
Cinnamomum bejolghota Extract Inhibits Colorectal Cancer Cell Metastasis and TGF-β1-Induced Epithelial-Mesenchymal Transition via Smad and Non-Smad Signaling Pathway



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
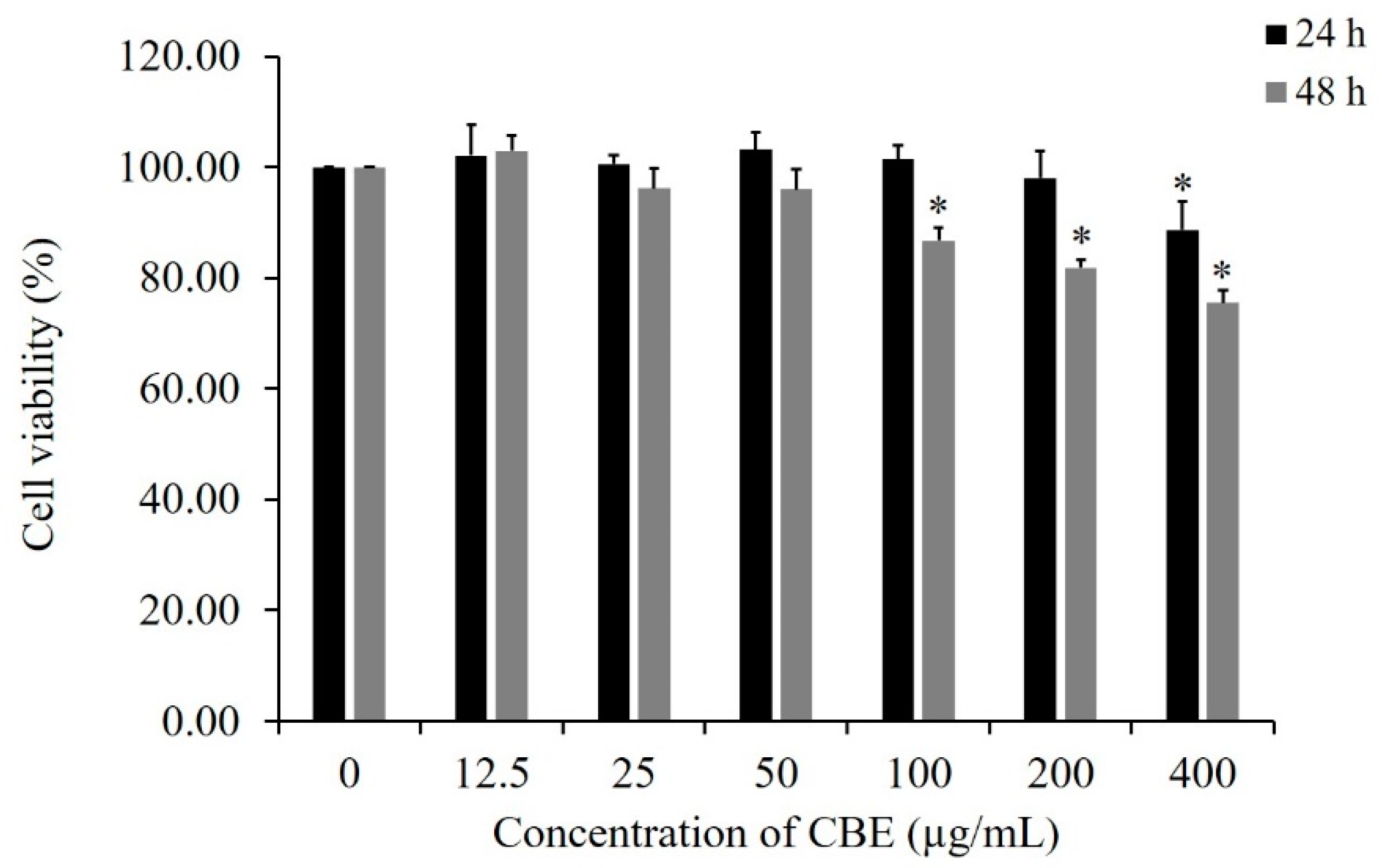
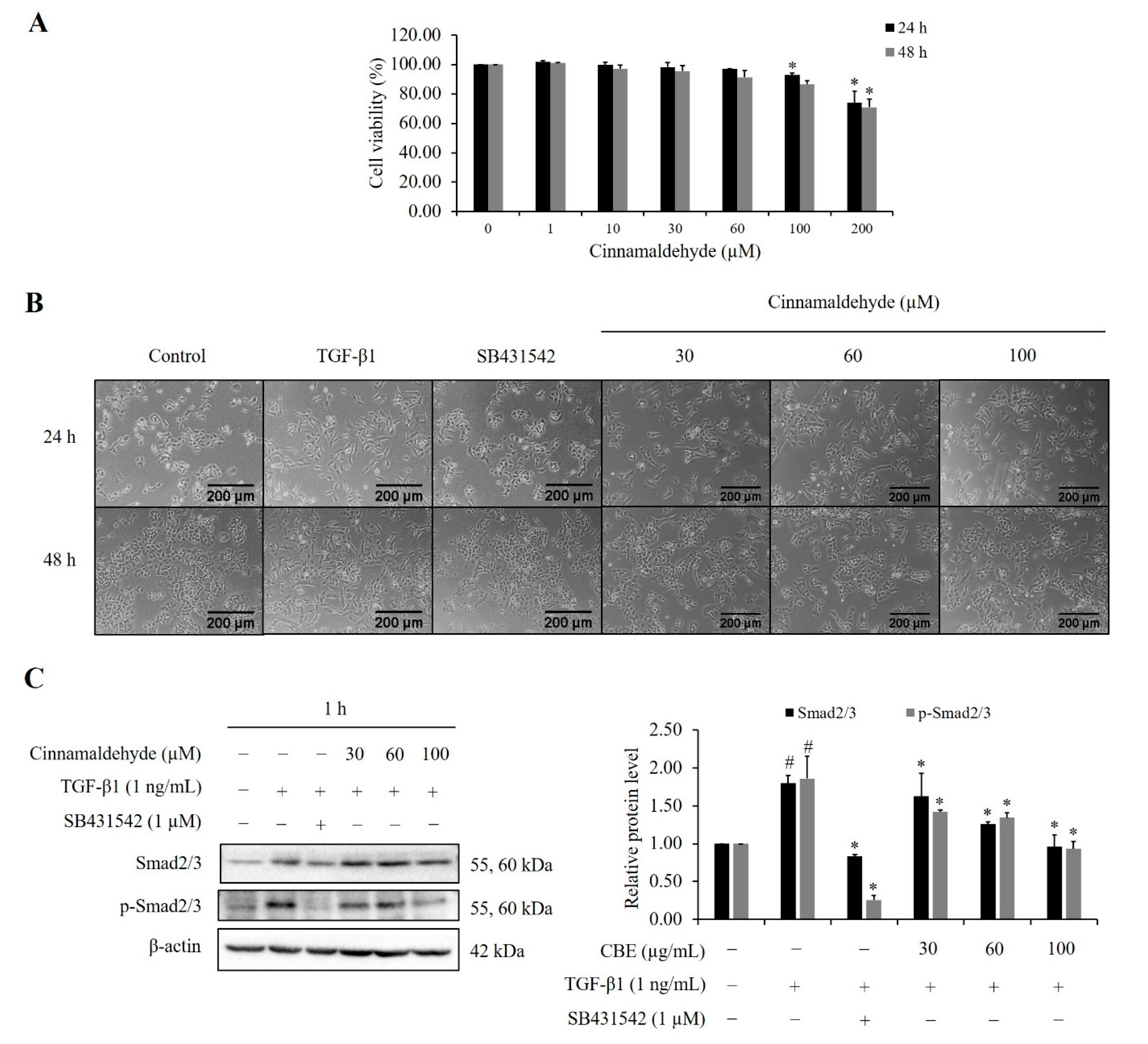
2.1. Cytotoxic Effect of CBE on the Growth of LoVo Cells
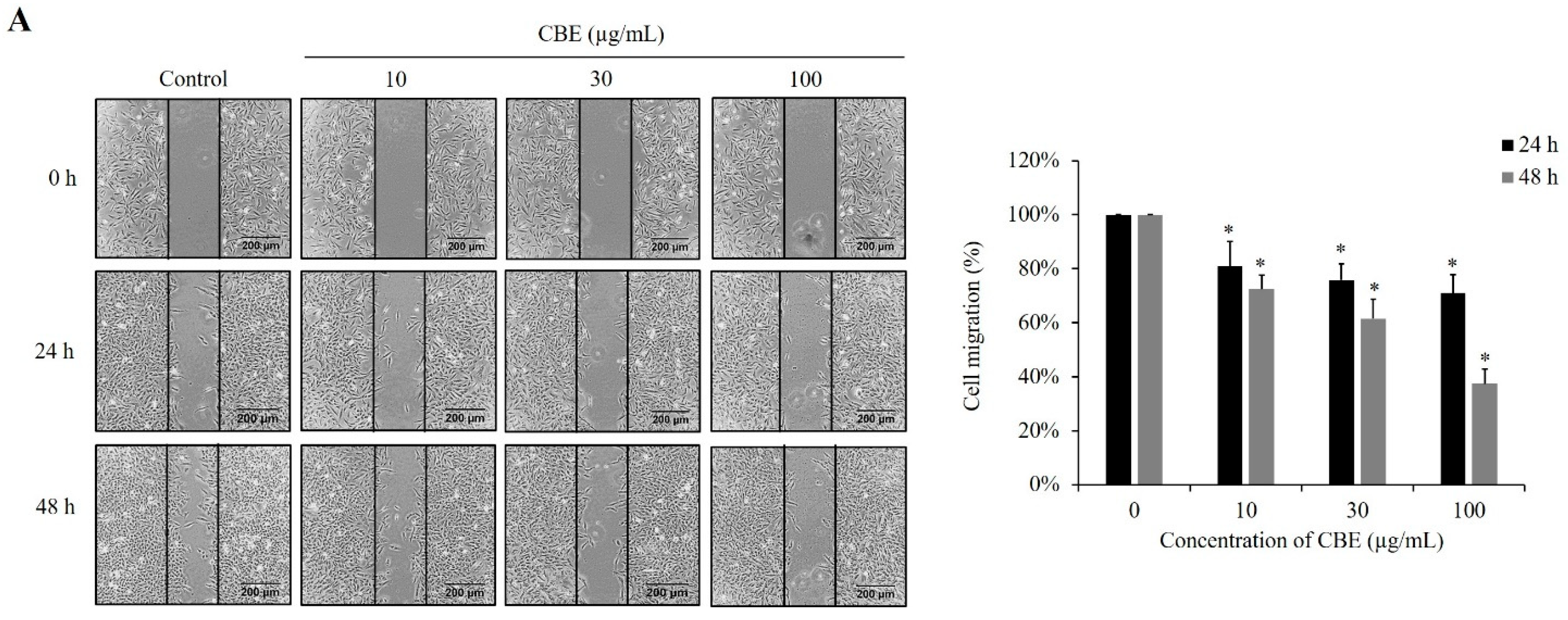
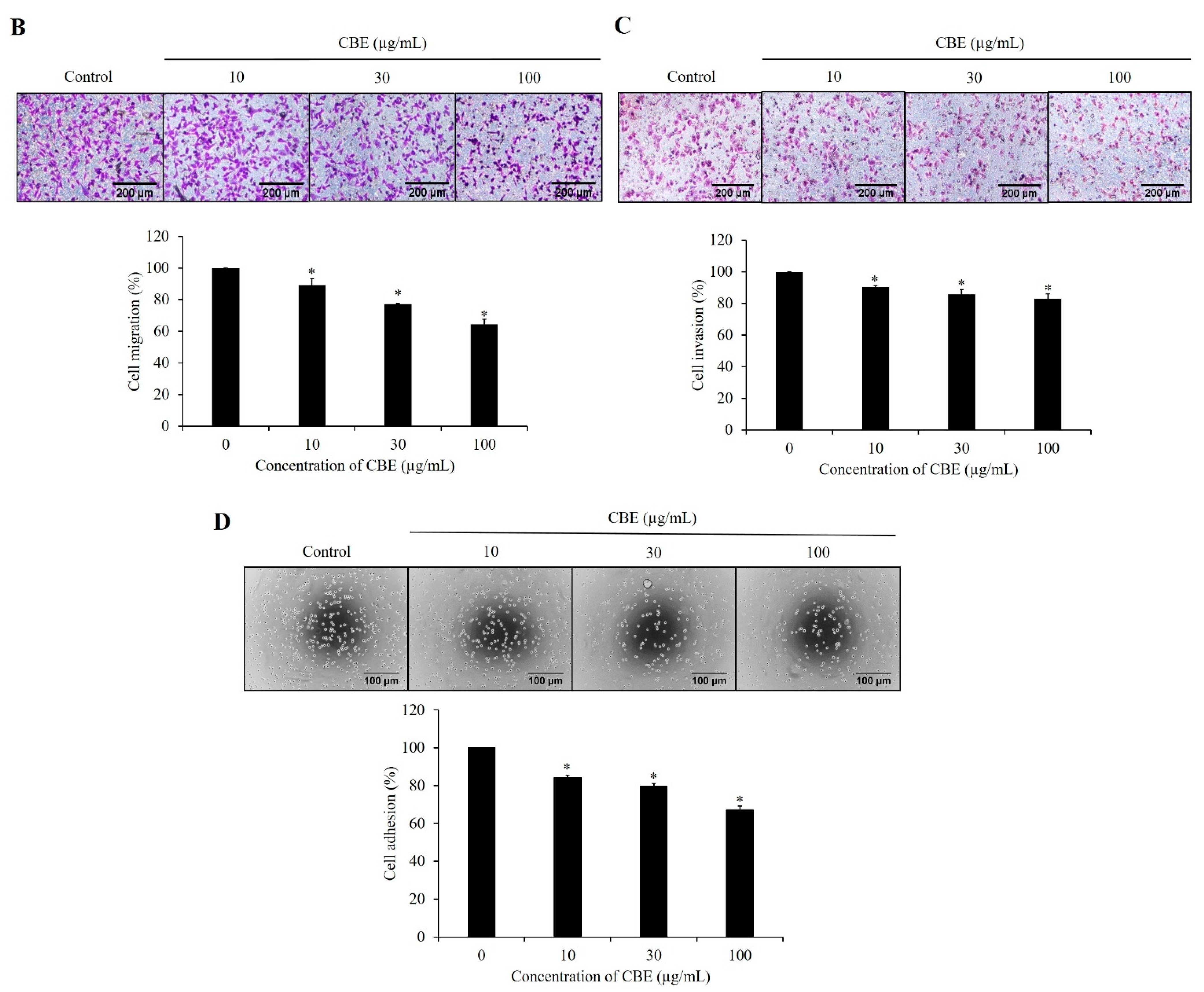
2.2. CBE Reduces Migration, Invasion, and Adhesion
2.3. Effect of CBE on TGF-β1-Induced Morphological Changes
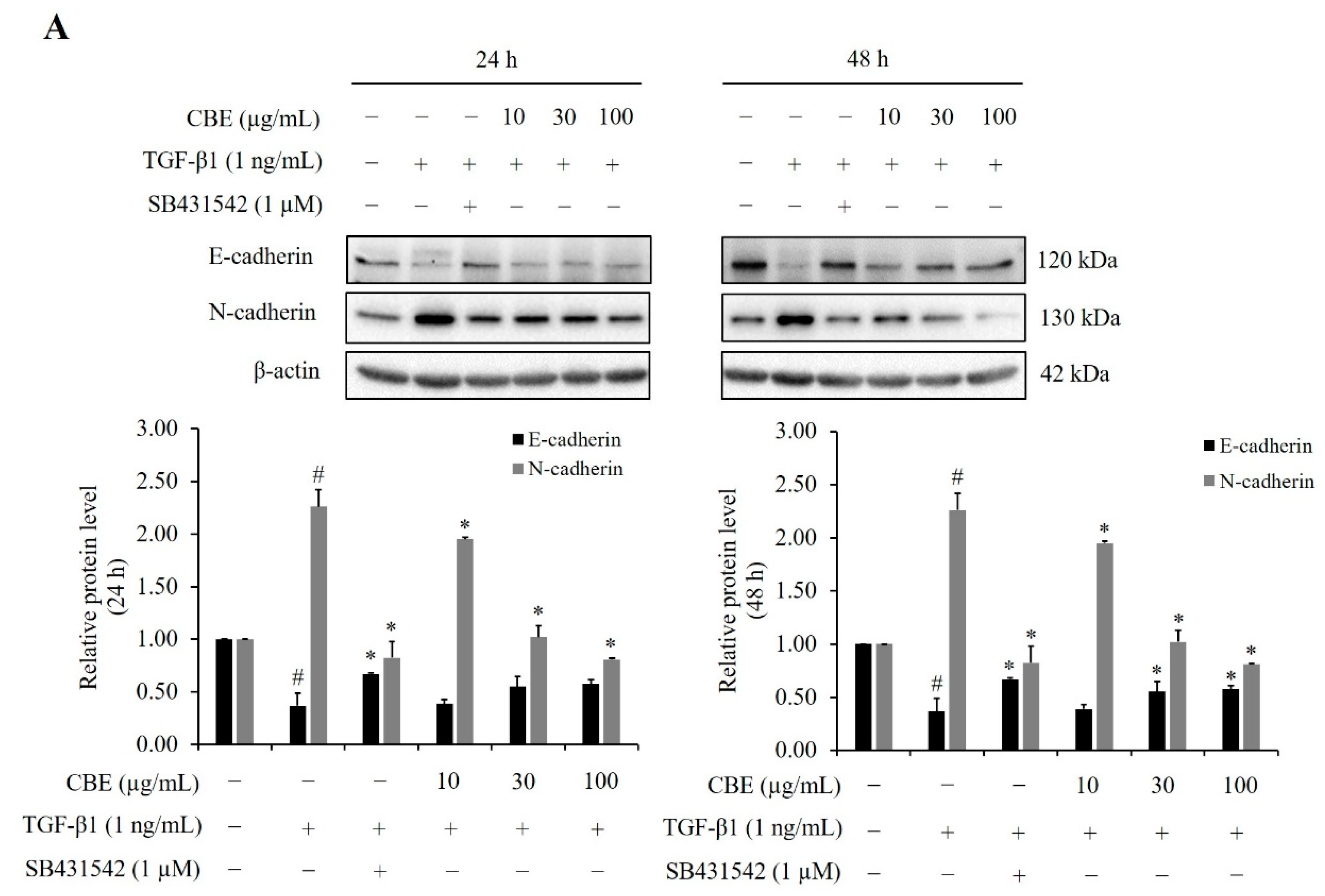
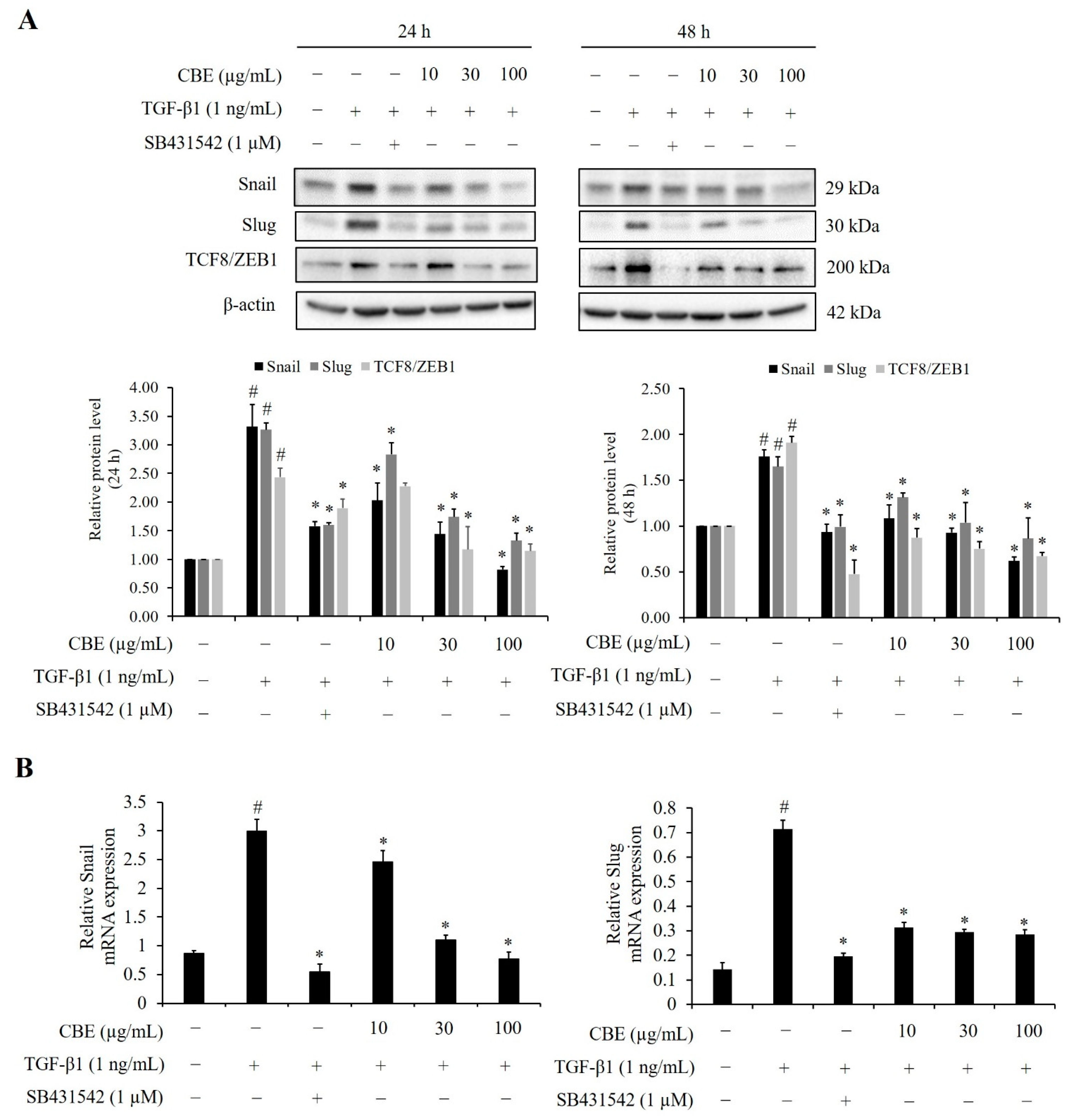
2.4. Effect of CBE on TGF-β-Induced EMT-Related Biomarker Protein Expression
2.5. CBE Reduces TGF-β-Induced EMT-Regulated Transcription Factors
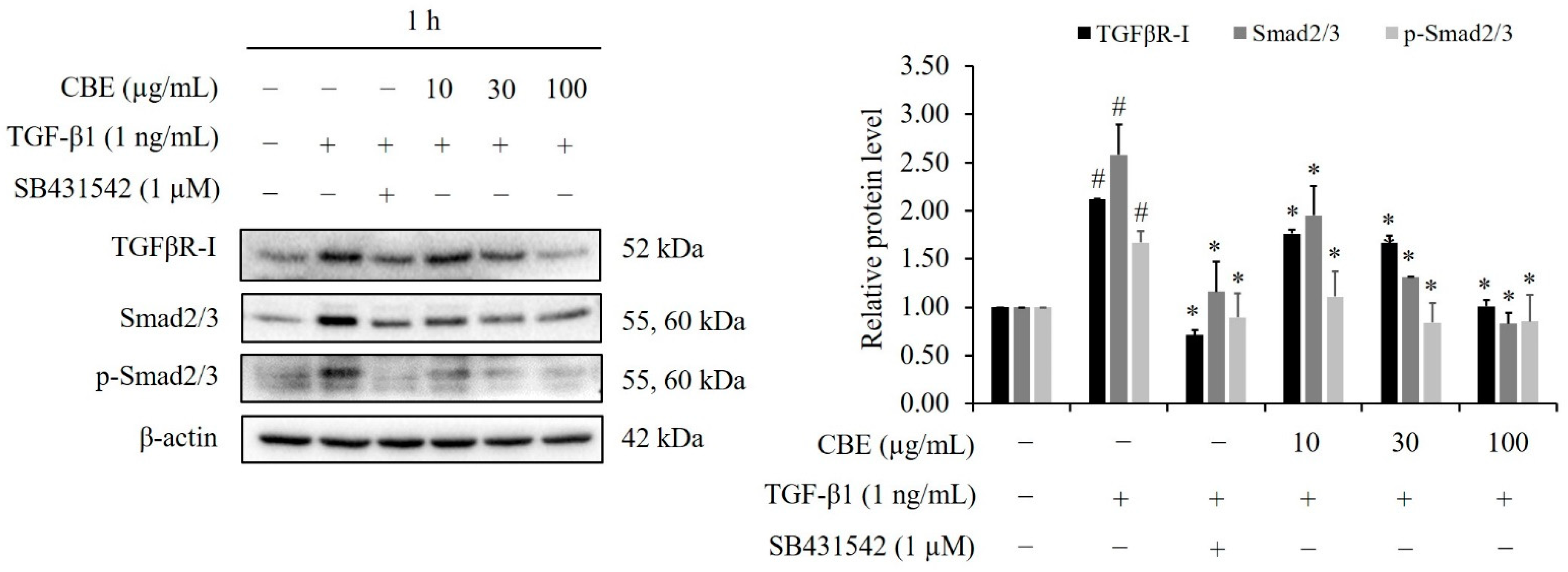
2.6. CBE Reduces TGF-β1-Induced Smad2/3 Phosphorylation
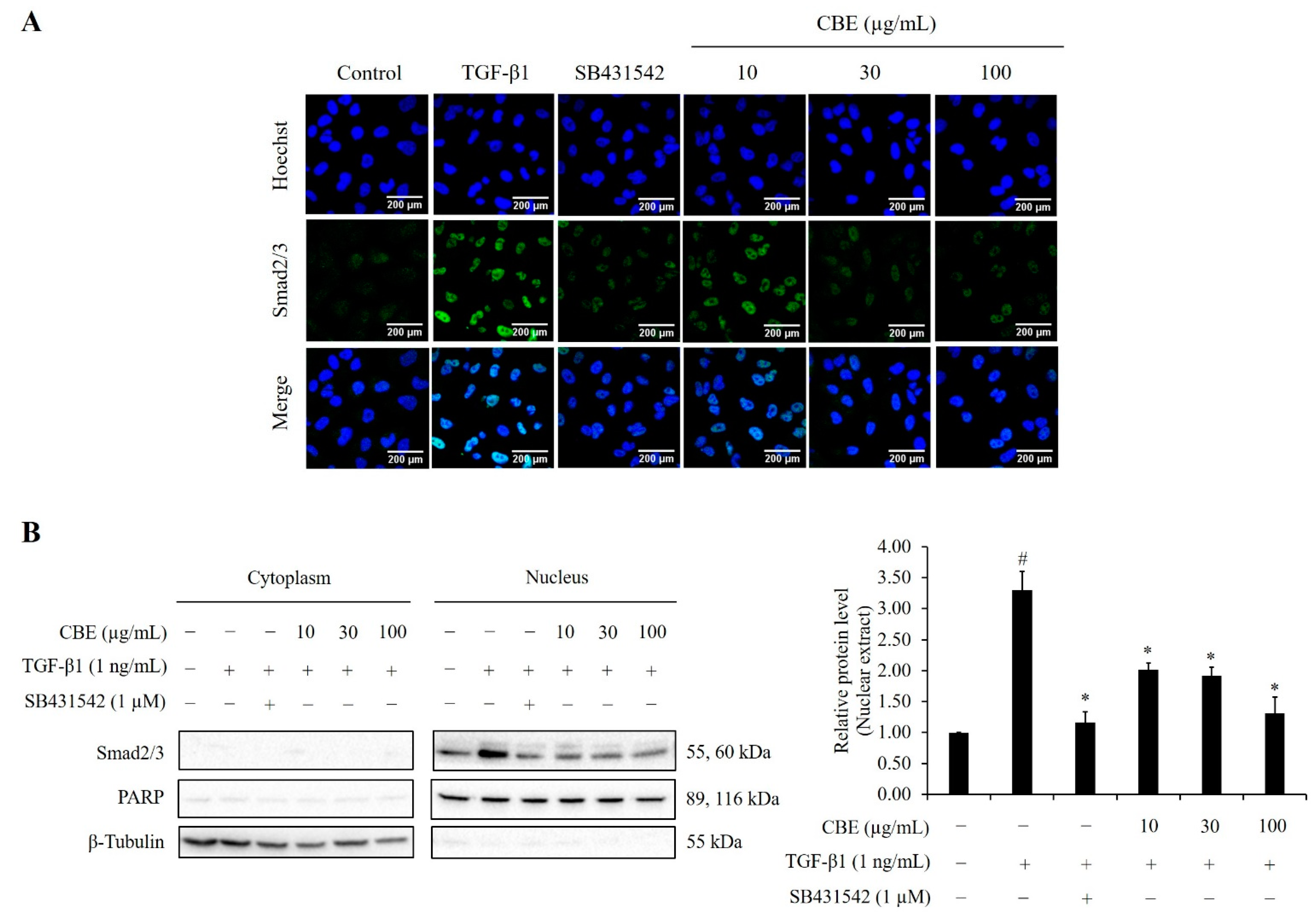
2.7. CBE Reduces TGF-β1-Induced Smad2/3 Nuclear Translocation
2.8. CBE Reduces TGF-β1-Induced EMT via a Smad-Independent Pathway
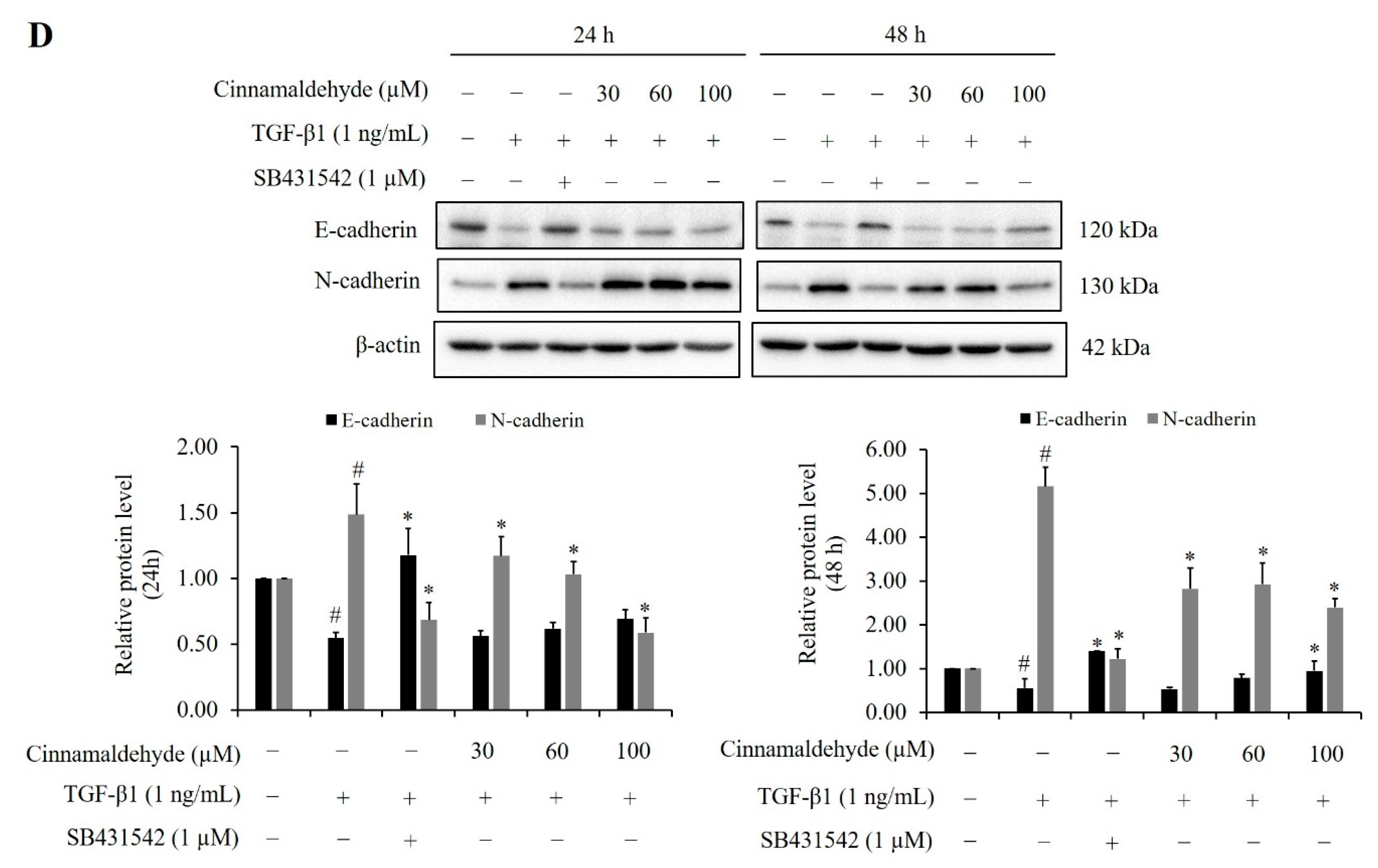
2.9. Effect of Cinnamaldehyde on TGF-β1-Induced Smad2/3 Phosphorylation and EMT Markers Expression
3. Discussion
4. Materials and Methods
4.1. Preparation of C. bejolghota Ethanolic Bark Extract
4.2. Chemicals and Reagents
4.3. Cell Culture
4.4. Cytotoxicity Assay
4.5. Scratch Assay
4.6. Transwell Assay
4.7. Cell Adhesion Assay
4.8. Morphological Assessment
4.9. Western Blot Analysis
4.10. RNA Extraction, RT-PCR, and Quantitative Real-Time PCR (qPCR)
4.11. Immunofluorescence Analysis
4.12. Cytoplasmic/Nuclear Fractionation Assay
4.13. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sung, H.; Ferlay, J.; Siegel, R.L. Global cancer statistics 2020: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef] [PubMed]
- Bundgaard-Nielsen, C.; Baandrup, U.T.; Nielsen, L.P.; Sørensen, S. The presence of bacteria varies between colorectal adenocarcinomas, precursor lesions and non-malignant tissue. BMC Cancer 2019, 19, 399. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Z.; Cao, Q.; Dai, G.; Wang, J.; Liu, C.; Lv, L.; Pan, J. Celastrol inhibits colorectal cancer through TGF-β1/Smad signalling. Onco Targets Ther. 2019, 119, 509–518. [Google Scholar] [CrossRef] [Green Version]
- Kalluri, R.; Weinberg, R.A. The basics of epithelial-mesenchymal transition. J. Clin. Investig. 2009, 119, 1420–1428. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Araki, K.; Shimura, H.; Suzuki, H.; Tsutsumi, S.; Wada, W.; Yajima, T.; Kobayahi, T.; Kubo, N.; Kuwano, H. E/N-cadherin switch mediates cancer progression via TGF-β-induced epithelial-to-mesenchymal transition in extrahepatic cholangiocarcinoma. Br. J. Cancer 2011, 105, 1885–1893. [Google Scholar] [CrossRef] [Green Version]
- Yan, M.D.; Yao, C.J.; Chow, J.M.; Chang, C.L.; Hwang, P.A.; Chuang, S.E.; Whang-Peng, J.; Lai, G.M. Fucoidan elevates microRNA-29b to regulate DNMT3B-MTSS1 axis and inhibit EMT in human hepatocellular carcinoma cells. Mar. Drugs 2015, 13, 6099–6116. [Google Scholar] [CrossRef] [Green Version]
- Xu, J.; Lamouille, S.; Derynck, R. TGF-beta-induced epithelial to mesenchymal transition. Cell Res. 2009, 19, 156–172. [Google Scholar] [CrossRef] [PubMed]
- Kubiczkova, L.; Sedlarikova, L.; Hajek, R.; Sevcikova, S. TGF-β—An excellent servant but a bad master. J. Transl. Med. 2012, 10, 183. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hata, A.; Chen, Y.G. TGF-β signaling from Receptors to Smads. Cold Spring Harb. Perspect. Biol. 2016, 8, a022061. [Google Scholar] [CrossRef] [Green Version]
- Nagaraj, N.S.; Datta, P.K. Targeting the transforming growth factor-beta signaling pathway in human cancer. Expert Opin. Investig. Drugs 2010, 19, 77–91. [Google Scholar] [CrossRef] [Green Version]
- Rani, A.; Pande, C.; Tewari, G.; Patni, K. A review on aroma profile of Cinnamomum species in north and north east India. World J. Pharm. Res. 2017, 6, 200–221. [Google Scholar]
- Gogoi, B.; Kakoti, B.; Bora, S.; Borah, N.S. Pharmacognostic and preliminary phytochemical evaluation of Cinnamomum bejolghota (Buch.-Ham.) sweet bark. Indian J. Nat. Prod. Resour. 2016, 7, 59–64. [Google Scholar]
- Atiphasaworn, P.; Monggoot, S.; Pripdeevech, P. Chemical composition, antibacterial and antifungal activities of Cinamomum bejolghota bark oil from Thailand. J. Appl. Pharm. 2017, 7, 69–73. [Google Scholar]
- Gogoi, B.; Kakoti, B.; Bora, N.S.; Borah, S. Antihyperglycemic and in vivo antioxidative activity evaluation of Cinnamomum bejolghota (Buch.-Ham.) in streptozotocin induced diabetic rats: An ethnomedicinal plant in Assam. Asian Pac. J. Trop. Med. 2014, 7, S427–S434. [Google Scholar] [CrossRef] [Green Version]
- Rao, L.; You, Y.X.; Su, Y.; Fan, Y.; Liu, Y.; He, Q.; Chen, Y.; Meng, J.; Hu, L.; Li, Y.; et al. Lignans and neolignans with antioxidant and human cancer cell proliferation inhibitory activities from Cinnamomum bejolghota confirm its functional food property. J. Agric. Food Chem. 2020, 68, 8825–8835. [Google Scholar] [CrossRef] [PubMed]
- Kryczka, J.; Stasiak, M.; Dziki, L.; Mik, M.; Dziki, A.; Cierniewski, C. Matrix metalloproteinase-2 cleavage of the β1 integrin ectodomain facilitates colon cancer cell motility. J. Biol. Chem. 2012, 287, 36556–36566. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ansieau, S.; Collin, G.; Hill, L. EMT or EMT-Promoting Transcription Factors, Where to Focus the Light? Front. Oncol. 2014, 4, 353. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, L. Regulation of Smad activities. Biochim. Biophys. Acta (BBA) Gene Struct. Expr. 2006, 1759, 503–513. [Google Scholar] [CrossRef] [Green Version]
- Koppikar, S.J.; Choudhari, A.S.; Suryavanshi, S.A.; Kumari, S.; Chattopadhyay, S.; Kaul-Ghanekar, R. Aqueous cinnamon extract (ACE-c) from the bark of Cinnamomum cassia causes apoptosis in human cervical cancer cell line (SiHa). BMC Cancer 2010, 10, 210. [Google Scholar] [CrossRef] [Green Version]
- Lin, C.Y.; Hsieh, Y.H.; Yang, S.F.; Chu, S.C.; Chen, P.N.; Hsieh, Y.S. Cinnamomum cassia extracts reverses TGF-β1-induced epithelial-mesenchymal transition in human lung adenocarcinoma cells and suppresses tumor growth in vivo. Environ. Toxicol. 2017, 32, 1878–1887. [Google Scholar] [CrossRef]
- Kee, J.Y.; Han, Y.H.; Mun, J.G.; Park, S.H.; Jeon, H.D.; Hong, S.H. Effect of Korean Red Ginseng extract on colorectal lung metastasis through inhibiting the epithelial-mesenchymal transition via transforming growth factor-β1/Smad-signaling-mediated Snail/E-cadherin expression. J. Ginseng Res. 2019, 43, 68–76. [Google Scholar] [CrossRef] [PubMed]
- Hu, B.; An, H.M.; Yan, X.; Zheng, J.L.; Huang, X.W.; Li, M. Traditional Chinese medicine formulation Yanggan Jiedu Sanjie inhibits TGF-β1-induced epithelial-mesenchymal transition and metastatic potential in human hepatocarcinoma Bel-7402 cells. BMC Complementary Altern. Med. 2019, 19, 67. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santibanez, J.F.; Obradović, H.; Kukolj, T.; Krstić, J. Transforming growth factor-β, matrix metalloproteinases, and urokinase-type plasminogen activator interaction in the cancer epithelial to mesenchymal transition. Dev. Dyn. 2018, 247, 382–395. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hwang, K.E.; Kim, H.J.; Song, I.S.; Park, C.; Jung, J.W.; Park, D.S.; Oh, S.H.; Kim, Y.S.; Kim, H.R. Salinomycin suppresses TGF-β1-induced EMT by down-regulating MMP-2 and MMP-9 via the AMPK/SIRT1 pathway in non-small cell lung cancer. Int. J. Med. Sci. 2021, 18, 715–726. [Google Scholar] [CrossRef]
- Wang, N.; Zhu, M.; Tsao, S.W.; Man, K.; Zhang, Z.; Feng, Y. Up-regulation of TIMP-1 by genipin inhibits MMP-2 activities and suppresses the metastatic potential of human hepatocellular carcinoma. PLoS ONE 2012, 7, e46318. [Google Scholar] [CrossRef] [Green Version]
- Lian, G.Y.; Wang, Q.M.; Mak, T.S.; Huang, X.R.; Yu, X.Q.; Lan, H.Y. Inhibition of tumor invasion and metastasis by targeting TGF-β-Smad-MMP2 pathway with Asiatic acid and Naringenin. Mol. Ther.-Oncolytics 2021, 20, 277–289. [Google Scholar] [CrossRef]
- Brabletz, T.; Kalluri, R.; Nieto, M.A.; Weinberg, R.A. EMT in cancer. Nat. Rev. Cancer 2018, 18, 128–134. [Google Scholar] [CrossRef]
- Zhu, S.M.; Park, Y.R.; Seo, S.Y.; Kim, I.H.; Lee, S.T.; Kim, S.W. Parthenolide inhibits transforming growth factor β1-induced epithelial-mesenchymal transition in colorectal cancer cells. Intest. Res. 2019, 17, 527–536. [Google Scholar] [CrossRef] [Green Version]
- Rouillard, A.D.; Gundersen, G.W.; Fernandez, N.F.; Wang, Z.; Monteiro, C.D.; McDermott, M.G.; Ma’ayan, A. The harmonizome: A collection of processed datasets gathered to serve and mine knowledge about genes and proteins. Database 2016, 2016, baw100. [Google Scholar] [CrossRef]
- Hao, Y.; Baker, D.; Ten Dijke, P. TGF-β-Mediated Epithelial-Mesenchymal Transition and Cancer Metastasis. Int. J. Mol. Sci. 2019, 20, 2767. [Google Scholar] [CrossRef] [Green Version]
- Ji, Q.; Liu, X.; Han, Z.; Zhou, L.; Sui, H.; Yan, L.; Jiang, H.; Ren, J.; Cai, J.; Li, Q. Resveratrol suppresses epithelial-to-mesenchymal transition in colorectal cancer through TGF-β1/Smads signaling pathway mediated Snail/E-cadherin expression. BMC Cancer 2015, 15, 97. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zare, Z.; Nayerpour Dizaj, T.; Lohrasbi, A.; Sheikhalishahi, Z.S.; Asadi, A.; Zakeri, M.; Hosseinabadi, F.; Abazari, O.; Abbasi, M.; Khanicheragh, P. Silibinin Inhibits TGF-β-induced MMP-2 and MMP-9 Through Smad Signaling Pathway in Colorectal Cancer HT-29 Cells. Basic Clin. Cancer Res. 2021, 12, 81–90. [Google Scholar] [CrossRef]
- Massagué, J.; Wotton, D. Transcriptional control by the TGF-beta/Smad signaling system. EMBO J. 2000, 19, 1745–1754. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, X.N.; Wang, S.; Yang, Q.; Wang, Y.J.; Chen, D.X.; Zhu, X.X. ESC reverses epithelial mesenchymal transition induced by transforming growth factor-β via inhibition of Smad signal pathway in HepG2 liver cancer cells. Cancer Cell Int. 2015, 15, 114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wendt, M.K.; Allington, T.M.; Schiemann, W.P. Mechanisms of the epithelial-mesenchymal transition by TGF-beta. Future Oncol. 2009, 5, 1145–1168. [Google Scholar] [CrossRef] [Green Version]
- Choudhury, S.; Ahmed, R.; Barthel, A.; Leclercq, P.A. Composition of the Bark and Flower Oils of Cinnamomum bejolghota (Buch.-Ham.) Sweet from two Locations of Assam, India. J. Essent. Oil Res. 1998, 10, 245–250. [Google Scholar] [CrossRef]
- Kittiwattanokhun, A.; Samosorn, S.; Innajak, S.; Watanapokasin, R. Inhibitory effects on chondrosarcoma cell metastasis by Senna alata extract. Biomed. Pharmacother. 2021, 137, 111337. [Google Scholar] [CrossRef]
- Utchariyakiat, I.; Surassmo, S.; Jaturanpinyo, M.; Khuntayaporn, P.; Chomnawang, M.T. Efficacy of cinnamon bark oil and cinnamaldehyde on anti-multidrug resistant Pseudomonas aeruginosa and the synergistic effects in combination with other antimicrobial agents. BMC Complementary Altern. Med. 2016, 16, 158. [Google Scholar] [CrossRef] [Green Version]
- Sadeghi, Z.; Valizadeh, J.; Azyzian Shermeh, O.; Akaberi, M. Antioxidant activity and total phenolic content of Boerhavia elegans (choisy) grown in Baluchestan, Iran. Avicenna J. Phytomed. 2015, 5, 1–9. [Google Scholar]












Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kittiwattanokhun, A.; Innajak, S.; Tashiro, E.; Imoto, M.; Watanapokasin, R. Cinnamomum bejolghota Extract Inhibits Colorectal Cancer Cell Metastasis and TGF-β1-Induced Epithelial-Mesenchymal Transition via Smad and Non-Smad Signaling Pathway. Sci. Pharm. 2022, 90, 30. https://doi.org/10.3390/scipharm90020030
Kittiwattanokhun A, Innajak S, Tashiro E, Imoto M, Watanapokasin R. Cinnamomum bejolghota Extract Inhibits Colorectal Cancer Cell Metastasis and TGF-β1-Induced Epithelial-Mesenchymal Transition via Smad and Non-Smad Signaling Pathway. Scientia Pharmaceutica. 2022; 90(2):30. https://doi.org/10.3390/scipharm90020030
Chicago/Turabian StyleKittiwattanokhun, Athicha, Sukanda Innajak, Etsu Tashiro, Masaya Imoto, and Ramida Watanapokasin. 2022. "Cinnamomum bejolghota Extract Inhibits Colorectal Cancer Cell Metastasis and TGF-β1-Induced Epithelial-Mesenchymal Transition via Smad and Non-Smad Signaling Pathway" Scientia Pharmaceutica 90, no. 2: 30. https://doi.org/10.3390/scipharm90020030
APA StyleKittiwattanokhun, A., Innajak, S., Tashiro, E., Imoto, M., & Watanapokasin, R. (2022). Cinnamomum bejolghota Extract Inhibits Colorectal Cancer Cell Metastasis and TGF-β1-Induced Epithelial-Mesenchymal Transition via Smad and Non-Smad Signaling Pathway. Scientia Pharmaceutica, 90(2), 30. https://doi.org/10.3390/scipharm90020030