
Cleft Candidate Genes and Their Products in Human Unilateral Cleft Lip Tissue




Abstract
:1. Introduction
2. Materials and Methods
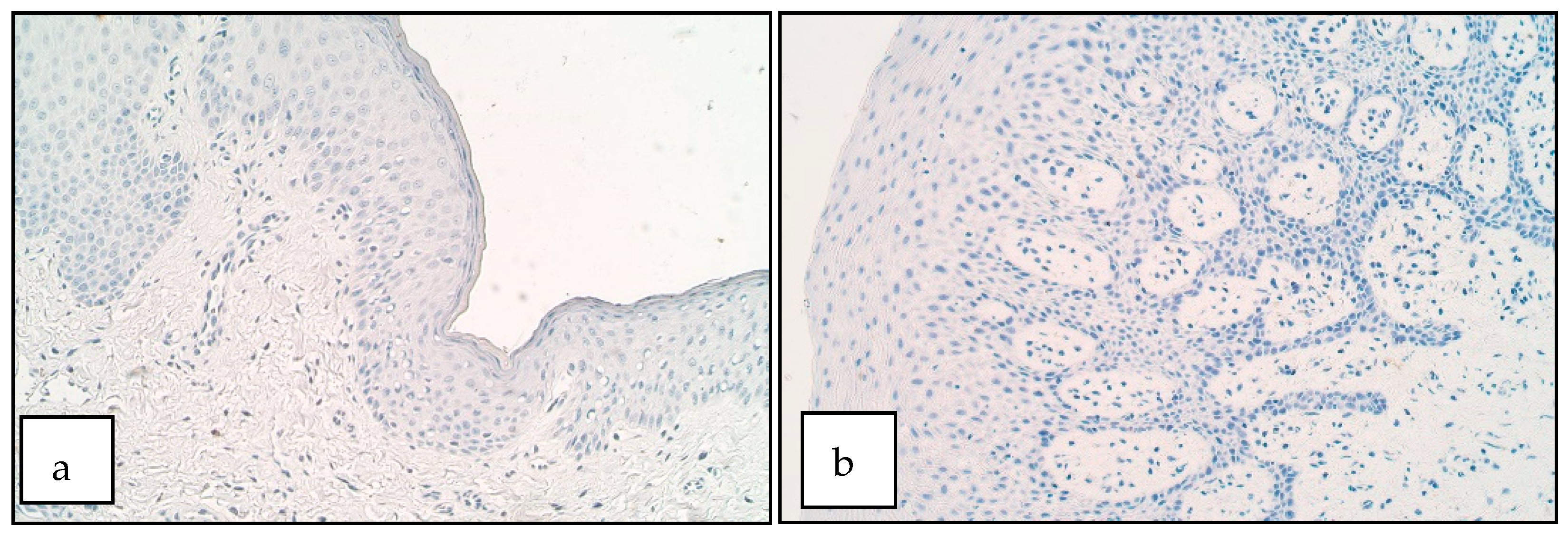
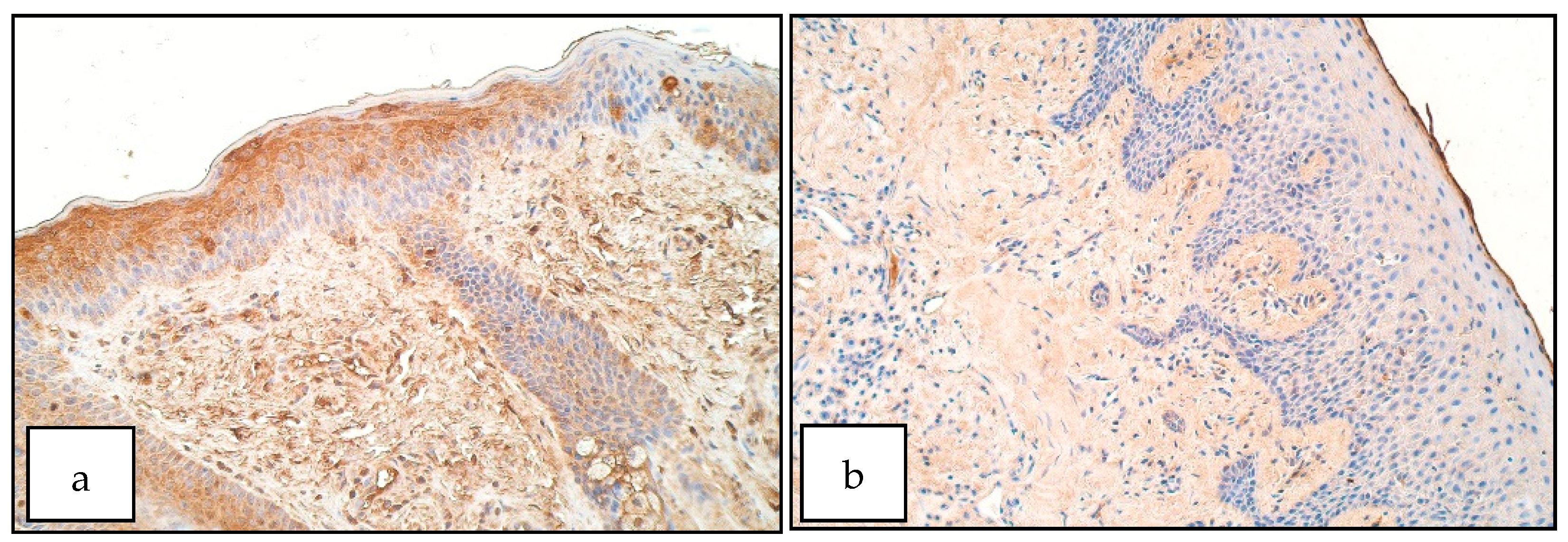
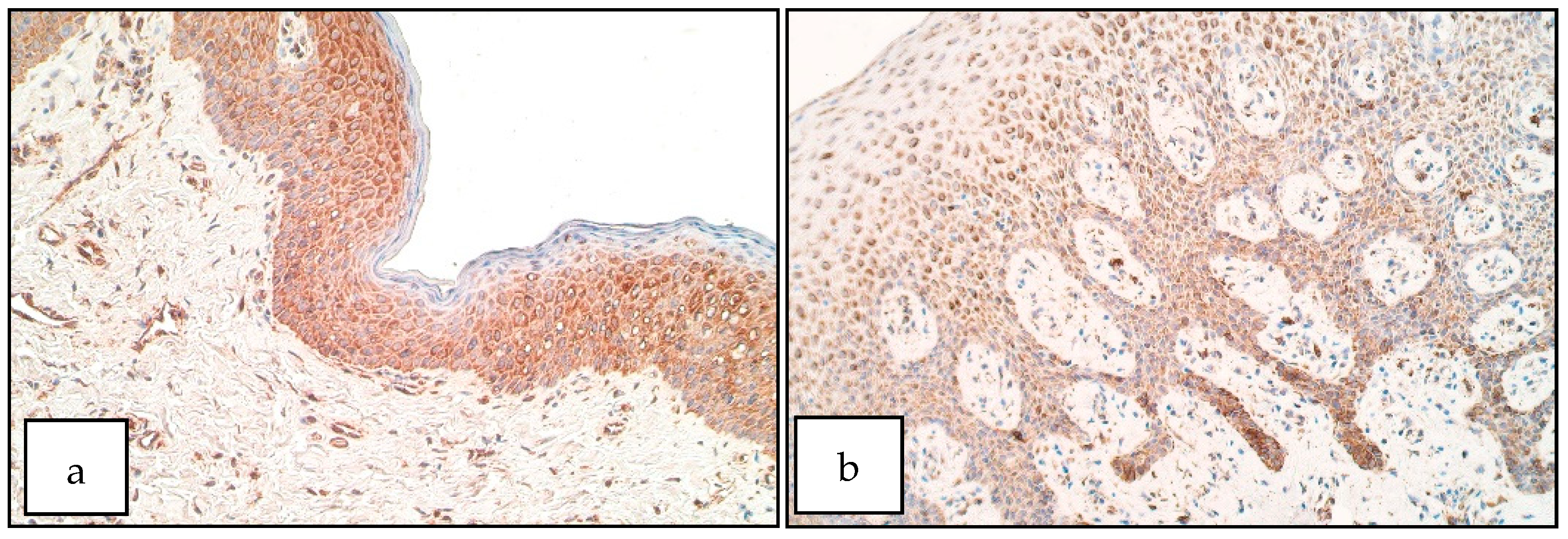
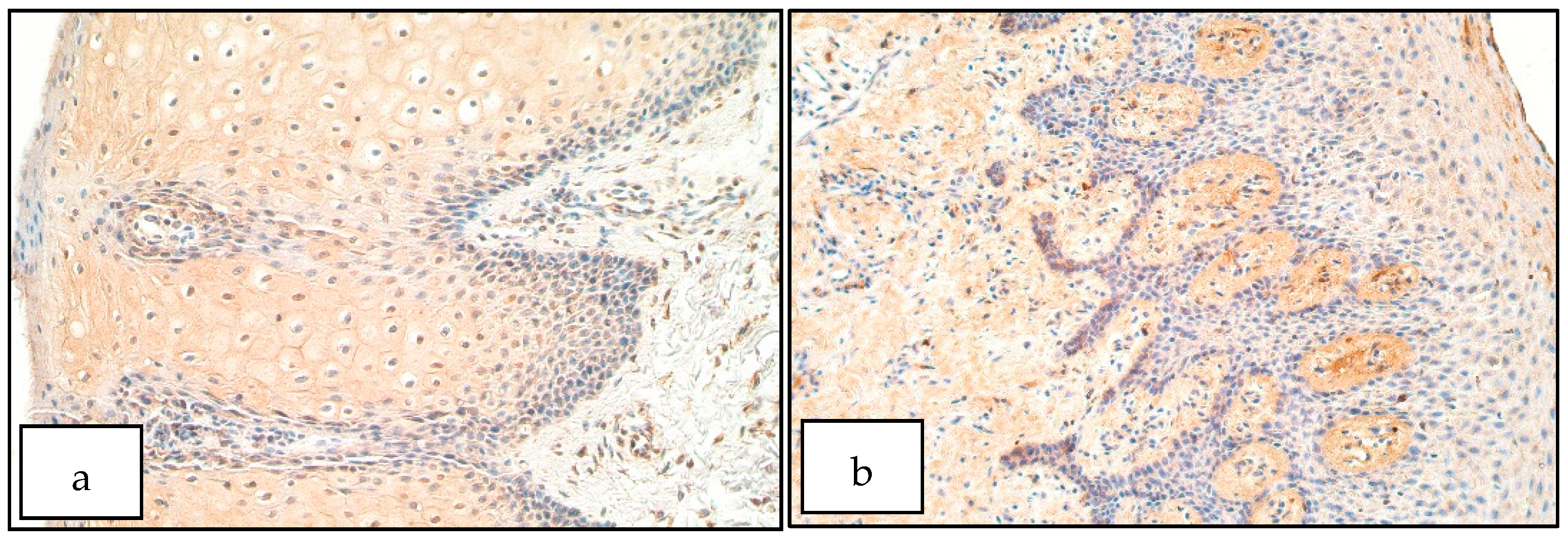
3. Results
4. Discussion
5. Conclusions
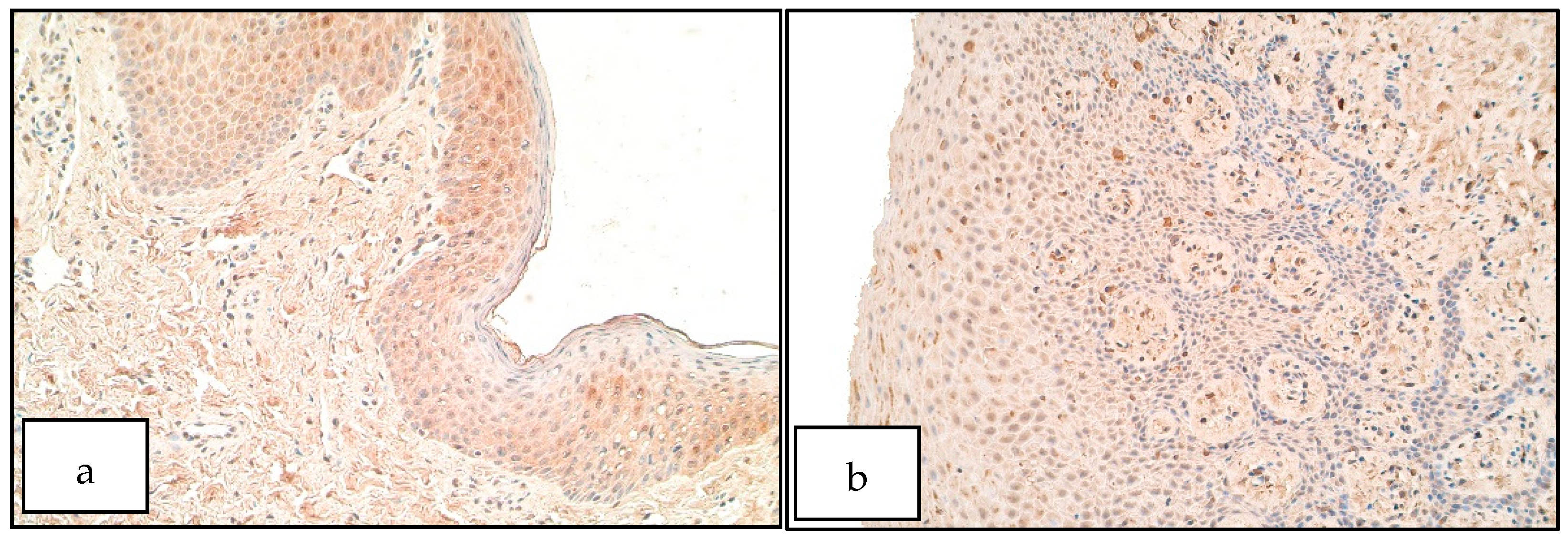
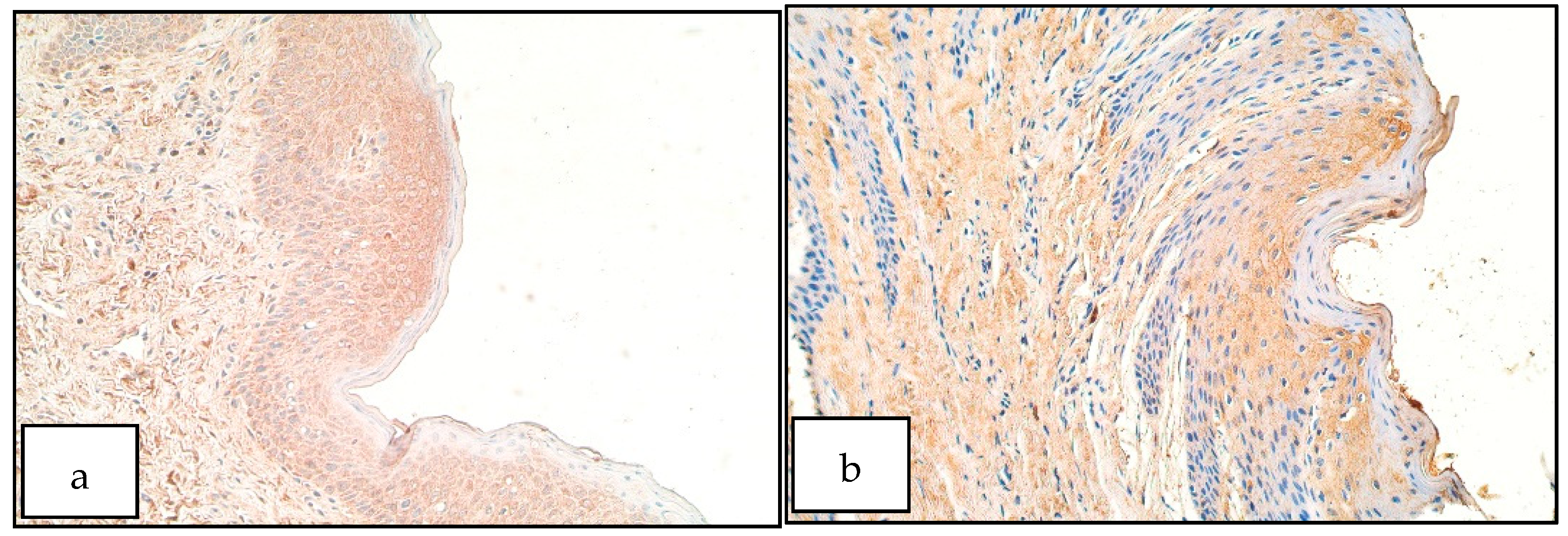
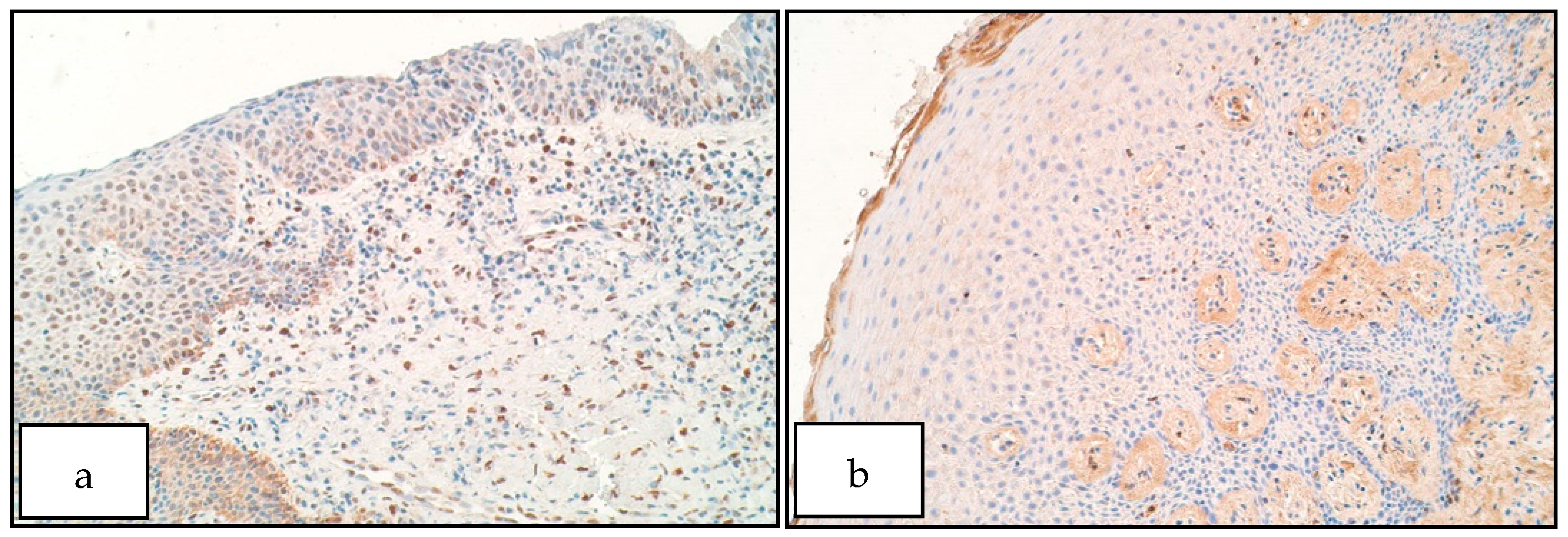
- Increase of SHH positive structures in the unilateral cleft lip affected connective tissue could indicate a disruption of correct tissue formation in cleft-affected tissue, which could have affected proper lip fusion and the development of cleft lip.
- Increase of SOX3 positive structures within patient epithelium could indicate a possible involvement of SOX3 in the formation of unilateral cleft lip possibly by affecting cell proliferation within the cleft-affected tissue.
- Increase of FOXE1 positive structures within the cleft-affected connective tissue could indicate improper cell proliferation and formation within the developing lip region, resulting in remodeling processes within cleft-affected tissues.
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Shkoukani, M.A.; Chen, M.; Vong, A. Cleft Lip—A Comprehensive Review. Front. Pediatr. 2013, 1, 53. [Google Scholar] [CrossRef] [PubMed]
- Saleem, K.; Zaib, T.; Sun, W.; Fu, S. Assessment of Candidate Genes and Genetic Heterogeneity in Human Non Syndromic Orofacial Clefts Specifically Non Syndromic Cleft Lip with or without Palate. Heliyon 2019, 5, e03019. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Merlo, G.R.; Zerega, B.; Paleari, L.; Trombino, S.; Mantero, S.; Levi, G. Multiple Functions of Dlx Genes. Int. J. Dev. Biol. 2000, 44, 619–626. [Google Scholar]
- Wu, D.; Mandal, S.; Choi, A.; Anderson, A.; Prochazkova, M.; Perry, H.; Gil-Da-Silva-Lopes, V.L.; Lao, R.; Wan, E.; Tang, P.L.-F.; et al. DLX4 Is Associated with Orofacial Clefting and Abnormal Jaw Development. Hum. Mol. Genet. 2015, 24, 4340–4352. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, M.; Bian, Z. Association between DLX4 Polymorphisms and Nonsyndromic Orofacial Clefts in a Chinese Han Population. Cleft Palate Craniofac. J. 2019, 56, 357–362. [Google Scholar] [CrossRef] [PubMed]
- Vieira, A.R.; Avila, J.R.; Daack-Hirsch, S.; Dragan, E.; Félix, T.M.; Rahimov, F.; Harrington, J.; Schultz, R.R.; Watanabe, Y.; Johnson, M.; et al. Medical Sequencing of Candidate Genes for Nonsyndromic Cleft Lip and Palate. PLoS Genet. 2005, 1, e64. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alappat, S.; Zhang, Z.Y.; Chen, Y.P. Msx Homeobox Gene Family and Craniofacial Development. Cell Res. 2003, 13, 429–442. [Google Scholar] [CrossRef]
- Molla, M.; Descroix, V.; Aïoub, M.; Simon, S.; Castañeda, B.; Hotton, D.; Bolaños, A.; Simon, Y.; Lezot, F.; Goubin, G.; et al. Enamel Protein Regulation and Dental and Periodontal Physiopathology in MSX2 Mutant Mice. Am. J. Pathol. 2010, 177, 2516–2526. [Google Scholar] [CrossRef]
- de Pelegrino, K.O.; Sugayama, S.; Lezirovitz, K.; Catelani, A.L.; Kok, F.; de Chauffaille, M.L. MSX2 Copy Number Increase and Craniosynostosis: Copy Number Variation Detected by Array Comparative Genomic Hybridization. Clinics 2012, 67, 981–985. [Google Scholar] [CrossRef]
- Müller, U. Discovery of MSX2 Mutation in Craniosynostosis: A Retrospective View. Monogr. Hum. Genet. 2011, 19, 8–12. [Google Scholar] [CrossRef]
- Satokata, I.; Ma, L.; Ohshima, H.; Bei, M.; Woo, I.; Nishizawa, K.; Maeda, T.; Takano, Y.; Uchiyama, M.; Heaney, S.; et al. Msx2 Deficiency in Mice Causes Pleiotropic Defects in Bone Growth and Ectodermal Organ Formation. Nat. Genet. 2000, 24, 391–395. [Google Scholar] [CrossRef] [PubMed]
- Tomotsune, D.; Shirai, M.; Takihara, Y.; Shimada, K. Regulation of Hoxb3 Expression in the Hindbrain and Pharyngeal Arches by Rae28, a Member of the Mammalian Polycomb Group of Genes. Mech. Dev. 2000, 98, 165–169. [Google Scholar] [CrossRef]
- Frisdal, A.; Trainor, P.A. Development and Evolution of the Pharyngeal Apparatus: Development and Evolution of the Pharyngeal Apparatus. Wiley Interdiscip. Rev. Dev. Biol. 2014, 3, 403–418. [Google Scholar] [CrossRef] [Green Version]
- Kurosaka, H.; Iulianella, A.; Williams, T.; Trainor, P.A. Disrupting Hedgehog and WNT Signaling Interactions Promotes Cleft Lip Pathogenesis. J. Clin. Investig. 2014, 124, 1660–1671. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Everson, J.L.; Fink, D.M.; Yoon, J.W.; Leslie, E.J.; Kietzman, H.W.; Ansen-Wilson, L.J.; Chung, H.M.; Walterhouse, D.O.; Marazita, M.L.; Lipinski, R.J. Sonic Hedgehog Regulation of Foxf2 Promotes Cranial Neural Crest Mesenchyme Proliferation and Is Disrupted in Cleft Lip Morphogenesis. Development 2017, 144, 2082–2091. [Google Scholar] [CrossRef] [Green Version]
- Hammond, N.L.; Brookes, K.J.; Dixon, M.J. Ectopic Hedgehog Signaling Causes Cleft Palate and Defective Osteogenesis. J. Dent. Res. 2018, 97, 1485–1493. [Google Scholar] [CrossRef]
- Menezes, R.; Letra, A.; Kim, A.H.; Küchler, E.C.; Day, A.; Tannure, P.N.; Gomes da Motta, L.; Paiva, K.B.S.; Granjeiro, J.M.; Vieira, A.R. Studies with Wnt Genes and Nonsyndromic Cleft Lip and Palate. Birth Defects Res. A Clin. Mol. Teratol. 2010, 88, 995–1000. [Google Scholar] [CrossRef] [Green Version]
- Chiquet, B.T.; Blanton, S.H.; Burt, A.; Ma, D.; Stal, S.; Mulliken, J.B.; Hecht, J.T. Variation in WNT Genes Is Associated with Non-Syndromic Cleft Lip with or without Cleft Palate. Hum. Mol. Genet. 2008, 17, 2212–2218. [Google Scholar] [CrossRef] [Green Version]
- Mani, P.; Jarrell, A.; Myers, J.; Atit, R. Visualizing Canonical Wnt Signaling during Mouse Craniofacial Development. Dev. Dyn. 2010, 239, 354–363. [Google Scholar] [CrossRef] [Green Version]
- Leslie, E.J.; Carlson, J.C.; Shaffer, J.R.; Butali, A.; Buxó, C.J.; Castilla, E.E.; Christensen, K.; Deleyiannis, F.W.B.; Leigh Field, L.; Hecht, J.T.; et al. Genome-Wide Meta-Analyses of Nonsyndromic Orofacial Clefts Identify Novel Associations between FOXE1 and All Orofacial Clefts, and TP63 and Cleft Lip with or without Cleft Palate. Hum. Genet. 2017, 136, 275–286. [Google Scholar] [CrossRef] [Green Version]
- Moreno, L.M.; Mansilla, M.A.; Bullard, S.A.; Cooper, M.E.; Busch, T.D.; Machida, J.; Johnson, M.K.; Brauer, D.; Krahn, K.; Daack-Hirsch, S.; et al. FOXE1 Association with Both Isolated Cleft Lip with or without Cleft Palate, and Isolated Cleft Palate. Hum. Mol. Genet. 2009, 18, 4879–4896. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moreno Uribe, L.M.; Fomina, T.; Munger, R.G.; Romitti, P.A.; Jenkins, M.M.; Gjessing, H.K.; Gjerdevik, M.; Christensen, K.; Wilcox, A.J.; Murray, J.C.; et al. A Population-Based Study of Effects of Genetic Loci on Orofacial Clefts. J. Dent. Res. 2017, 96, 1322–1329. [Google Scholar] [CrossRef] [Green Version]
- Murdoch, B.; DelConte, C.; García-Castro, M.I. Pax7 Lineage Contributions to the Mammalian Neural Crest. PLoS ONE 2012, 7, e41089. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Butali, A.; Suzuki, S.; Cooper, M.E.; Mansilla, A.M.; Cuenco, K.; Leslie, E.J.; Suzuki, Y.; Niimi, T.; Yamamoto, M.; Ayanga, G.; et al. Replication of Genome Wide Association Identified Candidate Genes Confirm the Role of Common and Rare Variants in PAX7 and VAX1 in the Etiology of Nonsyndromic CL(P). Am. J. Med. Genet. A 2013, 161, 965–972. [Google Scholar] [CrossRef] [Green Version]
- Sull, J.W.; Liang, K.-Y.; Hetmanski, J.B.; Fallin, M.D.; Ingersoll, R.G.; Park, J.; Wu-Chou, Y.-H.; Chen, P.K.; Chong, S.S.; Cheah, F.; et al. Maternal Transmission Effects of the PAX Genes among Cleft Case-Parent Trios from Four Populations. Eur. J. Hum. Genet. 2009, 17, 831–839. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zalc, A.; Rattenbach, R.; Auradé, F.; Cadot, B.; Relaix, F. Pax3 and Pax7 Play Essential Safeguard Functions against Environmental Stress-Induced Birth Defects. Dev. Cell 2015, 33, 56–66. [Google Scholar] [CrossRef] [Green Version]
- Rizzoti, K.; Lovell-Badge, R. SOX3 Activity during Pharyngeal Segmentation Is Required for Craniofacial Morphogenesis. Development 2007, 134, 3437–3448. [Google Scholar] [CrossRef] [Green Version]
- Watanabe, M.; Kawasaki, K.; Kawasaki, M.; Portaveetus, T.; Oommen, S.; Blackburn, J.; Nagai, T.; Kitamura, A.; Nishikawa, A.; Kodama, Y.; et al. Spatio-Temporal Expression of Sox Genes in Murine Palatogenesis. Gene Expr. Patterns 2016, 21, 111–118. [Google Scholar] [CrossRef] [Green Version]
- Kawasaki, K.; Kawasaki, M.; Watanabe, M.; Idrus, E.; Nagai, T.; Oommen, S.; Maeda, T.; Hagiwara, N.; Que, J.; Sharpe, P.T.; et al. Expression of Sox Genes in Tooth Development. Int. J. Dev. Biol. 2015, 59, 471–478. [Google Scholar] [CrossRef]
- Hsu, S.M.; Raine, L.; Fanger, H. The Use of Antiavidin Antibody and Avidin-Biotin-Peroxidase Complex in Immunoperoxidase Technics. Am. J. Clin. Pathol. 1981, 75, 816–821. [Google Scholar] [CrossRef] [PubMed]
- Jankovska, I.; Pilmane, M.; Akota, I. Expression of Gene Proteins, Interleukins and β-Defensin in Cleft-Affected Tissue. Stomatologija 2017, 19, 103–108. [Google Scholar]
- Kurosaka, H. The Roles of Hedgehog Signaling in Upper Lip Formation. BioMed Res. Int. 2015, 2015, 901041. [Google Scholar] [CrossRef] [Green Version]
- Sun, M.R.; Chung, H.M.; Matsuk, V.; Fink, D.M.; Stebbins, M.J.; Palecek, S.P.; Shusta, E.V.; Lipinski, R.J. Sonic Hedgehog Signaling in Cranial Neural Crest Cells Regulates Microvascular Morphogenesis in Facial Development. Front. Cell Dev. Biol. 2020, 8, 590539. [Google Scholar] [CrossRef] [PubMed]
- Xavier, G.M.; Seppala, M.; Barrell, W.; Birjandi, A.A.; Geoghegan, F.; Cobourne, M.T. Hedgehog Receptor Function during Craniofacial Development. Dev. Biol. 2016, 415, 198–215. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schock, E.N.; LaBonne, C. Sorting Sox: Diverse Roles for Sox Transcription Factors during Neural Crest and Craniofacial Development. Front. Physiol. 2020, 11, 606889. [Google Scholar] [CrossRef]
- Kamachi, Y.; Kondoh, H. Sox Proteins: Regulators of Cell Fate Specification and Differentiation. Development 2013, 140, 4129–4144. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bhatlekar, S.; Fields, J.Z.; Boman, B.M. Role of HOX Genes in Stem Cell Differentiation and Cancer. Stem Cells Int. 2018, 2018, 1–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paço, A.; Aparecida de Bessa Garcia, S.; Leitão Castro, J.; Costa-Pinto, A.R.; Freitas, R. Roles of the HOX Proteins in Cancer Invasion and Metastasis. Cancers 2020, 13, 10. [Google Scholar] [CrossRef] [PubMed]
- Letra, A.; Menezes, R.; Govil, M.; Fonseca, R.F.; McHenry, T.; Granjeiro, J.M.; Castilla, E.E.; Orioli, I.M.; Marazita, M.L.; Vieira, A.R. Follow-up Association Studies of Chromosome Region 9q and Nonsyndromic Cleft Lip/Palate. Am. J. Med. Genet. A 2010, 152, 1701–1710. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carlsson, P.; Mahlapuu, M. Forkhead Transcription Factors: Key Players in Development and Metabolism. Dev. Biol. 2002, 250, 1–23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Venza, I.; Visalli, M.; Parrillo, L.; De Felice, M.; Teti, D.; Venza, M. MSX1 and TGF-Beta3 Are Novel Target Genes Functionally Regulated by FOXE1. Hum. Mol. Genet. 2011, 20, 1016–1025. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Benayoun, B.A.; Caburet, S.; Veitia, R.A. Forkhead Transcription Factors: Key Players in Health and Disease. Trends Genet. 2011, 27, 224–232. [Google Scholar] [CrossRef] [PubMed]
- Biggs, L.C.; Goudy, S.L.; Dunnwald, M. Palatogenesis and Cutaneous Repair: A Two-Headed Coin: Palatogenesis and cutaneous repair. Dev. Dyn. 2015, 244, 289–310. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jeng, K.-S.; Chang, C.-F.; Lin, S.-S. Sonic Hedgehog Signaling in Organogenesis, Tumors, and Tumor Microenvironments. Int. J. Mol. Sci. 2020, 21, 758. [Google Scholar] [CrossRef] [Green Version]
- Bychkov, A.; Saenko, V.; Nakashima, M.; Mitsutake, N.; Rogounovitch, T.; Nikitski, A.; Orim, F.; Yamashita, S. Patterns of FOXE1 Expression in Papillary Thyroid Carcinoma by Immunohistochemistry. Thyroid 2013, 23, 817–828. [Google Scholar] [CrossRef] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Designations Used | Explanations |
|---|---|
| 0 | No positive structures in the visual field |
| 0/+ | Rare occurrence of positive structures in the visual field |
| + | Few positive structures in the visual field |
| +/++ | Few to moderate number of positive structures in the visual field |
| ++ | Moderate number of positive structures in the visual field |
| ++/+++ | Moderate to numerous positive structures in the visual field |
| +++ | Numerous positive structures in the visual field |
| +++/++++ | Numerous to abundant positive structures in the visual field |
| ++++ | Abundance of positive structures in the visual field |
| DLX4 | MSX2 | HOXB3 | SHH | PAX7 | SOX3 | WNT3A | FOXE1 | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| E | CT | E | CT | E | CT | E | CT | E | CT | E | CT | E | CT | E | CT | |
| Patients | ++ | ++ | 0 | 0-0/+ | ++/+++ | ++ | ++/+++ | ++/+++ | ++ | ++ | ++/+++ | ++ | +/++ | + | ++ | ++ |
| Controls | ++ | +/++ | 0 | 0 | + | ++ | + | +/++ | +/++ | +/++ | +/++ | +/++ | +/++ | + | +/++ | + |
| U | 20.0 | 17.5 | 25.0 | 22.5 | 11.0 | 21.0 | 10.0 | 6.5 | 10.0 | 14.5 | 7.0 | 15.0 | 21.0 | 23.0 | 14.0 | 5.0 |
| p | 0.594 | 0.371 | 1.000 | 0.768 | 0.099 | 0.679 | 0.075 | 0.019 | 0.075 | 0.206 | 0.028 | 0.254 | 0.679 | 0.859 | 0.206 | 0.013 |
| DLX4 | MSX2 | HOXB3 | SHH | PAX7 | SOX3 | WNT3A | FOXE1 | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| E | CT | E | CT | E | CT | E | CT | E | CT | E | CT | E | CT | E | CT | |
| P1 | ++ | ++ | 0 | 0/0/+ | +++ | +++ | ++ | ++ | ++ | ++/+++ | +++ | ++ | +/++ | + | +/++ | ++ |
| P2 | 0 | 0/+ | 0 | 0 | 0 | + | 0 | ++ | 0 | 0/+ | 0 | ++ | 0/+ | 0/+ | 0 | 0 |
| P3 | ++ | ++/+++ | 0 | 0/0/+ | +++ | +++/++++ | ++ | ++/+++ | ++/+++ | +++ | +++ | ++ | +++ | ++ | + | ++ |
| P4 | ++ | +/++ | 0 | 0 | ++/+++ | ++ | ++/+++ | +++ | ++/+++ | +++ | ++/+++ | ++ | + | + | ++ | ++ |
| P5 | ++ | ++ | 0 | 0/0/+ | ++/+++ | ++/+++ | +++ | +/++ | +++ | ++ | ++/+++ | ++/+++ | 0/+ | 0/+ | ++/+++ | ++ |
| P6 | ++ | +/++ | 0 | 0 | ++ | + | +++/++++ | ++/+++ | ++ | ++ | ++/+++ | ++ | +/++ | ++ | ++/+++ | ++ |
| P7 | ++/+++ | ++/+++ | 0 | 0 | +/++ | +/++ | +++ | +++ | ++ | ++/+++ | ++ | +/++ | ++ | +/++ | ++/+++ | ++/+++ |
| P8 | ++ | ++ | 0 | 0/0/+ | ++ | ++/+++ | ++/+++ | +++ | ++ | +/++ | ++/+++ | ++ | ++ | + | ++ | ++/+++ |
| P9 | +/++ | ++ | 0 | 0 | +/++ | ++ | ++/+++ | ++/+++ | ++ | ++ | ++/+++ | +/++ | + | + | ++/+++ | +++ |
| P10 | +++ | ++/+++ | 0 | 0/0/+ | ++/+++ | +/++ | ++/+++ | ++ | ++/+++ | ++ | ++ | +/++ | ++/+++ | ++ | ++ | ++ |
| Median (P) | ++ | ++ | 0 | 0/0/+ | ++/+++ | ++ | ++/+++ | ++/+++ | ++ | ++ | ++/+++ | ++ | +/++ | + | ++ | ++ |
| C1 | ++ | + | 0 | 0 | + | ++ | + | +/++ | ++ | +/++ | ++ | ++ | ++ | + | ++ | + |
| C2 | ++/+++ | ++/+++ | 0 | 0/0/+ | ++ | +/++ | ++/+++ | ++ | +/++ | ++/+++ | ++ | +/++ | +/++ | +/++ | +/++ | +/++ |
| C3 | 0 | +/++ | 0 | 0 | + | ++ | + | +/++ | + | + | + | + | +/++ | + | 0/+ | 0/+ |
| C4 | + | + | 0 | 0 | + | ++ | ++ | ++ | + | + | +/++ | ++ | + | + | ++ | + |
| C5 | ++ | ++ | 0 | 0/0/+ | ++ | +/++ | + | +/++ | ++ | ++ | +/++ | +/++ | 0/+ | ++ | + | + |
| Median (C) | ++ | +/++ | 0 | 0 | + | ++ | + | +/++ | +/++ | +/++ | +/++ | +/++ | +/++ | + | +/++ | + |
| Strength of Correlation | Correlations between Immunopositive Structures in Patient Group Epithelium | rs | p-Value |
|---|---|---|---|
| Very strong: 0.8–1.0 | SHH and FOXE1 | 0.918 | <0.001 |
| Strong: 0.6–0.79 | HOXB3 and SOX3 | 0.742 | 0.014 |
| HOXB3 and PAX7 | 0.677 | 0.032 | |
| DLX4 and WNT3A | 0.660 | 0.038 |
| Strength of Correlation | Correlations between Immunopositive Structures in Patient Group Epithelium | rs | p-Value |
|---|---|---|---|
| Very strong: 0.8–1.0 | WNT3A in epithelium and WNT3A in connective tissue | 0.837 | 0.003 |
| SOX3 in epithelium and HOXB3 in connective tissue | 0.817 | 0.004 | |
| Strong: 0.6–0.79 | HOXB3 in epithelium and HOXB3 in connective tissue | 0.727 | 0.017 |
| DLX4 in epithelium and DLX4 in connective tissue | 0.691 | 0.027 | |
| PAX7 in epithelium and HOXB3 in connective tissue | 0.677 | 0.032 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vaivads, M.; Akota, I.; Pilmane, M. Cleft Candidate Genes and Their Products in Human Unilateral Cleft Lip Tissue. Diseases 2021, 9, 26. https://doi.org/10.3390/diseases9020026
Vaivads M, Akota I, Pilmane M. Cleft Candidate Genes and Their Products in Human Unilateral Cleft Lip Tissue. Diseases. 2021; 9(2):26. https://doi.org/10.3390/diseases9020026
Chicago/Turabian StyleVaivads, Mārtiņš, Ilze Akota, and Māra Pilmane. 2021. "Cleft Candidate Genes and Their Products in Human Unilateral Cleft Lip Tissue" Diseases 9, no. 2: 26. https://doi.org/10.3390/diseases9020026
APA StyleVaivads, M., Akota, I., & Pilmane, M. (2021). Cleft Candidate Genes and Their Products in Human Unilateral Cleft Lip Tissue. Diseases, 9(2), 26. https://doi.org/10.3390/diseases9020026