chem. 2013, 138, 1543–1550. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Mora, P.; Frias, J.; Peñas, E.; Zieliński, H.; Giménez-Bastida, J.A.; Wiczkowski, W.; Zielińska, D.; Martínez-Villaluenga, C. Simultaneous release of peptides and phenolics with antioxidant, ACE-inhibitory and anti-inflammatory activities from pinto bean (Phaseolus vulgaris L. var. pinto) proteins by subtilisins. J. Funct. Foods 2015, 18, 319–332. [Google Scholar] [CrossRef]
- Kan, L.; Nie, S.; Hu, J.; Liu, Z.; Xie, M. Antioxidant activities and anthocyanins composition of seed coats from twenty-six kidney bean cultivars. J. Funct. Foods 2016, 26, 622–631. [Google Scholar] [CrossRef]
- Hua, C.; Hai-Rong, M.; Yan-Hua, G.; Xue, Z.; Madina, H.; Rui, H.; Akber, A.H. Isoflavones Extracted from Chickpea Cicer arietinum L. Sprouts Induce Mitochondria-Dependent Apoptosis in Human Breast Cancer Cells. Phytother. Res. 2015, 29, 210–219. [Google Scholar]
- Segura-Nieto, M.; Jim, E.R. Genetic Modifications of Plant Seed Storage Proteins for Food Production; Technomic Publishing: Lancaster, PA, USA, 1999. [Google Scholar]
- Sharon, N.; Lis, H. Legume lectins—A large family of homologous proteins. FASEB J. 1990, 4, 3198–3208. [Google Scholar] [CrossRef] [PubMed]
- Laskowski, M., Jr.; Kato, I. Protein Inhibitors of Proteinases. Ann. Rev. Biochem. 1980, 49, 593–626. [Google Scholar] [CrossRef] [PubMed]
- Padhi, E.M.T.; Liu, R.; Hernandez, M.; Tsao, R.; Ramdath, D.D. Total polyphenol content, carotenoid, tocopherol and fatty acid composition of commonly consumed Canadian pulses and their contribution to antioxidant activity. J. Funct. Foods 2017, 38, 602–611. [Google Scholar] [CrossRef]
- Kan, L.; Nie, S.; Hu, J.; Wang, S.; Bai, Z.; Wang, J.; Zhou, Y.; Jiang, J.; Zeng, Q.; Song, K. Comparative study on the chemical composition, anthocyanins, tocopherols and carotenoids of selected legumes. Food Chem. 2018, 260, 317–326. [Google Scholar] [CrossRef] [PubMed]
- Cooper, G.M. The Development and Causes of Cancer, 2nd ed.; Sinauer Associates: Sunderland, MA, USA, 2000. [Google Scholar]
- Aparicio-Fernández, X.; Reynoso-Camacho, R.; Castaño-Tostado, E.; García-Gasca, T.; González de Mejía, E.; Guzmán-Maldonado, S.H.; Elizondo, G.; Yousef, G.G.; Lila, M.A.; Loarca-Pina, G. Antiradical Capacity and Induction of Apoptosis on HeLa Cells by a Phaseolus vulgaris Extract. Plant Foods Hum. Nutr. 2008, 63, 35–40. [Google Scholar] [CrossRef] [PubMed]
- Mudryj, A.N.; Yu, N.; Aukema, H.M. Nutritional and health benefits of pulses. Appl. Physiol. Nutr. Metab. 2014, 39, 1197–1204. [Google Scholar] [CrossRef] [PubMed]
- Guadagni, F.; Ferroni, P.; Palmirotta, R.; Portarena, I.; Formica, V.; Roselli, M. Review. TNF/VEGF cross-talk in chronic inflammation-related cancer initiation and progression: An early target in anticancer therapeutic strategy. In Vivo 2007, 21, 147–161. [Google Scholar] [PubMed]
- Limtrakul, P.; Yodkeeree, S.; Pitchakarn, P.; Punfa, W. Suppression of Inflammatory Responses by Black Rice Extract in RAW 264.7 Macrophage Cells via Downregulation of NF-kB and AP-1 Signaling Pathways. Asian Pac. J. Cancer Prev. 2015, 16, 4277–4283. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gilmore, T.D. Introduction to NF-kappaB: Players, pathways, perspectives. Oncogene 2006, 25, 6680–6684. [Google Scholar] [CrossRef] [PubMed]
- Mantovani, A.; Garlanda, C.; Allavena, P. Molecular pathways and targets in cancer-related inflammation. Ann. Med. 2010, 42, 161–170. [Google Scholar] [CrossRef] [PubMed]
- Mantovani, A.; Allavena, P.; Sica, A.; Balkwill, F. Cancer-related inflammation. Nature 2008, 454, 436–444. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luo, J.; Cai, W.; Wu, T.; Xu, B. Phytochemical distribution in hull and cotyledon of adzuki bean (Vigna angularis L.) and mung bean (Vigna radiate L.), and their contribution to antioxidant, anti-inflammatory and anti-diabetic activities. Food Chem. 2016, 201, 350–360. [Google Scholar] [CrossRef] [PubMed]
- Moreno-Jiménez, M.R.; Cervantes-Cardoza, V.; Gallegos-Infante, J.A.; González-Laredo, R.F.; Estrella, I.; García-Gasca, T.d.J.; Herrera-Carrera, E.; Díaz-Rivas, J.O.; Rocha-Guzmán, N.E. Phenolic composition changes of processed common beans: Their antioxidant and anti-inflammatory effects in intestinal cancer cells. Food Res. Int. 2015, 76, 79–85. [Google Scholar] [CrossRef]
- López-Barrios, L.; Antunes-Ricardo, M.; Gutiérrez-Uribe, J.A. Changes in antioxidant and antiinflammatory activity of black bean (Phaseolus vulgaris L.) protein isolates due to germination and enzymatic digestion. Food Chem. 2016, 203, 417–424. [Google Scholar] [CrossRef] [PubMed]
- Yeap, S.K.; Mohd Yusof, H.; Mohamad, N.E.; Beh, B.K.; Ho, W.Y.; Ali, N.M.; Alitheen, N.B.; Koh, S.P.; Long, K. In Vivo Immunomodulation and Lipid Peroxidation Activities Contributed to Chemoprevention Effects of Fermented Mung Bean against Breast Cancer. Evid.-Based Complement. Altern. Med. 2013, 2013, 708464. [Google Scholar] [CrossRef] [PubMed]
- Thompson, M.D.; Brick, M.A.; McGinley, J.N.; Thompson, H.J. Chemical Composition and Mammary Cancer Inhibitory Activity of Dry Bean All rights reserved. No part of this periodical may be reproduced or transmitted in any form or by any means, electronic or mechanical, including photocopying, recording, or any information storage and retrieval system, without permission in writing from the publisher. Permission for printing and for reprinting the material contained herein has been obtained by the publisher. Crop Sci. 2009, 49, 179–186. [Google Scholar]
- García-Lafuente, A.; Moro, C.; Manchón, N.; Gonzalo-Ruiz, A.; Villares, A.; Guillamón, E.; Rostagno, M.; Mateo-Vivaracho, L. In vitro anti-inflammatory activity of phenolic rich extracts from white and red common beans. Food Chem. 2014, 161, 216–223. [Google Scholar] [CrossRef] [PubMed]
- Sánchez, X.; Martinez, C.J.; Garzón, V.R.V.; Álvarez González, I.; Villa-Treviño, S.; Madrigal Bujaidar, E.; Dávila Ortiz, G.; Baltiérrez Hoyos, R. Cooked Chickpea Consumption Inhibits Colon Carcinogenesis in Mice Induced with Azoxymethane and Dextran Sulfate Sodium. J. Am. Coll. Nutr. 2017, 36, 391–398. [Google Scholar]
- Xu, B.; Chang, S.K.C. Comparative study on antiproliferation properties and cellular antioxidant activities of commonly consumed food legumes against nine human cancer cell lines. Food Chem. 2012, 134, 1287–1296. [Google Scholar] [CrossRef] [PubMed]
- Niveditha, V.R.; Venkatramana, D.K.; Sridhar, K.R. Cytotoxic effects of methanol extract of raw, cooked and fermented split beans of Canavalia on cancer cell lines MCF-7 and HT-29. IIOAB J. 2013, 4, 20–23. [Google Scholar]
- Kim, D.K.; Jeong, S.C.; Gorinstein, S.; Chon, S.-U. Total Polyphenols, Antioxidant and Antiproliferative Activities of Different Extracts in Mungbean Seeds and Sprouts. Plant Foods Hum. Nutr. 2012, 67, 71–75. [Google Scholar] [CrossRef] [PubMed]
- Ombra, M.N.; D’Acierno, A.; Nazzaro, F.; Riccardi, R.; Spigno, P.; Zaccardelli, M.; Pane, C.; Maione, M.; Fratianni, F. Phenolic Composition and Antioxidant and Antiproliferative Activities of the Extracts of Twelve Common Bean (Phaseolus vulgaris L.) Endemic Ecotypes of Southern Italy before and after Cooking. Oxid. Med. Cell. Longev. 2016, 2016, 1398298. [Google Scholar] [CrossRef] [PubMed]
- Zou, Y.P.; Chang, K.-C. Antioxidant and Antiproliferative Properties of Extract and Fractions from Small Red Bean (Phaseolus vulgaris L.). J. Food Nutr. 2014. [Google Scholar] [CrossRef]
- Chen, Z.; Wang, J.; Liu, W.; Chen, H. Physicochemical characterization, antioxidant and anticancer activities of proteins from four legume species. J. Food Sci. Technol. 2017, 54, 964–972. [Google Scholar] [CrossRef] [PubMed]
- Ma, D.Z.; Wang, H.X.; Ng, T.B. A peptide with potent antifungal and antiproliferative activities from Nepalese large red beans. Peptides 2009, 30, 2089–2094. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.X.; Ng, T.B. Isolation and characterization of an antifungal peptide with antiproliferative activity from seeds of Phaseolus vulgaris cv. ‘Spotted Bean’. Appl. Microbiol. Biotechnol. 2007, 74, 125–130. [Google Scholar] [CrossRef] [PubMed]
- Magee, P.J.; Owusu-Apenten, R.; McCann, M.J.; Gill, C.I.; Rowland, I.R. Chickpea (Cicer arietinum) and Other Plant-Derived Protease Inhibitor Concentrates Inhibit Breast and Prostate Cancer Cell Proliferation In Vitro. Nutr. Cancer 2012, 64, 741–748. [Google Scholar] [CrossRef] [PubMed]
- Joghatai, M.; Barari, L.; Mousavie Anijdan, S.H.; Elmi, M.M. The evaluation of radio-sensitivity of mung bean proteins aqueous extract on MCF-7, hela and fibroblast cell line. Int. J. Radiat. Biol. 2018, 94, 478–487. [Google Scholar] [CrossRef] [PubMed]
- Lam, S.K.; Ng, T.B. Isolation and characterization of a French bean hemagglutinin with antitumor, antifungal, and anti-HIV-1 reverse transcriptase activities and an exceptionally high yield. Phytomedicine 2010, 17, 457–462. [Google Scholar] [CrossRef] [PubMed]
- Wong, J.H.; Ng, T.B. A homotetrameric agglutinin with antiproliferative and mitogenic activities from haricot beans. J. Chromatogr. B-Anal. Technol. Biomed. Life Sci. 2005, 828, 130–135. [Google Scholar]
- Wang, S.; Rao, P.; Ye, X. Isolation and biochemical characterization of a novel leguminous defense peptide with antifungal and antiproliferative potency. Appl. Microbiol. Biotechnol. 2009, 82, 79–86. [Google Scholar] [CrossRef] [PubMed]
- Ngai, P.H.K.; Ng, T.B. Coccinin, an antifungal peptide with antiproliferative and HIV-1 reverse transcriptase inhibitory activities from large scarlet runner beans. Peptides 2004, 25, 2063–2068. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Lin, J.; Ye, M.; Ng, T.B.; Rao, P.; Ye, X. Isolation and characterization of a novel mung bean protease inhibitor with antipathogenic and anti-proliferative activities. Peptides 2006, 27, 3129–3136. [Google Scholar] [CrossRef] [PubMed]
- Ang, A.S.W.; Cheung, R.C.F.; Dan, X.; Chan, Y.S.; Pan, W.; Ng, T.B. Purification and Characterization of a Glucosamine-Binding Antifungal Lectin from Phaseolus vulgaris cv. Chinese Pinto Beans with Antiproliferative Activity Towards Nasopharyngeal Carcinoma Cells. Appl. Biochem. Biotechnol. 2014, 172, 672–686. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.; Wang, H.; Ng, T.B. Trypsin Isoinhibitors with Antiproliferative Activity toward Leukemia Cells from Phaseolus vulgaris cv White Cloud Bean. J. Biomed. Biotechnol. 2010, 2010, 219793. [Google Scholar] [CrossRef] [PubMed]
- Clemente, A.; Carmen Marín-Manzano, M.; Jiménez, E.; Carmen Arques, M.; Domoney, C. The anti-proliferative effect of TI1B, a major Bowman–Birk isoinhibitor from pea (Pisum sativum L.), on HT29 colon cancer cells is mediated through protease inhibition. Br. J. Nutr. 2012, 108, S135–S144. [Google Scholar] [CrossRef] [PubMed]
- Barari, L.; Abedian, Z.; Asadi, A.; Elmi, F.; Elmi, M. Anti-proliferative Effect of Chickpea Extract on Hela, MCF-7, Saos and Fibroblast Cell Lines. Br. J. Pharm. Res. 2017, 15, 1–9. [Google Scholar] [CrossRef]
- Kumar, S.; Kapoor, V.; Gill, K.; Singh, K.; Xess, I.; Das, S.N.; Dey, S. Antifungal and Antiproliferative Protein from Cicer arietinum: A Bioactive Compound against Emerging Pathogens. BioMed Res. Int. 2014, 2014, 387203. [Google Scholar] [CrossRef] [PubMed]
- Loizzo, M.R.; Rashed, K.; Said, A.; Bonesi, M.; Menichini, F.; Tundis, R. Antiproliferative and antioxidant properties of Alhagi maurorum Boiss (Leguminosae) aerial parts. Ind. Crops Prod. 2014, 53, 289–295. [Google Scholar] [CrossRef]
- Bennink, M.R. Consumption of Black Beans and Navy Beans (Phaseolus vulgaris) Reduced Azoxymethane-Induced Colon Cancer in Rats. Nutr. Cancer 2002, 44, 60–65. [Google Scholar] [CrossRef] [PubMed]
- Aparicio-Fernández, X.; García-Gasca, T.; Yousef, G.G.; Lila, M.A.; González de Mejia, E.; Loarca-Pina, G. Chemopreventive Activity of Polyphenolics from Black Jamapa Bean (Phaseolus vulgaris L.) on HeLa and HaCaT Cells. J. Agric. Food Chem. 2006, 54, 2116–2122. [Google Scholar] [CrossRef] [PubMed]
- Hafidh, R.R.; Abdulamir, A.S.; Bakar, F.A.; Jalilian, F.A.; Abas, F.; Sekawi, Z. Novel molecular, cytotoxical, and immunological study on promising and selective anticancer activity of Mung bean sprouts. BMC Complement. Altern. Med. 2012, 12, 208. [Google Scholar] [CrossRef] [PubMed]
- Busambwa, K.; Sunkara, R.; Diby, N.; Offei-Okyne, R.; Boateng, J.; Verghese, M. Cytotoxic and Apoptotic Effects of Sprouted and Non-sprouted Lentil, Green and Yellow Split-peas. Int. J. Cancer Res. 2016, 12, 51–60. [Google Scholar]
- Thompson, M.D.; Thompson, H.J.; Brick, M.A.; McGinley, J.N.; Jiang, W.; Zhu, Z.; Wolfe, P. Mechanisms Associated with Dose-Dependent Inhibition of Rat Mammary Carcinogenesis by Dry Bean (Phaseolus vulgaris, L.). J. Nutr. 2008, 138, 2091–2097. [Google Scholar] [CrossRef] [PubMed]
- Aregueta-Robles, U.; Fajardo-Ramirez, O.R.; Villela, L.; Gutierrez-Uribe, J.A.; Hernandez-Hernandez, J.; Lopez-Sanchez, R.D.; Scott, S.P.; Serna-Saldivar, S. Cytotoxic Activity of a Black Bean (Phaseolus Vulgaris L.) Extract and Its Flavonoid Fraction in Both in Vitro and in Vivo Models of Lymphoma. Rev. Investig. Clin.-Clin. Transl. Investig. 2018, 70, 32–39. [Google Scholar] [CrossRef] [PubMed]
- Stanisavljević, N.S.; Ilić, M.D.; Matić, I.Z.; Jovanović, Ž.S.; Čupić, T.; Dabić, D.Č.; Natić, M.M.; Tešić, Ž.L. Identification of Phenolic Compounds from Seed Coats of Differently Colored European Varieties of Pea (Pisum sativum L.) and Characterization of Their Antioxidant and In Vitro Anticancer Activities. Nutr. Cancer 2016, 68, 988–1000. [Google Scholar] [CrossRef] [PubMed]
- Luna-Vital, D.A.; de Mejia, E.G.; Dia, V.P.; Loarca-Pina, G. Peptides in common bean fractions inhibit human colorectal cancer cells. Food Chem. 2014, 157, 347–355. [Google Scholar] [CrossRef] [PubMed]
- Luna-Vital, D.A.; González de Mejía, E.; Loarca-Piña, G. Selective mechanism of action of dietary peptides from common bean on HCT116 human colorectal cancer cells through loss of mitochondrial membrane potential and DNA damage. J. Funct. Foods 2016, 23, 24–39. [Google Scholar] [CrossRef]
- Dan, X.; Ng, T.B.; Wong, J.H.; Chan, Y.S.; Cheung, R.C.F.; Chan, W.Y. A hemagglutinin isolated from Northeast China black beans induced mitochondrial dysfunction and apoptosis in colorectal cancer cells. Biochim. Biophys. Acta (BBA)—Mol. Cell Res. 2016, 1863, 2201–2211. [Google Scholar] [CrossRef] [PubMed]
- Chan, Y.S.; Xia, L.; Ng, T.B. White kidney bean lectin exerts anti-proliferative and apoptotic effects on cancer cells. Int. J. Biol. Macromol. 2016, 85, 335–345. [Google Scholar] [CrossRef] [PubMed]
- Chan, Y.S.; Yu, H.; Xia, L.; Ng, T.B. Lectin from green speckled lentil seeds (Lens culinaris) triggered apoptosis in nasopharyngeal carcinoma cell lines. Chin. Med. 2015, 10, 25. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Zhao, C.; Li, Z.; Zhao, Y.; Shan, S.; Shi, T.; Li, J. Reconstructed mung bean trypsin inhibitor targeting cell surface GRP78 induces apoptosis and inhibits tumor growth in colorectal cancer. Int. J. Biochem. Cell Biol. 2014, 47, 68–75. [Google Scholar] [CrossRef] [PubMed]
- Feregrino-Perez, A.A.; Piñol-Felis, C.; Gomez-Arbones, X.; Guevara-González, R.G.; Campos-Vega, R.; Acosta-Gallegos, J.; Loarca-Piña, G. A Non-digestible Fraction of the Common Bean (Phaseolus vulgaris L.) Induces Cell Cycle Arrest and Apoptosis During Early Carcinogenesis. Plant Foods Hum. Nutr. 2014, 69, 248–254. [Google Scholar] [CrossRef] [PubMed]
- Cruz-Bravo, R.K.; Guevara-González, R.G.; Ramos-Gómez, M.; Oomah, B.D.; Wiersma, P.; Campos-Vega, R.; Loarca-Piña, G. The fermented non-digestible fraction of common bean (Phaseolus vulgaris L.) triggers cell cycle arrest and apoptosis in human colon adenocarcinoma cells. Genes Nutr. 2013, 9, 359. [Google Scholar] [CrossRef] [PubMed]
- Campos-Vega, R.; Guevara-Gonzalez, R.G.; Guevara-Olvera, B.L.; Dave Oomah, B.; Loarca-Piña, G. Bean (Phaseolus vulgaris L.) polysaccharides modulate gene expression in human colon cancer cells (HT-29). Food Res. Int. 2010, 43, 1057–1064. [Google Scholar] [CrossRef]
- Lima, A.I.G.; Mota, J.; Monteiro, S.A.V.S.; Ferreira, R.M.S.B. Legume seeds and colorectal cancer revisited: Protease inhibitors reduce MMP-9 activity and colon cancer cell migration. Food Chem. 2016, 197, 30–38. [Google Scholar] [CrossRef] [PubMed]
- Banerji, A.; Fernandes, A.; Bane, S.; Ahire, S. The field bean protease inhibitor has the potential to suppress B16F10 melanoma cell lung metastasis in mice. Cancer Lett. 1998, 129, 15–20. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
| Sample | Compound | Cell Line/Model | Mechanisms/Outcomes of Anti-Inflammatory Effect | Reference |
|---|---|---|---|---|
| Black, spotted and common bean (Phaseolus spp) | Phenolics | HT-29 human colon cancer cells | Inhibition of pro-inflammatory proteins cyclooxygenase-2 (COX-2), TNFα and NF-κβ, and increased expression of anti-inflammatory protein IL-10. | [32] |
| Lentil (Lens esculenta) Faba bean (Vicia faba) | Phenolics | Faba bean hull extracts exhibited mild LOX inhibitory activity, while lentils inhibited 15-LOX, COX-1 and COX-2. | [14] | |
| Pigmented and non-pigmented beans (dry) (Phaseolus spp) | Phenolics | Female Sprague Dawley rats | Exhibited lowed incidences of mammary cancer, cancer multiplicity and tumour burden. | [35] |
| White kidney bean (Ph. vulgaris) Round purple bean (Phaseolus spp) | Phenolics | RAW 264.7 macrophage cells | Anti-inflammatory effect via a reduction in LPS stimulated macrophages of cytokine mRNA expression and nitric oxide production. | [36] |
| Adzuki bean (Ph. angularis) Mung bean (Vigna radiate) | Phenolics- hull and cotyledon | Protease inhibition was highest in hull faction followed by wholegrain and cotyledon. | [31] | |
| Mung bean (Vigna radiate) | Phenolics- vitexin and isovitexin | RAW 264.7 macrophage cells | Exhibited anti-inflammatory effects. | [8] |
| Black Bean (Phaseolus spp) | Protein hydrolysates | RAW 264.7 macrophage cell | Nitric oxide synthesis was inhibited when treated with protein hydrolysates from germinated beans | [33] |
| Chickpea (Cicer arietinum) | ICR male mice | Reduction in the oxidation of lipid, protein and DNA, downregulation of cyclooxygenase COX-2 and inducible nitric oxide synthase and oncogenic protein β-catenin. | [37] | |
| Mung bean (Vigna radiate) | Gamma-aminobutyric acid (GABA) | Yac-1 and 4T1 lymphoma cells Female Balb/c mice | High anti-cancer cytokine levels, spleen T cell populations, splenocyte cytotoxicity, and spleen/tumour antioxidant levels. Mitotic divisions in spleen were also reduced along with antioxidant and nitric oxide levels. | [34] |
| Sample | Compound | Cell Line/Model | Mechanisms for Apoptosis | Reference |
|---|---|---|---|---|
| Black Bean (Phaseolus spp) | Phenolics—hulls extract and flavonoid fraction | OCI-Ly7 lymphoma cells mouse | Induced cell population to S-phase, increased the overall survival of mice fed with bean extracts by blocking progression to G2/M phase. | [64] |
| Black Bean (Phaseolus spp) | Phenolics | HeLa human cervical cancer cells HaCaT human premalignant keratinocytes | Reduced number of cells in the G0/G1 phase in comparison to control and induced apoptosis. | [60] |
| Black Bean (Phaseolus spp) | Phenolics | HeLa human cervical cancer cells | Methanol extracts induced apoptosis via upregulation of pro-apoptotic proteins, Bax and Caspase-3. | [24] |
| Mung bean sprouts (Vigna radiate) | Phenolics | HeLa human cervical cancer cells, HepG2 human liver cancer cells | Regulation of tumour necrosis factor (TNF-α), Interferon (FN-β, IFNγ), Interleukin (IL-4), apoptotic genes and tumour suppressor genes. | [61] |
| Red beans (dry) (Phaseolus spp) | Phenolics—isoflavones | Female Sprague Dawley rats | Increase in pro-apoptotic proteins BCL-2–associated X protein and reduction in inhibitory apoptotic protein B cell lymphoma 2 and X-linked inhibitor, hence induction of apoptosis was via the mitochondrial pathway. | [63] |
| Peas (Cajanus cajan) | Phenolics | Caco-2 human colon cancer cell | Apoptotic action was linked to the induction of lactate dehydrogenase (LDH) release, DNA fragmentation and upregulation of caspase-3 activity. | [62] |
| Lentil (Lens esculenta) | ||||
| Peas (Cajanus cajan) | Phenolics—epigallocatechin and luteolin | LS174 human colon adenocarcinoma, MDA-MB-453 human breast carcinoma, A594 human lung carcinoma, K562 myelogenous leukaemia | Induction of caspase 3 pathway. | [65] |
| Chickpea (Cicer arietinum) | Phenolics—isoflavones | SKBr3, MCF-7 human breast cancer cells | Upregulation of cytoclasis, apoptotic body formation, caspase 7, caspase 9, P53, and P21 decrease in mitochondrial membrane potential, expression of Bcl-2-associated X protein and increased Bcl-2. | [17] |
| Korean kidney bean husk (Ph. vulgaris) | Phenolics | HT-29 human colon cancer cells | Upregulation of proteins adenosine monophosphate-activated protein kinase (p-AMPK), protein acetyl-CoA carboxylase (p-ACC), p53 and p21. | [10] |
| Speckled lentil (Lens esculenta; Ervum lens) | Peptide-lectin | Nasopharyngeal carcinoma CNE1 and CNE2 cells | Phosphatidylserine externalization, mitochondrial depolarization and cell cycle arrest. An extrinsic apoptotic pathway involving caspase 3, 8, and 9 were also detected in CNE1 cells | [70] |
| Black Bean (Phaseolus spp) | Peptide—hemagglutinin | HCT116, HT-29 human colon cancer cells | Hemagglutinin successfully penetrate the cytoplasm of colorectal cancer cells and instigate mitochondrial dysfunction and apoptotic activity | [68] |
| Mung bean (Vigna radiate) | Peptide—GRP78 binding peptide WIFPWIQL and the active fragment of mung bean trypsin inhibitor | HT-29, SW620, DLD1 human colon cancer cell, FHC human normal colon cells, female mice with severe combined immune deficiency (SCID/NOD) | Activation of multiple apoptotic pathways and induction of G1 phase arrest. | [71] |
| White kidney bean (Ph. vulgaris) | Peptide—WKBL (lectin) | HONE1 epithelial tumour cells | Caspase 3, 8 and 9 were upregulated in cells as a result of WKBL treatment outlining apoptosis as the mechanism of action following an extrinsic pathway. | [69] |
| HepG2 human liver cancer cells | ||||
| MCF-7 human breast cancer cells | ||||
| WRL68 human normal adherent cells | ||||
| Black, spotted and common bean (Phaseolus spp) | Peptides | HCT116, RKO, KM12L4 human colorectal cancer cells | P53 expression was up-regulated along with modification of p21 and cyclin B1 | [66] |
| Large Lima beans (Ph. lunatus) | Peptides—GLTSK, LSGNK, GEGSGA, MTEEY, and MPACGSS | HCT116, CCD-33Co human normal colon cells | GLTSK caused mitochondrial membrane disruption via loss of mitochondrial potential (Δψm) and increased intracellular ROS. GEGSGA caused DNA damage via cleavage of PARP and cell cycle arrest in G1 phase, suggesting oxaliplatin initiated activation and nuclear translocation of p53. | [67] |
| Black Bean (Phaseolus spp) | Short-chain fatty acid | HT-29 human colon cancer cells | Modulation of: RB1, CDC2, CDC25A, NFKB and E2F genes; and pro-apoptotic genes: APAF1, BID, CASP9, FASLG, TNFR10B and BCL2A genes. | [73] |
| Black Bean (Phaseolus spp) | Short-chain fatty acids—butyrate | Male Sprague Dawley rats | Rats demonstrated apoptotic effects and cell cycle arrest in G1 phase. | [72] |
| Black Bean (Phaseolus spp) | Short-chain fatty acids—butyrate | HT-29 human colon cancer cells | DNA fragmentation induced by the extracts indicating that cells were undergoing apoptosis. | [11] |
| Bean (Phaseolus spp) | Polysaccharides—human gut fermented bean | HT-29 human colon cancer cells | Extract induced apoptosis via modulation of 72 p53-mediated signal transduction response genes in human colorectal cancer cells | [74] |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rao, S.; Chinkwo, K.A.; Santhakumar, A.B.; Blanchard, C.L. Inhibitory Effects of Pulse Bioactive Compounds on Cancer Development Pathways. Diseases 2018, 6, 72. https://doi.org/10.3390/diseases6030072
Rao S, Chinkwo KA, Santhakumar AB, Blanchard CL. Inhibitory Effects of Pulse Bioactive Compounds on Cancer Development Pathways. Diseases. 2018; 6(3):72. https://doi.org/10.3390/diseases6030072
Chicago/Turabian StyleRao, Shiwangni, Kenneth A. Chinkwo, Abishek B. Santhakumar, and Christopher L. Blanchard. 2018. "Inhibitory Effects of Pulse Bioactive Compounds on Cancer Development Pathways" Diseases 6, no. 3: 72. https://doi.org/10.3390/diseases6030072
APA StyleRao, S., Chinkwo, K. A., Santhakumar, A. B., & Blanchard, C. L. (2018). Inhibitory Effects of Pulse Bioactive Compounds on Cancer Development Pathways. Diseases, 6(3), 72. https://doi.org/10.3390/diseases6030072