
The Interface amongst Conserved and Specialized Pathways in Non-Paclitaxel and Paclitaxel Accumulating Taxus Cultures

Abstract
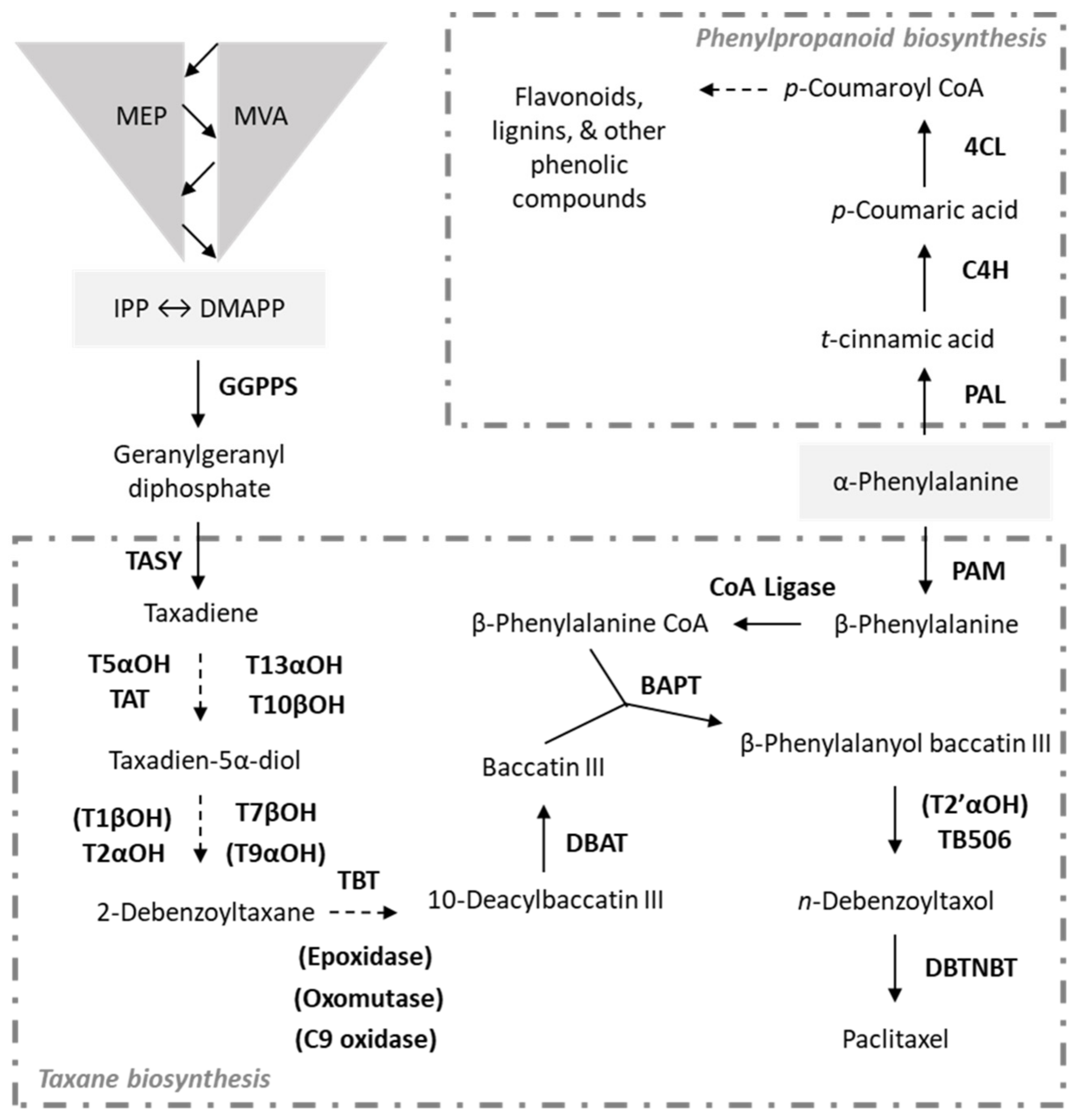
:1. Introduction
2. Results and Discussion
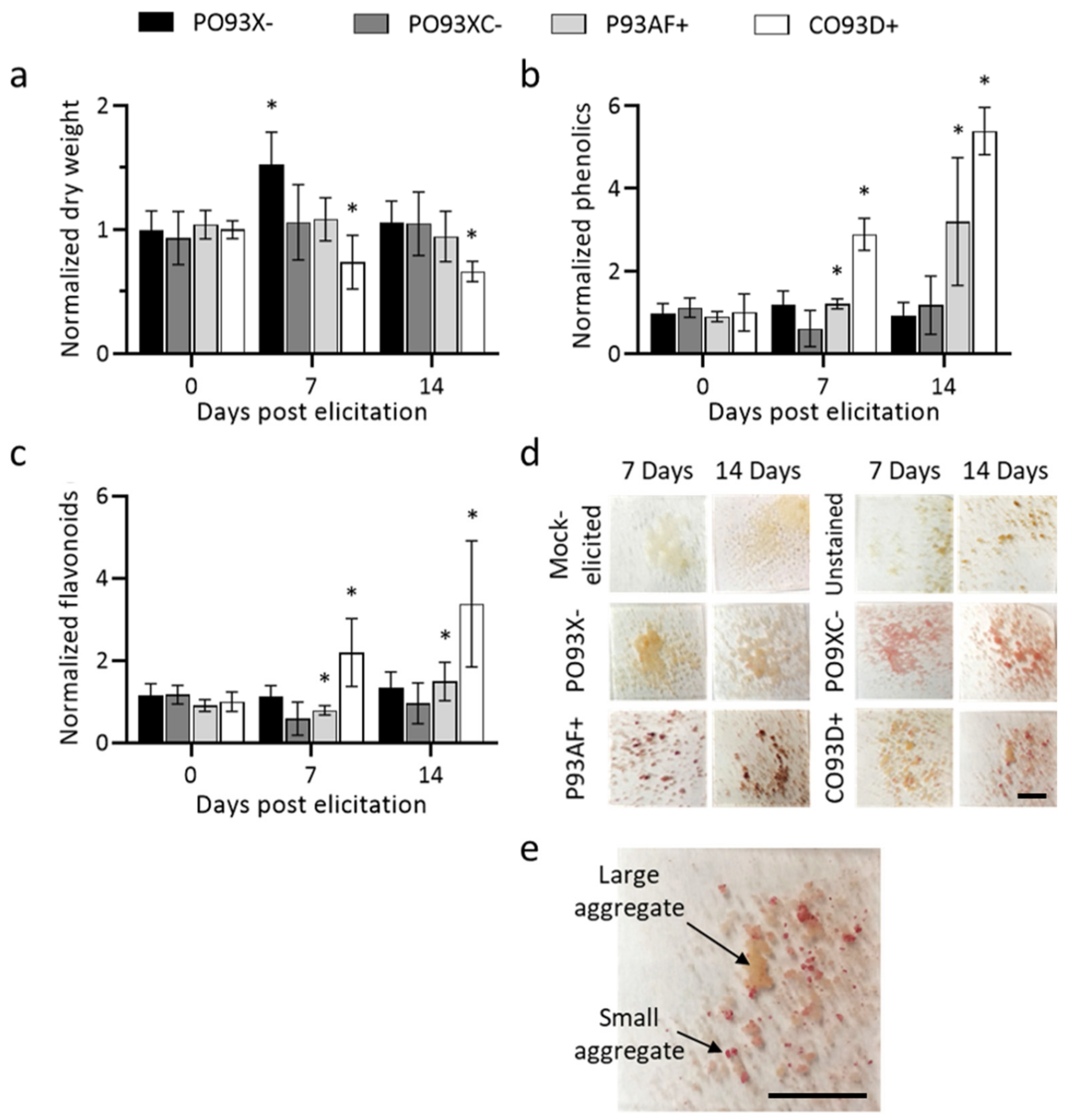
2.1. Methyl Jasmonate Treatment of Non-Paclitaxel and Paclitaxel Accumulating Cultures
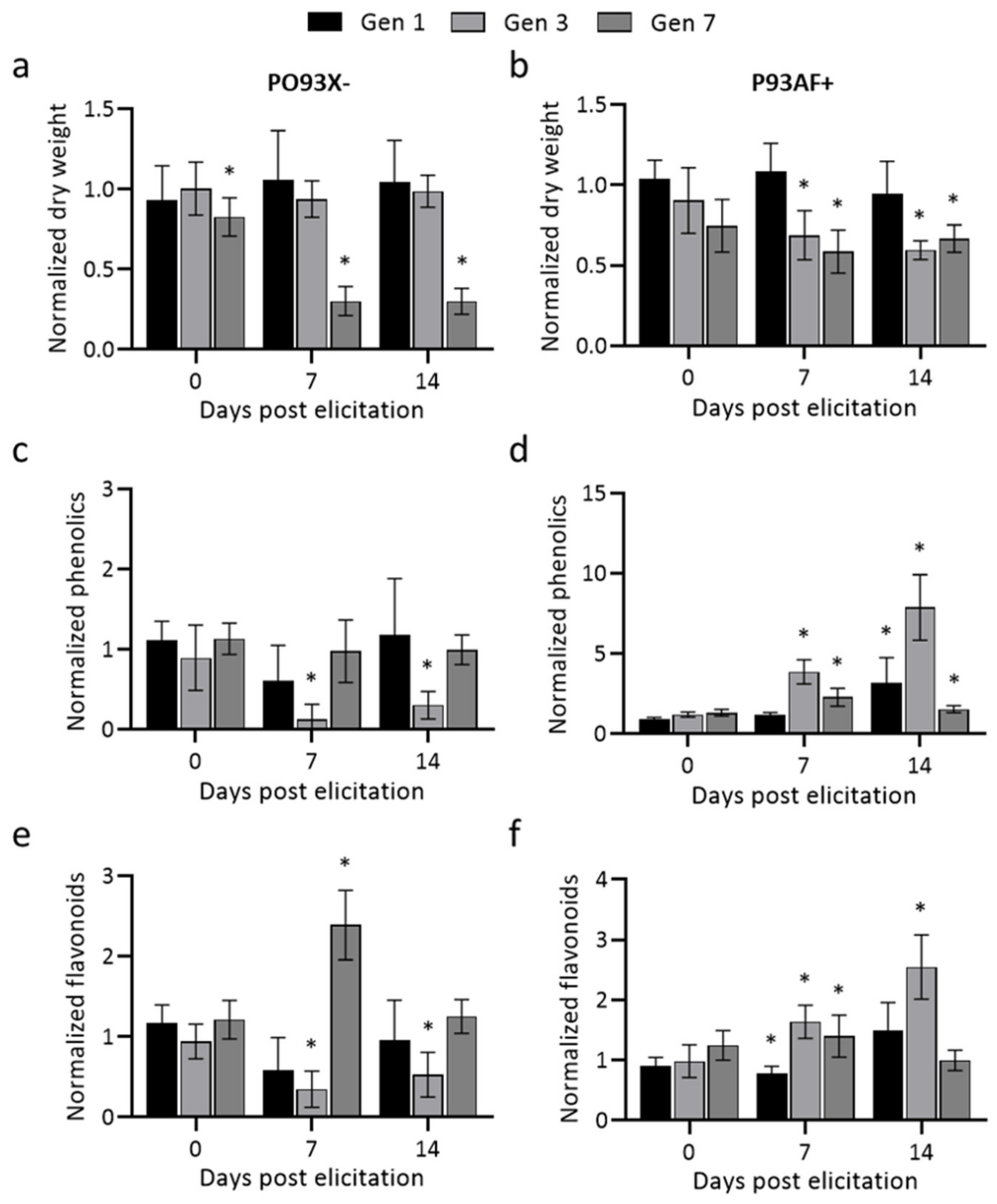
2.2. The Effect of Methyl Jasmonate Treatment in Different Generations
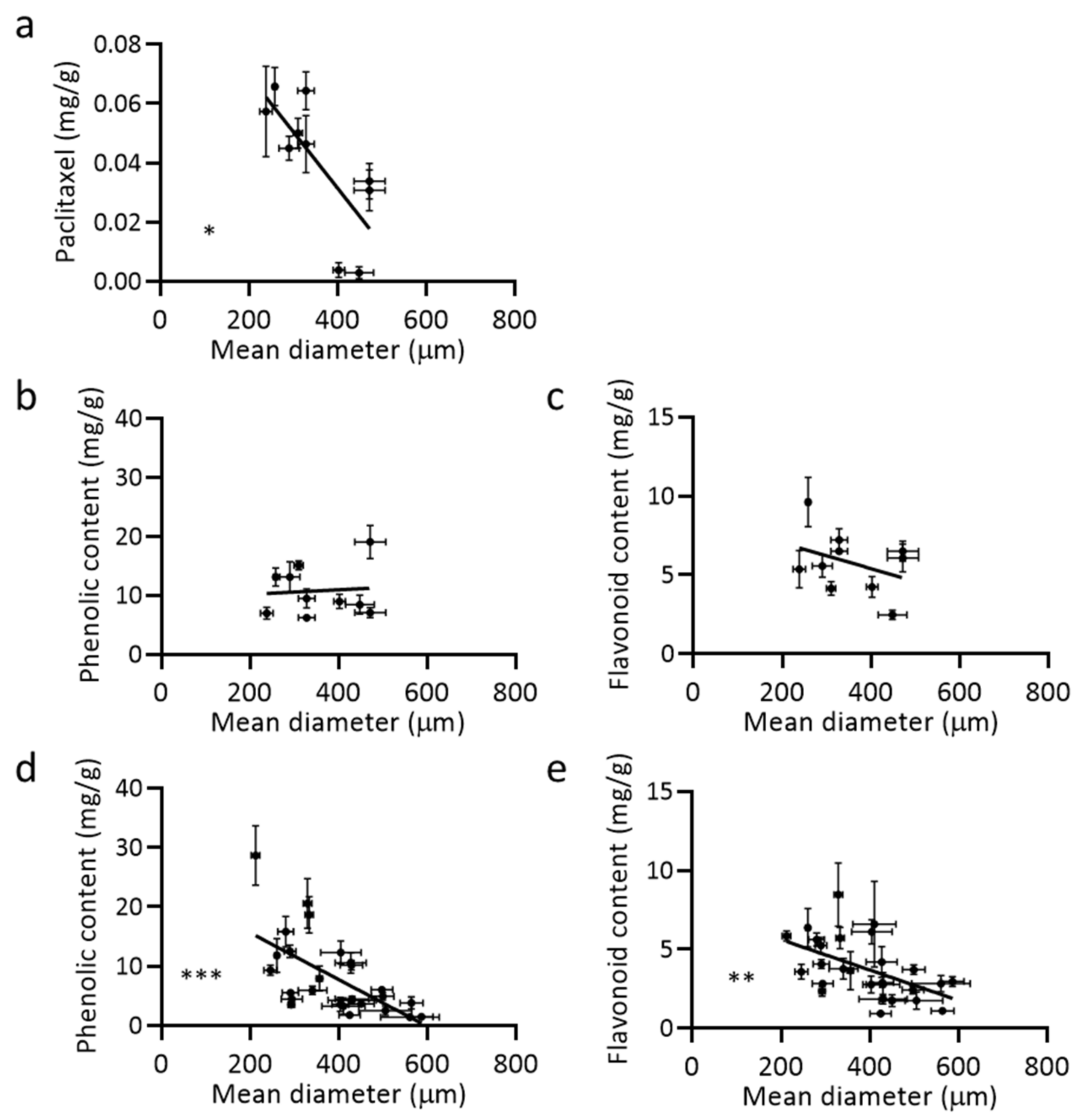
2.3. The Relationship between Metabolite Accumulation and Aggregate Size
3. Materials and Methods
3.1. Maintenance and Treatment of Taxus Cultures
3.2. Long-Term Experimental Set-Up
3.3. Biomass Measurements Using the Coulter Counter
3.4. Phenolic and Flavonoid Content Assays
3.5. Paclitaxel and Baccatin III Quantification
3.6. Lignin Staining
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Weng, J.-K.; Philippe, R.N.; Noel, J.P. The rise of chemodiversity in plants. Science 2012, 336, 1667–1670. [Google Scholar] [CrossRef] [Green Version]
- Matsuura, H.N.; Malik, S.; de Costa, F.; Yousefzadi, M.; Mirjalili, M.H.; Arroo, R.; Bhambra, A.S.; Strnad, M.; Bonfill, M.; Fett-Neto, A.G. Specialized plant metabolism characteristics and impact on target molecule biotechnological production. Mol. Biotechnol. 2018, 60, 169–183. [Google Scholar] [CrossRef]
- Wilson, S.A.; Roberts, S.C. Recent advances towards development and commercialization of plant cell culture processes for the synthesis of biomolecules. Plant Biotechnol. J. 2012, 10, 249–268. [Google Scholar] [CrossRef] [PubMed]
- Weaver, B.A. How Taxol/paclitaxel kills cancer cells. Mol. Biol. Cell 2014, 25, 2677–2681. [Google Scholar] [CrossRef]
- Roberts, S.C. Production and engineering of terpenoids in plant cell culture. Nat. Chem. Biol. 2007, 3, 387. [Google Scholar] [CrossRef] [PubMed]
- Sanchez-Muñoz, R.; Perez-Mata, E.; Almagro, L.; Cusido, R.M.; Bonfill, M.; Palazon, J.; Moyano, E. A Novel Hydroxylation Step in the Taxane Biosynthetic Pathway: A New Approach to Paclitaxel Production by Synthetic Biology. Front. Bioeng. Biotechnol. 2020, 8, 410. [Google Scholar] [CrossRef] [PubMed]
- Jalalpour, Z.; Shabani, L.; Afghani, L.; Sharifi-Tehrani, M.; AMINI, S.A. Stimulatory effect of methyl jasmonate and squalestatin on phenolic metabolism through induction of LOX activity in cell suspension culture of yew. Turk. J. Biol. 2014, 38, 76–82. [Google Scholar] [CrossRef]
- Yang, S.J.; Fang, J.M.; Cheng, Y.S. Lignans, flavonoids and phenolic derivatives from Taxus mairei. J. Chin. Chem. Soc. 1999, 46, 811–818. [Google Scholar] [CrossRef]
- Mertoğluelmas, G. Chemical components of heartwood and sapwood of common Yew (Taxus baccata L.). J. Environ. Biol. 2003, 24, 417–420. [Google Scholar]
- Wickremesinhe, E.R.; Arteea, R.N. Taxus callus cultures: Initiation, growth optimization, characterization and taxol production. Plant Cell Tissue Organ Cult. 1993, 35, 181–193. [Google Scholar] [CrossRef]
- Ketchum, R.; Gibson, D.; Gallo, L.G. Media optimization for maximum biomass production in cell cultures of pacific yew. Plant Cell Tissue Organ Cult. 1995, 42, 185–193. [Google Scholar] [CrossRef]
- Kolewe, M.E.; Henson, M.A.; Roberts, S.C. Analysis of aggregate size as a process variable affecting paclitaxel accumulation in Taxus suspension cultures. Biotechnol. Prog. 2011, 27, 1365–1372. [Google Scholar] [CrossRef] [PubMed]
- Patil, R.A.; Kolewe, M.E.; Roberts, S.C. Cellular aggregation is a key parameter associated with long term variability in paclitaxel accumulation in Taxus suspension cultures. Plant Cell Tissue Organ Cult. 2013, 112, 303–310. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramirez-Estrada, K.; Vidal-Limon, H.; Hidalgo, D.; Moyano, E.; Golenioswki, M.; Cusidó, R.M.; Palazon, J. Elicitation, an effective strategy for the biotechnological production of bioactive high-added value compounds in plant cell factories. Molecules 2016, 21, 182. [Google Scholar] [CrossRef] [PubMed]
- Yukimune, Y.; Tabata, H.; Higashi, Y.; Hara, Y. Methyl jasmonate-induced overproduction of paclitaxel and baccatin III in Taxus cell suspension cultures. Nat. Biotechnol. 1996, 14, 1129–1132. [Google Scholar] [CrossRef] [PubMed]
- Li, S.-T.; Zhang, P.; Zhang, M.; Fu, C.-H.; Zhao, C.-F.; Dong, Y.-S.; Guo, A.-Y.; Yu, L.-J. Transcriptional profile of Taxus chinensis cells in response to methyl jasmonate. BMC Genom. 2012, 13, 295. [Google Scholar] [CrossRef] [Green Version]
- Patil, R.A.; Kolewe, M.E.; Normanly, J.; Walker, E.L.; Roberts, S.C. Contribution of taxane biosynthetic pathway gene expression to observed variability in paclitaxel accumulation in Taxus suspension cultures. Biotechnol. J. 2012, 7, 418–427. [Google Scholar] [CrossRef]
- Zhou, T.; Luo, X.; Yu, C.; Zhang, C.; Zhang, L.; Song, Y.-b.; Dong, M.; Shen, C. Transcriptome analyses provide insights into the expression pattern and sequence similarity of several taxol biosynthesis-related genes in three Taxus species. BMC Plant Biol. 2019, 19, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Bulgakov, V.P.; Tchernoded, G.; Veselova, M.; Fedoreyev, S.; Muzarok, T.; Zhuravlev, Y. Catechin production in cultured cells of Taxus cuspidata and Taxus baccata. Biotechnol. Lett. 2011, 33, 1879–1883. [Google Scholar] [CrossRef]
- Patil, R.A.; Lenka, S.K.; Normanly, J.; Walker, E.L.; Roberts, S.C. Methyl jasmonate represses growth and affects cell cycle progression in cultured Taxus cells. Plant Cell Rep. 2014, 33, 1479–1492. [Google Scholar] [CrossRef] [Green Version]
- Furmanowa, M.; Glowniak, K.; Syklowska-Baranek, K.; Zgórka, G.; Józefczyk, A. Effect of picloram and methyl jasmonate on growth and taxane accumulation in callus culture of Taxus× media var. Hatfieldii. Plant Cell Tissue Organ Cult. 1997, 49, 75–79. [Google Scholar] [CrossRef]
- Kim, J.I.; Zhang, X.; Pascuzzi, P.E.; Liu, C.J.; Chapple, C. Glucosinolate and phenylpropanoid biosynthesis are linked by proteasome-dependent degradation of PAL. New Phytol. 2020, 225, 154–168. [Google Scholar] [CrossRef] [Green Version]
- Kim, B.J.; Gibson, D.M.; Shuler, M.L. Effect of subculture and elicitation on instability of taxol production in Taxus sp. suspension cultures. Biotechnol. Prog. 2004, 20, 1666–1673. [Google Scholar] [CrossRef]
- Fu, C.; Li, L.; Wu, W.; Li, M.; Yu, X.; Yu, L. Assessment of genetic and epigenetic variation during long-term Taxus cell culture. Plant Cell Rep. 2012, 31, 1321–1331. [Google Scholar] [CrossRef]
- Sanchez-Muñoz, R.; Moyano, E.; Khojasteh, A.; Bonfill, M.; Cusido, R.M.; Palazon, J. Genomic methylation in plant cell cultures: A barrier to the development of commercial long-term biofactories. Eng. Life Sci. 2019, 19, 872–879. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pesquet, E.; Wagner, A.; Grabber, J.H. Cell culture systems: Invaluable tools to investigate lignin formation and cell wall properties. Curr. Opin. Biotechnol. 2019, 56, 215–222. [Google Scholar] [CrossRef] [PubMed]
- Chang, X.F.; Chandra, R.; Berleth, T.; Beatson, R.P. Rapid, microscale, acetyl bromide-based method for high-throughput determination of lignin content in Arabidopsis thaliana. J. Agric. Food Chem. 2008, 56, 6825–6834. [Google Scholar] [CrossRef] [PubMed]
- Gao, Z.; Gao, S.; Li, P.; Zhang, Y.; Ma, B.; Wang, Y. Exogenous methyl jasmonate promotes salt stress–induced growth inhibition and prioritizes defense response of Nitraria tangutorum Bobr. Physiol. Plant. 2021, 172, 162–175. [Google Scholar] [CrossRef] [PubMed]
- Benevenuto, R.F.; Seldal, T.; Hegland, S.J.; Rodriguez-Saona, C.; Kawash, J.; Polashock, J. Transcriptional profiling of methyl jasmonate-induced defense responses in bilberry (Vaccinium myrtillus L.). BMC Plant Biol. 2019, 19, 1–18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilson, S.A.; Maindarkar, S.N.; McKee, M.C.; Vilkhovoy, M.; Henson, M.A.; Roberts, S.C. A population balance model to modulate shear for the control of aggregation in Taxus suspension cultures. Biotechnol. Prog. 2020, 36, e2932. [Google Scholar] [CrossRef] [PubMed]
- Kolewe, M.E.; Henson, M.A.; Roberts, S.C. Characterization of aggregate size in Taxus suspension cell culture. Plant Cell Rep. 2010, 29, 485–494. [Google Scholar] [CrossRef] [Green Version]
- Ainsworth, E.A.; Gillespie, K.M. Estimation of total phenolic content and other oxidation substrates in plant tissues using Folin–Ciocalteu reagent. Nat. Protoc. 2007, 2, 875–877. [Google Scholar] [CrossRef] [PubMed]
- Chang, C.-C.; Yang, M.-H.; Wen, H.-M.; Chern, J.-C. Estimation of total flavonoid content in propolis by two complementary colorimetric methods. J. Food Drug Anal. 2002, 10. [Google Scholar] [CrossRef]
- Naill, M.C.; Roberts, S.C. Preparation of single cells from aggregated Taxus suspension cultures for population analysis. Biotechnol. Bioeng. 2004, 86, 817–826. [Google Scholar] [CrossRef] [PubMed]
- Bucholtz, D.L.; Cantrell, R.P.; Axtell, J.D.; Lechtenberg, V.L. Lignin biochemistry of normal and brown midrib mutant sorghum. J. Agric. Food Chem. 1980, 28, 1239–1241. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Generation | 7 Days PE (mg/g) | 14 Days PE (mg/g) |
|---|---|---|
| 1 | 0.05 ± 0.005 | 0.05 ± 0.004 |
| 3 | 0.03 ± 0.007 | 0.06 ± 0.006 |
| 5 | 0.03 ± 0.006 | 0.05 ± 0.009 |
| 7 | 0.002 ± 0.002 | 0.003 ± 0.002 |
| 9 | 0.07 ± 0.006 | 0.06 ± 0.02 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
McKee, M.C.; Wilson, S.A.; Roberts, S.C. The Interface amongst Conserved and Specialized Pathways in Non-Paclitaxel and Paclitaxel Accumulating Taxus Cultures. Metabolites 2021, 11, 688. https://doi.org/10.3390/metabo11100688
McKee MC, Wilson SA, Roberts SC. The Interface amongst Conserved and Specialized Pathways in Non-Paclitaxel and Paclitaxel Accumulating Taxus Cultures. Metabolites. 2021; 11(10):688. https://doi.org/10.3390/metabo11100688
Chicago/Turabian StyleMcKee, Michelle C., Sarah A. Wilson, and Susan C. Roberts. 2021. "The Interface amongst Conserved and Specialized Pathways in Non-Paclitaxel and Paclitaxel Accumulating Taxus Cultures" Metabolites 11, no. 10: 688. https://doi.org/10.3390/metabo11100688
APA StyleMcKee, M. C., Wilson, S. A., & Roberts, S. C. (2021). The Interface amongst Conserved and Specialized Pathways in Non-Paclitaxel and Paclitaxel Accumulating Taxus Cultures. Metabolites, 11(10), 688. https://doi.org/10.3390/metabo11100688