Application of Carbon-13 Isotopomer Analysis to Assess Perinatal Myocardial Glucose Metabolism in Sheep

 ,
,
Abstract
:
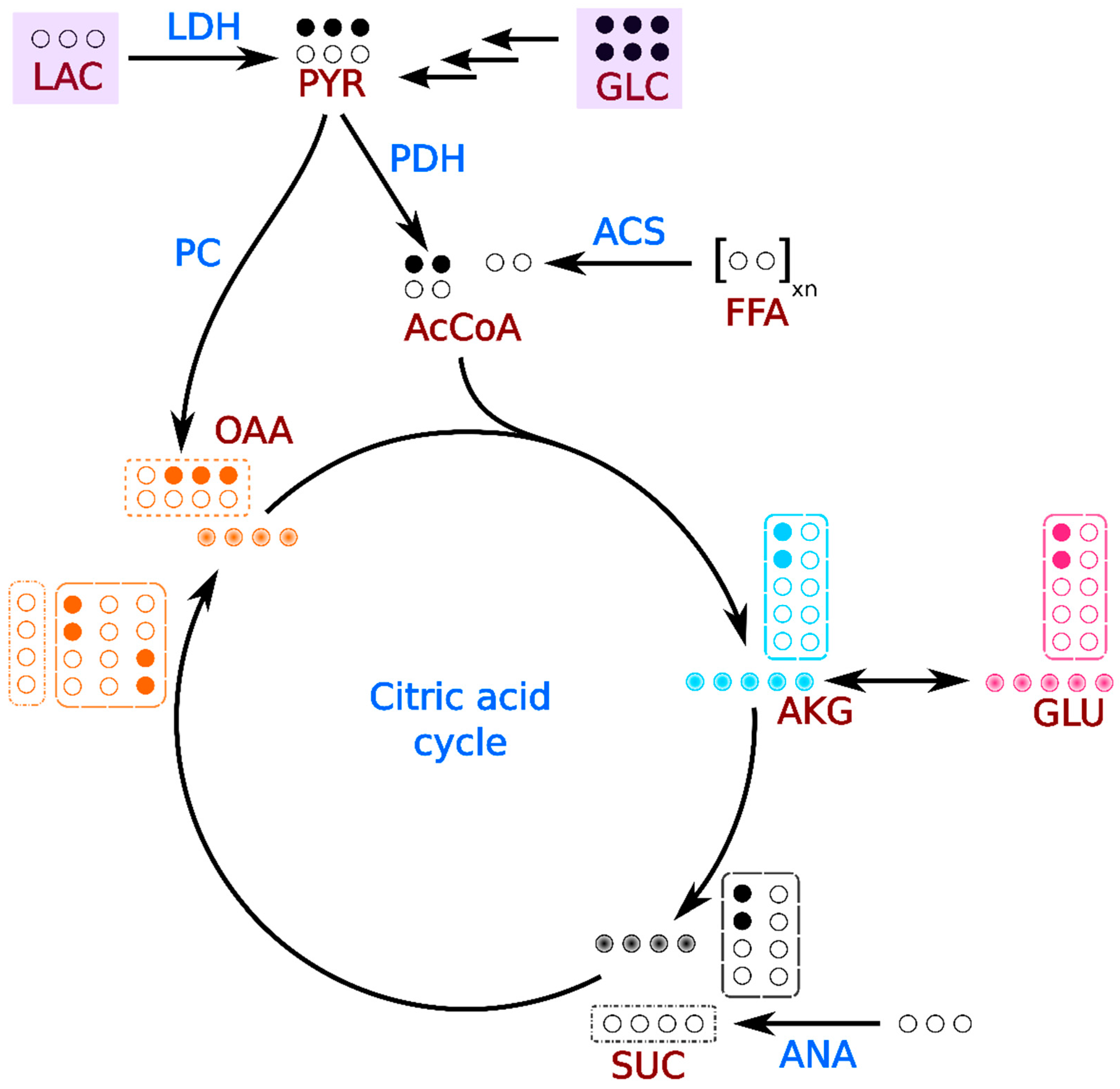
1. Introduction
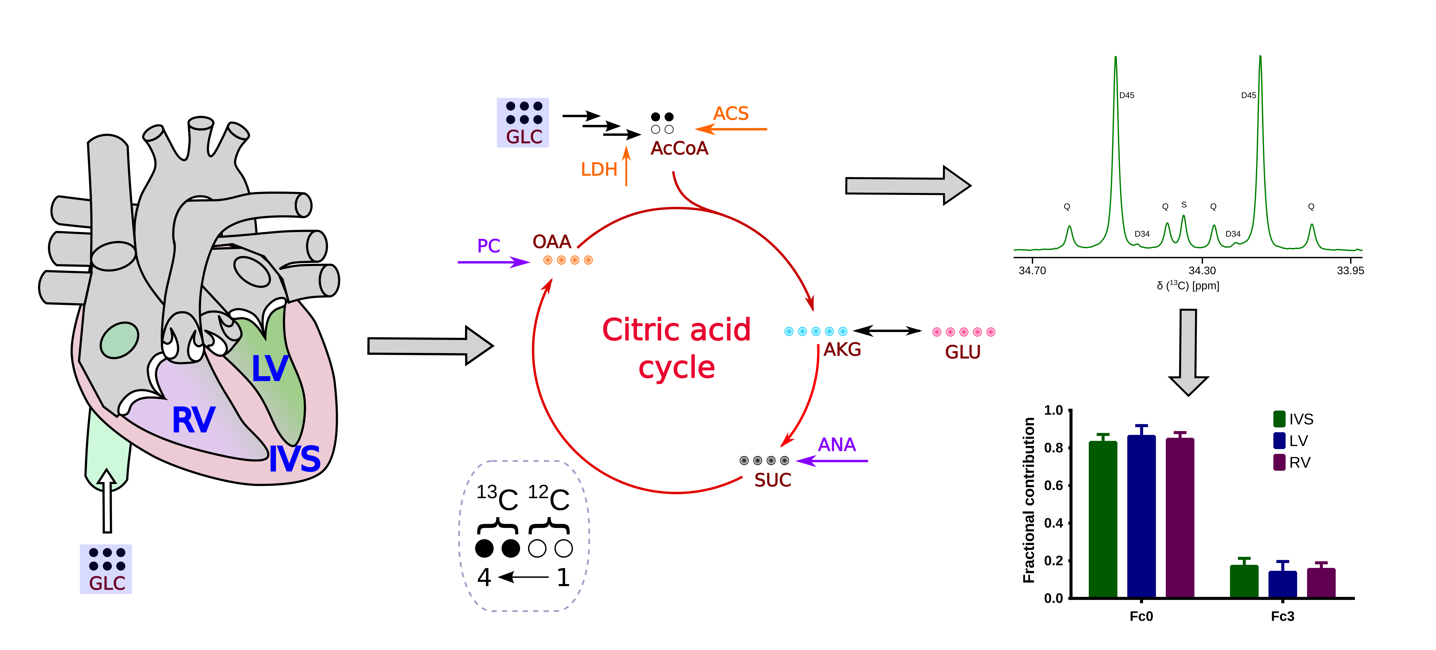
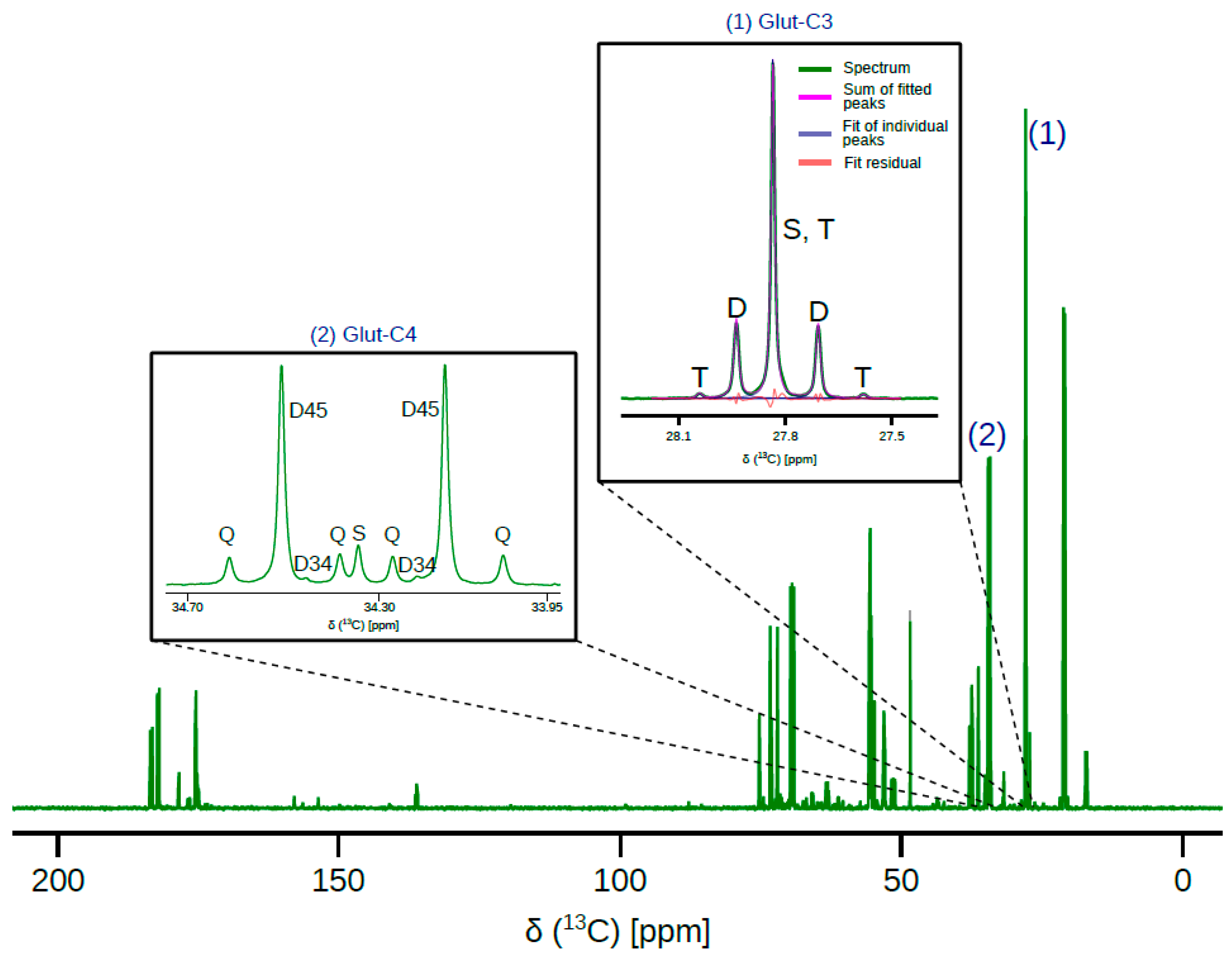
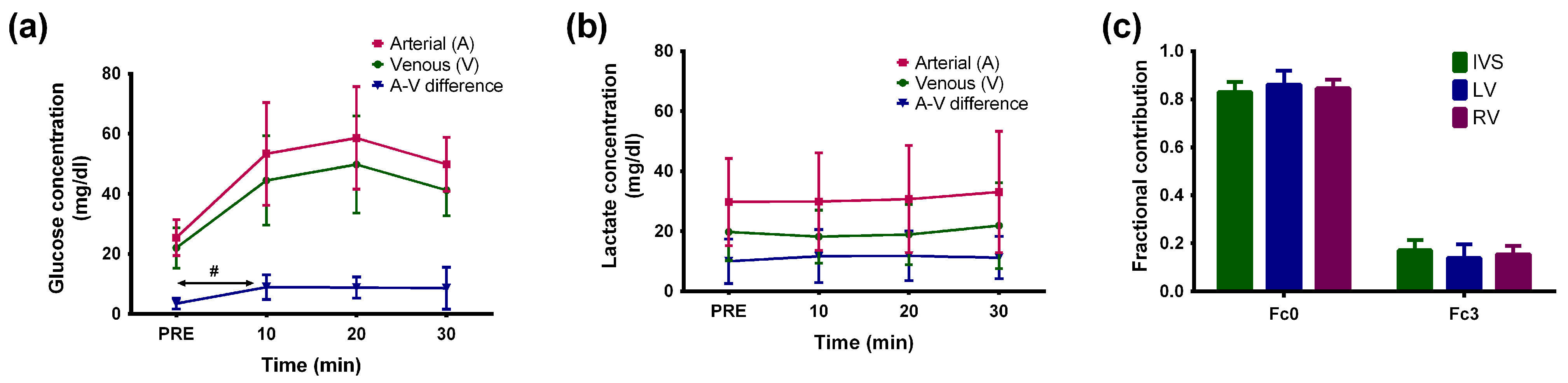
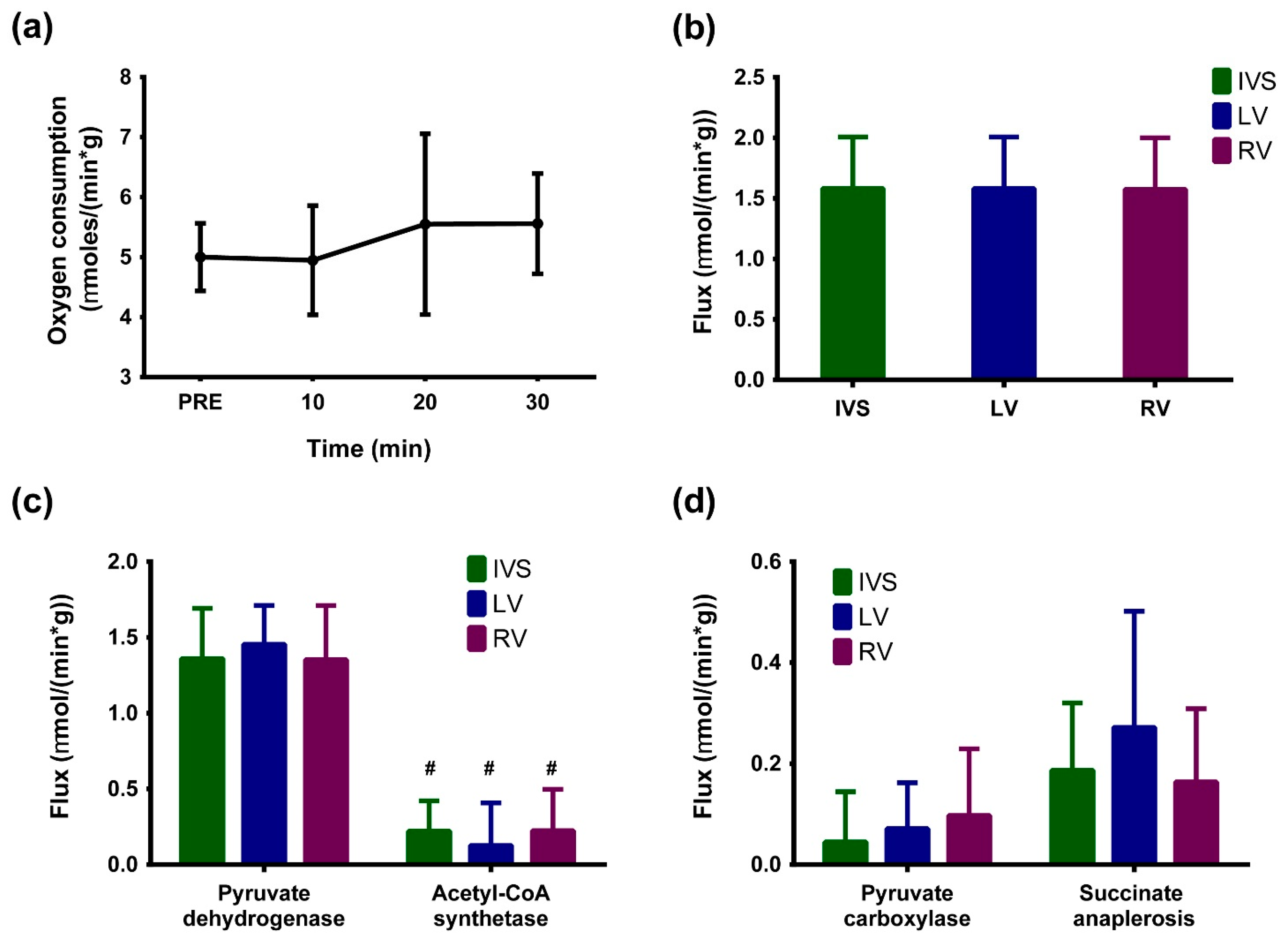
2. Results
3. Discussion
4. Materials and Methods
4.1. Animal Use
4.2. Extraction of Metabolites
4.3. NMR Spectra Acquisition
4.4. Steady State Analysis
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Conflicts of Interest
References
- Lopaschuk, G.D.; Spafford, M.A. Energy substrate utilization by isolated working hearts from newborn rabbits. Am. J. Physiol.-Heart Circ. Physiol. 1990, 258, H1274–H1280. [Google Scholar] [CrossRef] [PubMed]
- Lopaschuk, G.D.; Spafford, M.A.; Marsh, D.R. Glycolysis is predominant source of myocardial ATP production immediately after birth. Am. J. Physiol.-Heart Circ. Physiol. 1991, 261, H1698–H1705. [Google Scholar] [CrossRef] [PubMed]
- Bartelds, B.; Knoester, H.; Beaufort-Krol, G.C.M.; Smid, G.B.; Takens, J.; Zijlstra, W.G.; Heymans, H.S.A.; Kuipers, J.R.G. Myocardial Lactate Metabolism in Fetal and Newborn Lambs. Circulation 1999, 99, 1892–1897. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jonker, S.S.; Zhang, L.; Louey, S.; Giraud, G.D.; Thornburg, K.L.; Faber, J.J. Myocyte enlargement, differentiation, and proliferation kinetics in the fetal sheep heart. J. Appl. Physiol. 2007, 102, 1130–1142. [Google Scholar] [CrossRef] [Green Version]
- Camacho, P.; Fan, H.; Liu, Z.; He, J.-Q. Large Mammalian Animal Models of Heart Disease. J. Cardiovasc. Dev. Dis. 2016, 3, 30. [Google Scholar] [CrossRef] [Green Version]
- Jeffrey, F.M.; Storey, C.J.; Sherry, A.D.; Malloy, C.R. 13C isotopomer model for estimation of anaplerotic substrate oxidation via acetyl-CoA. Am. J. Physiol.-Endocrinol. Metab. 1996, 271, E788–E799. [Google Scholar] [CrossRef]
- Chance, E.M.; Seeholzer, S.H.; Kobayashi, K.; Williamson, J.R. Mathematical analysis of isotope labeling in the citric acid cycle with applications to 13C NMR studies in perfused rat hearts. J. Biol. Chem. 1983, 258, 13785–13794. [Google Scholar] [CrossRef]
- Ragavan, M.; Kirpich, A.; Fu, X.; Burgess, S.C.; McIntyre, L.M.; Merritt, M.E. A comprehensive analysis of myocardial substrate preference emphasizes the need for a synchronized fluxomic/metabolomic research design. Am. J. Physiol.-Heart Circ. Physiol. 2017, 312, H1215–H1223. [Google Scholar] [CrossRef] [Green Version]
- Burgess, S.C.; Jeffrey, F.M.H.; Storey, C.; Milde, A.; Hausler, N.; Merritt, M.E.; Mulder, H.; Holm, C.; Sherry, A.D.; Malloy, C.R. Effect of murine strain on metabolic pathways of glucose production after brief or prolonged fasting. Am. J. Physiol.-Endocrinol. Metab. 2005, 289, E53–E61. [Google Scholar] [CrossRef] [Green Version]
- Jin, E.S.; Sherry, A.D.; Malloy, C.R. Interaction between the Pentose Phosphate Pathway and Gluconeogenesis from Glycerol in the Liver. J. Biol. Chem. 2014, 289, 32593–32603. [Google Scholar] [CrossRef] [Green Version]
- Mashimo, T.; Pichumani, K.; Vemireddy, V.; Hatanpaa, K.J.; Singh, D.K.; Sirasanagandla, S.; Nannepaga, S.; Piccirillo, S.G.; Kovacs, Z.; Foong, C.; et al. Acetate Is a Bioenergetic Substrate for Human Glioblastoma and Brain Metastases. Cell 2014, 159, 1603–1614. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fisher, D.J.; Heymann, M.A.; Rudolph, A.M. Myocardial oxygen and carbohydrate consumption in fetal lambs in utero and in adult sheep. Am. J. Physiol.-Heart Circ. Physiol. 1980, 238, H399–H405. [Google Scholar] [CrossRef]
- Fisher, D.J.; Heymann, M.A.; Rudolph, A.M. Fetal myocardial oxygen and carbohydrate consumption during acutely induced hypoxemia. Am. J. Physiol.-Heart Circ. Physiol. 1982, 242, H657–H661. [Google Scholar] [CrossRef]
- Richards, E.M.; Wood, C.E.; Rabaglino, M.B.; Antolic, A.; Keller-Wood, M. Mechanisms for the adverse effects of late gestational increases in maternal cortisol on the heart revealed by transcriptomic analyses of the fetal septum. Physiol. Genom. 2014, 46, 547–559. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pound, K.M.; Sorokina, N.; Ballal, K.; Berkich, D.A.; Fasano, M.; LaNoue, K.F.; Taegtmeyer, H.; O’Donnell, J.M.; Lewandowski, E.D. Substrate–Enzyme Competition Attenuates Upregulated Anaplerotic Flux Through Malic Enzyme in Hypertrophied Rat Heart and Restores Triacylglyceride Content Attenuating Upregulated Anaplerosis in Hypertrophy. Circ. Res. 2009, 104, 805–812. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Malloy, C.R.; Thompson, J.R.; Jeffrey, F.M.H.; Sherry, A.D. Contribution of exogenous substrates to acetyl coenzyme A: Measurement by carbon-13 NMR under non-steady-state conditions. Biochemistry 1990, 29, 6756–6761. [Google Scholar] [CrossRef] [PubMed]
- Milani-Nejad, N.; Janssen, P.M.L. Small and Large Animal Models in Cardiac Contraction Research: Advantages and Disadvantages. Pharm. Ther. 2014, 141, 235–249. [Google Scholar] [CrossRef] [Green Version]
- Turer, A.; Altamirano, F.; Schiattarella, G.G.; May, H.; Gillette, T.G.; Malloy, C.R.; Merritt, M.E. Remodeling of substrate consumption in the murine sTAC model of heart failure. J. Mol. Cell. Cardiol. 2019, 134, 144–153. [Google Scholar] [CrossRef]
- Lehman, J.J.; Kelly, D.P. Gene Regulatory Mechanisms Governing Energy Metabolism during Cardiac Hypertrophic Growth. Heart Fail. Rev. 2002, 7, 175–185. [Google Scholar] [CrossRef]
- Walejko, J.M.; Koelmel, J.P.; Garrett, T.J.; Edison, A.S.; Keller-Wood, M. Multiomics approach reveals metabolic changes in the heart at birth. Am. J. Physiol.-Endocrinol. Metab. 2018, 315, E1212–E1223. [Google Scholar] [CrossRef] [Green Version]
- Antolic, A.; Richards, E.M.; Wood, C.E.; Keller-Wood, M. A Transcriptomic Model of Postnatal Cardiac Effects of Prenatal Maternal Cortisol Excess in Sheep. Front. Physiol. 2019, 10, 816. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aldoretta, P.W.; Carver, T.D.; Hay, W.W., Jr. Maturation of Glucose-Stimulated Insulin Secretion in Fetal Sheep. Neonatology 1998, 73, 375–386. [Google Scholar] [CrossRef] [PubMed]
- Reinhardt, C.P.; Dalhberg, S.; Tries, M.A.; Marcel, R.; Leppo, J.A. Stable labeled microspheres to measure perfusion: Validation of a neutron activation assay technique. Am. J. Physiol.-Heart Circ. Physiol. 2001, 280, H108–H116. [Google Scholar] [CrossRef] [PubMed]
- Heymann, M.A.; Payne, B.D.; Hoffman, J.I.E.; Rudolph, A.M. Blood flow measurements with radionuclide-labeled particles. Prog. Cardiovasc. Dis. 1977, 20, 55–79. [Google Scholar] [CrossRef]
- Ramaswamy, V.; Hooker, J.W.; Withers, R.S.; Nast, R.E.; Brey, W.W.; Edison, A.S. Development of a 13C-optimized 1.5-mm high temperature superconducting NMR probe. J. Magn. Reson. 2013, 235, 58–65. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sherry, A.D.; Jeffrey, F.M.H.; Malloy, C.R. Analytical solutions for 13C isotopomer analysis of complex metabolic conditions: Substrate oxidation, multiple pyruvate cycles, and gluconeogenesis. Metab. Eng. 2004, 6, 12–24. [Google Scholar] [CrossRef] [PubMed]
- Malloy, C.R.; Jones, J.G.; Jeffrey, F.M.; Jessen, M.E.; Sherry, A.D. Contribution of various substrates to total citric acid cycle flux and ]anaplerosis as determined by13C isotopomer analysis and O2 consumption in the heart. Magn. Reson. Mater. Phys. Biol. Med. 1996, 4, 35–46. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Explanation | Starting Value (Range) |
|---|---|---|
| lac123 | 13C enrichment of lactate (derived from [U-13C]glucose | Data from 1H NMR spectra |
| pdh | Pyruvate dehydrogenase flux | 0.3 (0–1) |
| ys | Anaplerosis through succinate | 0.3 (>= 0) |
| ypc | Pyruvate carboxylase | 0.05 (>= 0) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ragavan, M.; Li, M.; Giacalone, A.G.; Wood, C.E.; Keller-Wood, M.; Merritt, M.E. Application of Carbon-13 Isotopomer Analysis to Assess Perinatal Myocardial Glucose Metabolism in Sheep. Metabolites 2021, 11, 33. https://doi.org/10.3390/metabo11010033
Ragavan M, Li M, Giacalone AG, Wood CE, Keller-Wood M, Merritt ME. Application of Carbon-13 Isotopomer Analysis to Assess Perinatal Myocardial Glucose Metabolism in Sheep. Metabolites. 2021; 11(1):33. https://doi.org/10.3390/metabo11010033
Chicago/Turabian StyleRagavan, Mukundan, Mengchen Li, Anthony G. Giacalone, Charles E. Wood, Maureen Keller-Wood, and Matthew E. Merritt. 2021. "Application of Carbon-13 Isotopomer Analysis to Assess Perinatal Myocardial Glucose Metabolism in Sheep" Metabolites 11, no. 1: 33. https://doi.org/10.3390/metabo11010033
APA StyleRagavan, M., Li, M., Giacalone, A. G., Wood, C. E., Keller-Wood, M., & Merritt, M. E. (2021). Application of Carbon-13 Isotopomer Analysis to Assess Perinatal Myocardial Glucose Metabolism in Sheep. Metabolites, 11(1), 33. https://doi.org/10.3390/metabo11010033