
Concentrations of Fecal Bile Acids in Participants with Functional Gut Disorders and Healthy Controls

 ,
,  ,
,  ,
, 
Abstract
:1. Introduction
2. Results
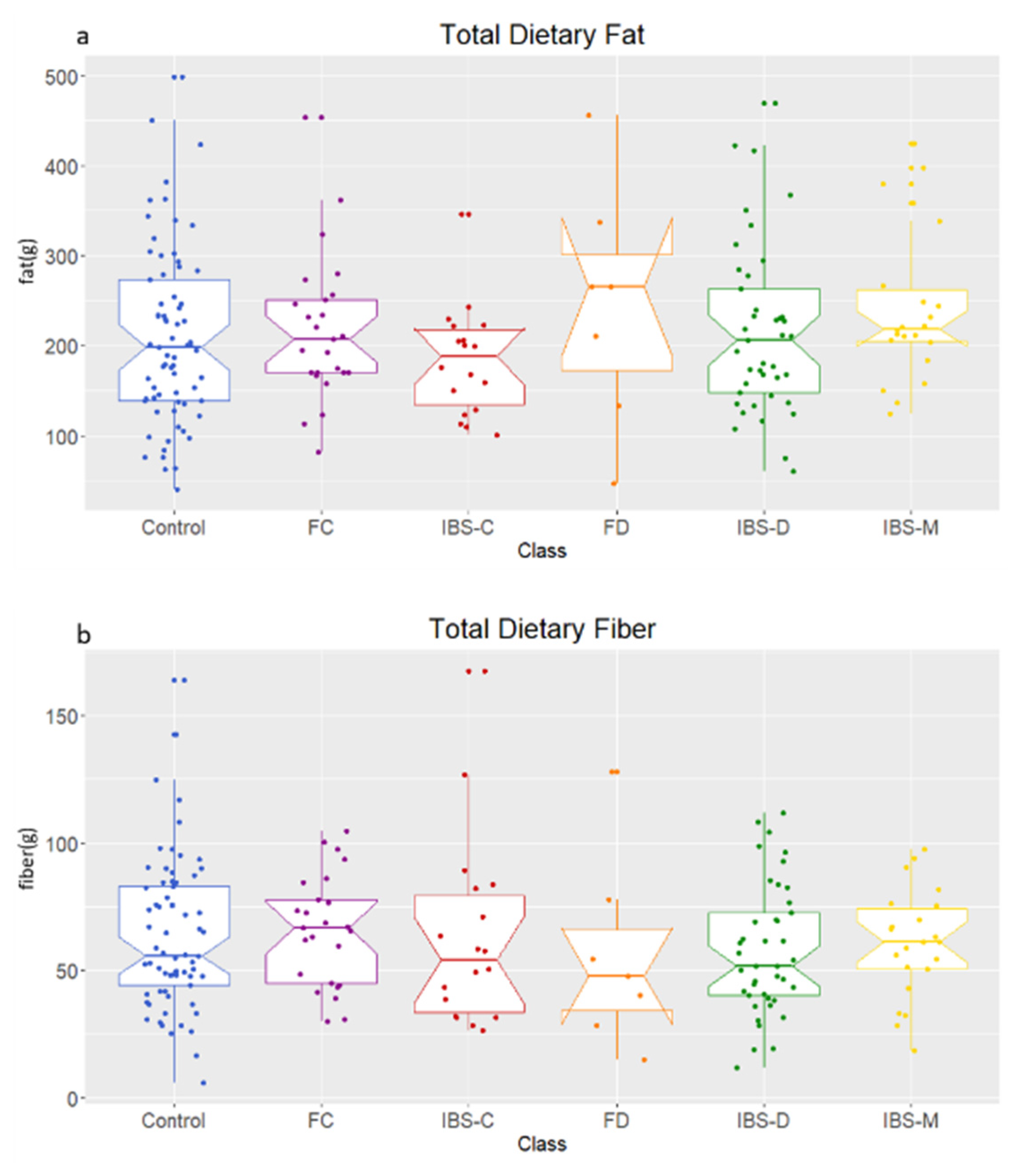
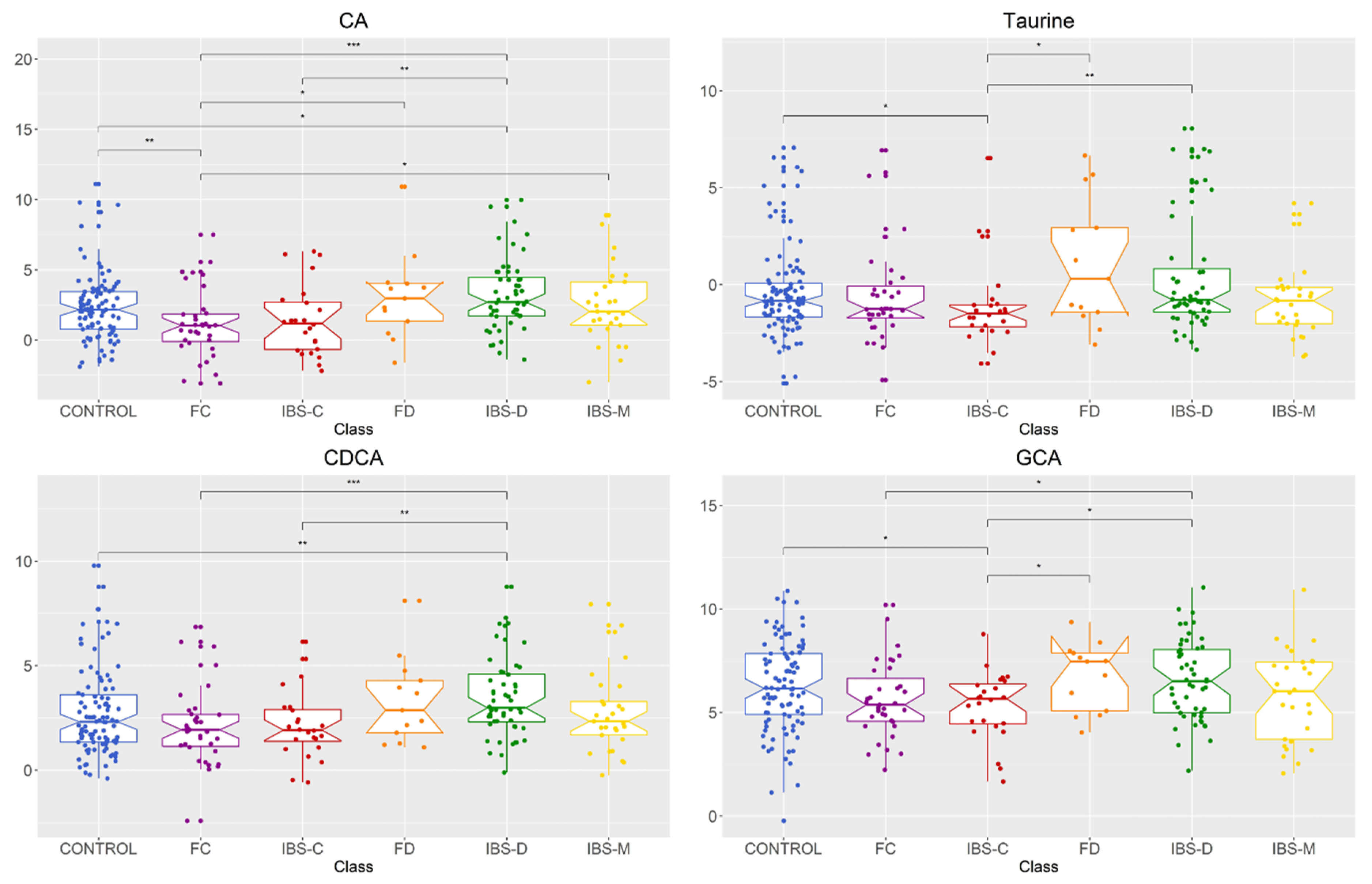
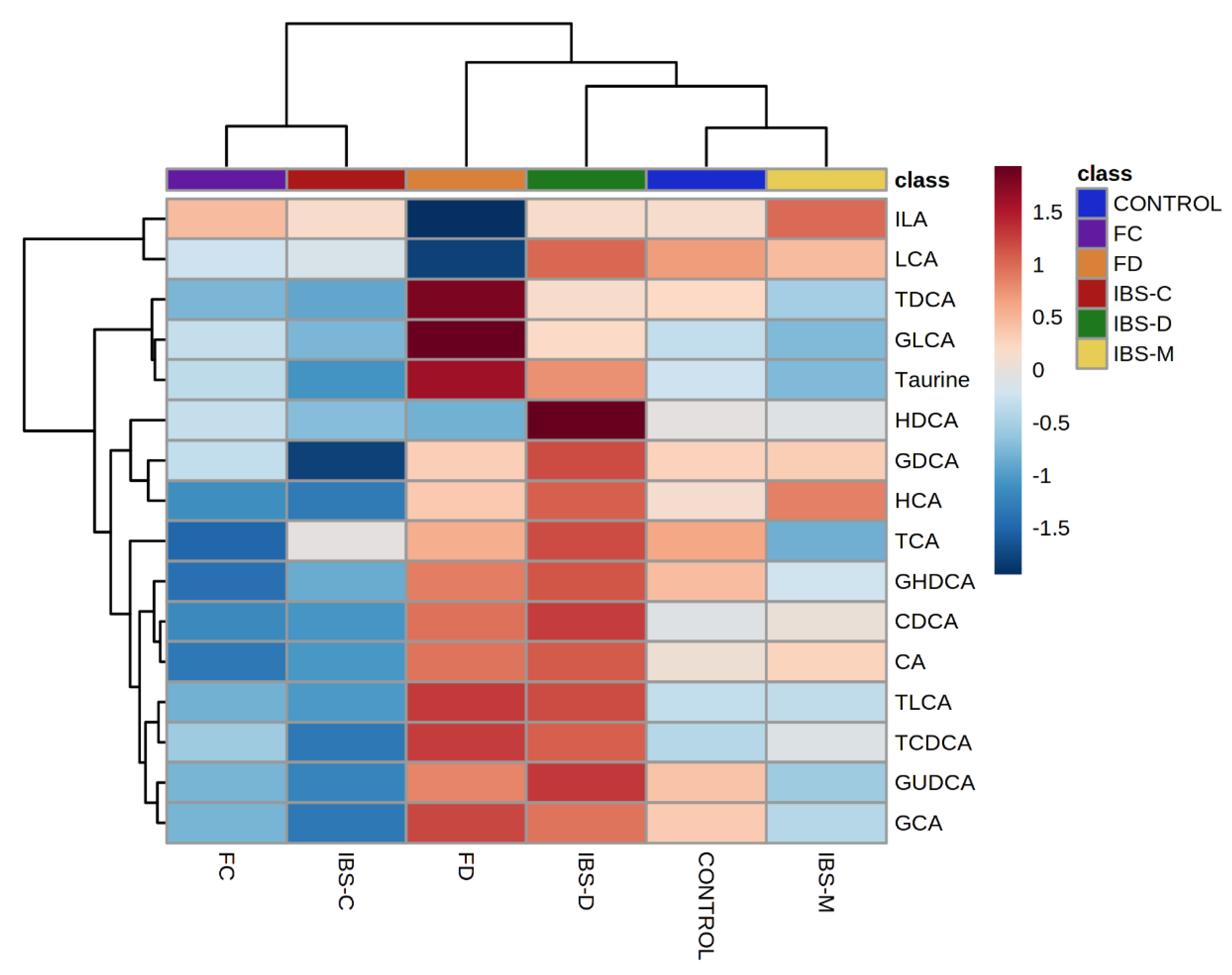
2.1. Comparison of Fecal Bile Acid Concentrations between Healthy Control, IBS Subtypes, FC, and FD Groups
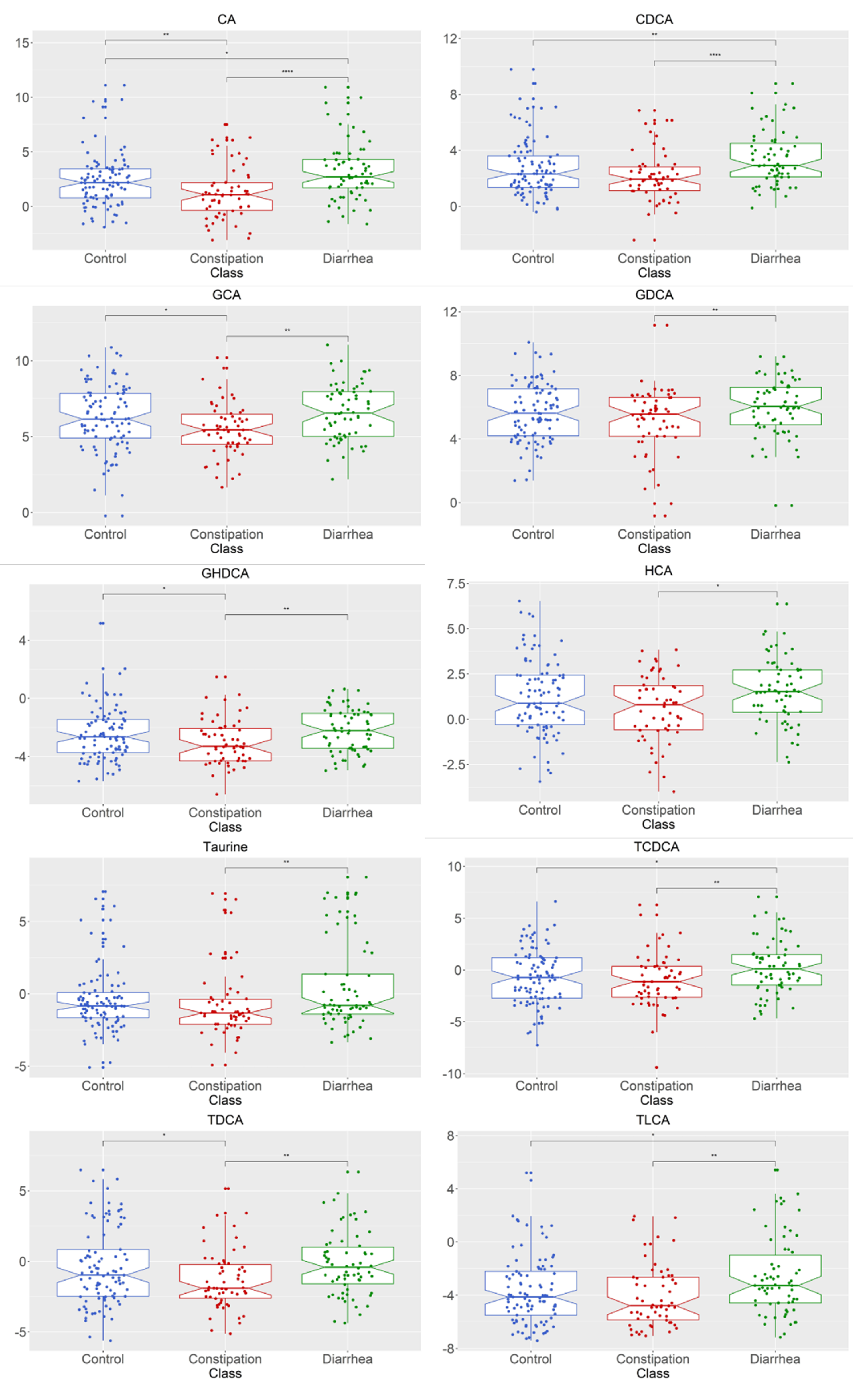
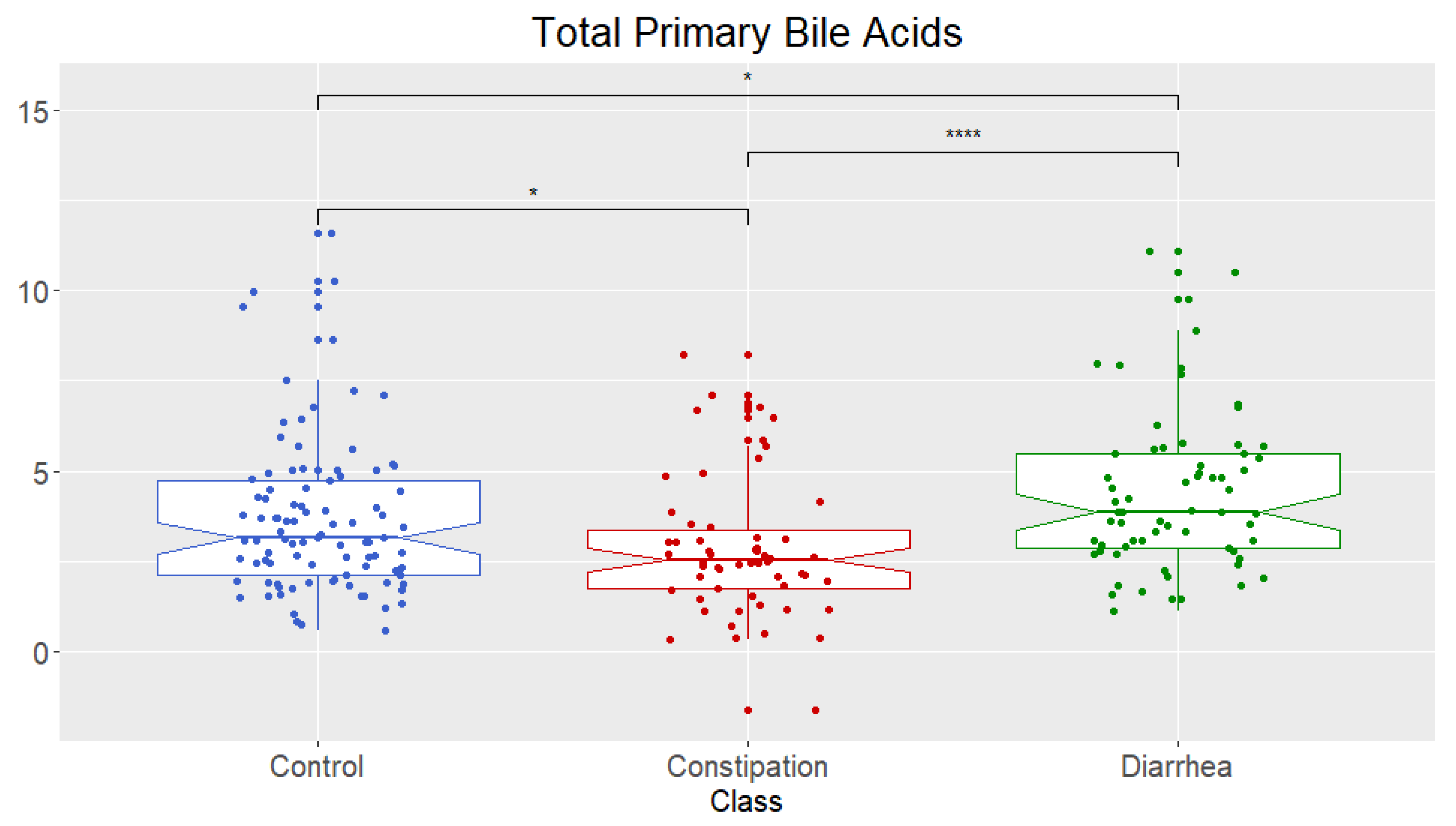
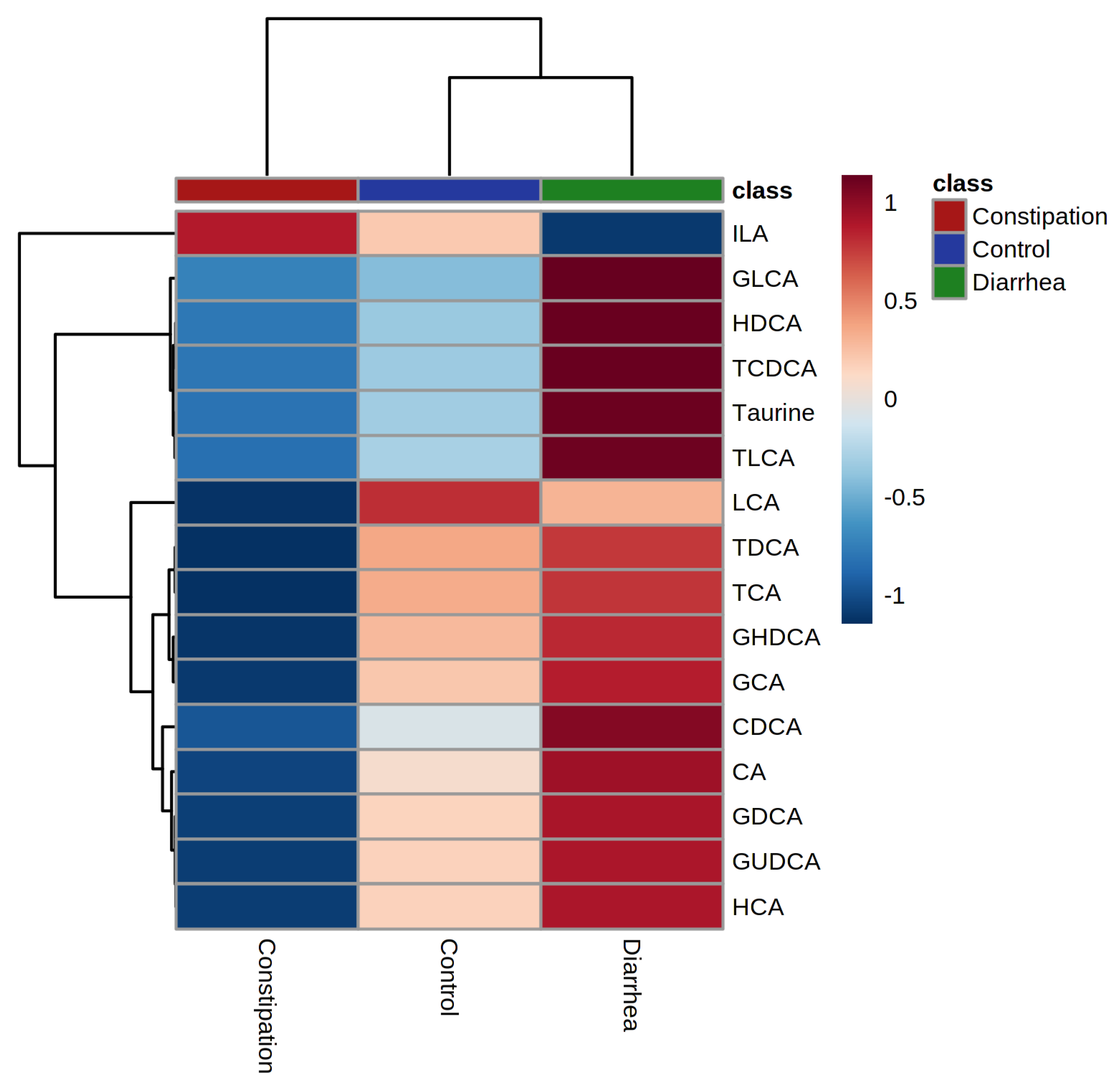
2.2. Bile Acid Concentrations Compared between Healthy Control and Combined Functional Groups
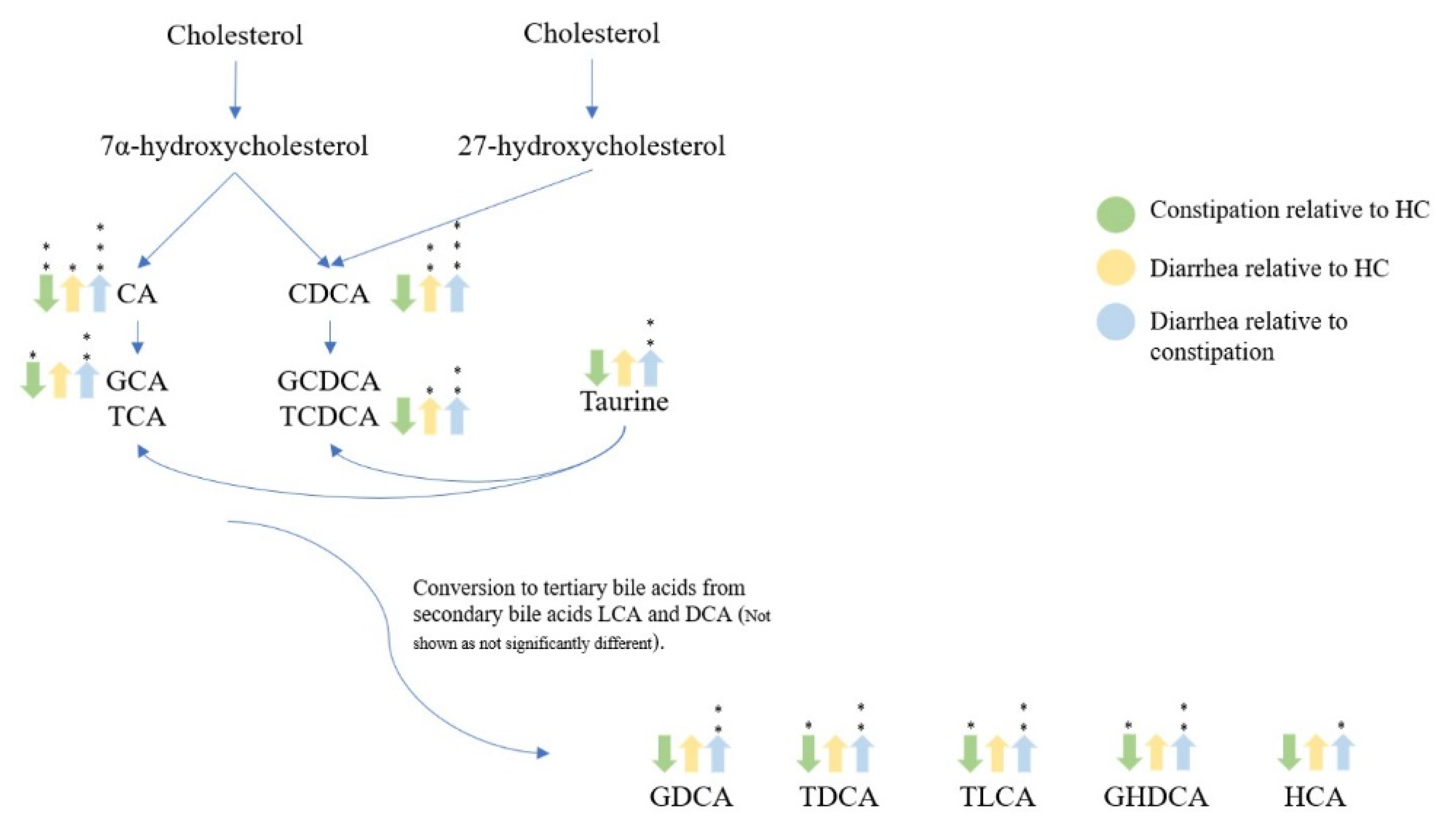
3. Discussion
4. Materials and Methods
4.1. Participants
4.2. Diet Record and Sample Collection
4.3. Standards and Reagents
4.4. Sample Extraction
4.5. Statistical Analyses
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group p-Value | Gender p-Value | Group × Gender p-Value | |
|---|---|---|---|
| CA | 0.0002 *** | 0.168 | 0.092 |
| CDCA | 0.001 *** | 0.034 | 0.152 |
| GHDCA | 0.015 * | 0.646 | 0.214 |
| GUDCA | 0.138 | 0.205 | 0.848 |
| GDCA | 0.030 * | 0.286 | 0.016 * |
| HDCA | 0.160 | 0.045 | 0.003 * |
| HCA | 0.025 * | 0.277 | 0.140 |
| GCA | 0.006 ** | 0.488 | 0.613 |
| ILA | 0.893 | 0.821 | 0.632 |
| GLCA | 0.295 | 0.765 | 0.250 |
| Taurine | 0.018 * | 0.402 | 0.768 |
| LCA | 0.581 | 0.539 | 0.316 |
| TLCA | 0.007 ** | 0.348 | 0.653 |
| TCDCA | 0.021 * | 0.320 | 0.456 |
| TDCA | 0.018 * | 0.445 | 0.083 |
| TCA | 0.098 | 0.157 | 0.134 |
| Control | IBS-C | FC | IBS-D | FD | IBS-M | |
|---|---|---|---|---|---|---|
| GHDCA | 0.72 ± 3.61 | 0.29 ± 0.56 | 0.21 ± 0.26 | 0.34 ± 0.31 | 0.40 ± 0.49 | 0.24 ± 0.24 |
| GUDCA | 0.81 ± 3.88 | 0.32 ± 0.57 | 0.47 ± 1.26 | 0.40 ± 0.38 | 0.47 ± 0.58 | 0.28 ± 0.25 |
| GDCA | 110.41 ± 167.88 | 53.49 ± 54.29 | 126.42 ± 371.25 | 119.12 ± 130.64 | 91.47 ± 88.84 | 92.10 ± 112.37 |
| HDCA | 90.55 ± 101.77 | 92.99 ± 96.97 | 87.33 ± 77.63 | 122.15 ± 135.87 | 84.13 ± 88.98 | 71.12 ± 71.11 |
| HCA | 6.77 ± 13.63 | 2.26 ± 1.97 | 3.43 ± 3.87 | 5.62 ± 6.66 | 9.87 ± 21.42 | 5.21 ± 7.40 |
| GCA | 199.35 ± 317.56 | 68.1 ± 86.16 | 118.23 ± 222.78 | 221.83 ± 346.59 | 177.95 ± 169.83 | 166.30 ± 353.11 |
| ILA | 31.51 ± 20.67 | 30.91 ± 18.1 | 31.25 ± 17.92 | 33.17 ± 23.27 | 28.97 ± 15.79 | 35.01 ± 22.27 |
| GLCA | 0.18 ± 0.18 | 0.17 ± 0.13 | 0.21 ± 0.21 | 0.20 ± 0.15 | 2.01 ± 6.22 | 0.16 ± 0.11 |
| Taurine | 5.56 ± 18.91 | 4.65 ± 18.29 | 7.29 ± 23.0 | 17.85 ± 46.93 | 16.59 ± 29.4 | 1.84 ± 4.07 |
| LCA | 548.75 ± 336.88 | 483.52 ± 270.23 | 481.59 ± 271.46 | 618.36 ± 426.56 | 451.99 ± 189.5 | 513.83 ± 289.08 |
| CDCA | 29.65 ± 102.48 | 9.31 ± 15.35 | 11.61 ± 23.37 | 31.92 ± 67.42 | 32.39 ± 70.86 | 22.83 ± 50.26 |
| CA | 56.16 ± 255.46 | 12.44 ± 23.53 | 11.16 ± 30.87 | 58.79 ± 172.97 | 160.13 ± 513.21 | 37.52 ± 99.60 |
| TLCA | 0.94 ± 4.46 | 0.26 ± 0.77 | 0.35 ± 0.8 | 1.69 ± 6.10 | 2.17 ± 3.82 | 0.76 ± 3.06 |
| TCDCA | 3.35 ± 10.5 | 1.19 ± 2.48 | 4.65 ± 14.37 | 6.35 ± 19.67 | 7.28 ± 12.12 | 3.30 ± 8.97 |
| TDCA | 4.84 ± 12.5 | 0.64 ± 0.68 | 2.27 ± 6.24 | 3.54 ± 11.74 | 5.34 ± 5.89 | 3.26 ± 10.22 |
| TCA | 4.14 ± 7.82 | 2.06 ± 3.1 | 2.10 ± 4.87 | 6.47 ± 20.76 | 2.70 ± 4.0 | 6.55 ± 28.14 |
References
- Zhou, H.; Hylemon, P.B. Bile acids are nutrient signaling hormones. Steroids 2014, 86, 62–68. [Google Scholar] [CrossRef] [Green Version]
- Joyce, S.A.; Gahan, C.G.M. Bile Acid Modifications at the Microbe-Host Interface: Potential for Nutraceutical and Pharmaceutical Interventions in Host Health. Annu. Rev. Food Sci. Technol. 2016, 7, 313–333. [Google Scholar] [CrossRef] [PubMed]
- Molinaro, A.; Wahlstrom, A.; Marschall, H.U. Role of Bile Acids in Metabolic Control. Trends Endocrinol. Metab. 2018, 29, 31–41. [Google Scholar] [CrossRef]
- Oduyebo, I.; Camilleri, M. Bile acid disease: The emerging epidemic. Curr. Opin. Gastroenterol. 2017, 33, 189–195. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Molinero, N.; Ruiz, L.; Sanchez, B.; Margolles, A.; Delgado, S. Intestinal Bacteria Interplay with Bile and Cholesterol Metabolism: Implications on Host Physiology. Front. Physiol. 2019, 10, 185. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wahlstrom, A.; Sayin, S.I.; Marschall, H.U.; Backhed, F. Intestinal Crosstalk between Bile Acids and Microbiota and Its Impact on Host Metabolism. Cell Metab. 2016, 24, 41–50. [Google Scholar] [CrossRef] [Green Version]
- Joyce, S.A.; MacSharry, J.; Casey, P.G.; Kinsella, M.; Murphy, E.F.; Shanahan, F.; Hill, C.; Gahan, C.G. Regulation of host weight gain and lipid metabolism by bacterial bile acid modification in the gut. Proc. Natl. Acad. Sci. USA 2014, 111, 7421–7426. [Google Scholar] [CrossRef] [Green Version]
- Vijayvargiya, P.; Busciglio, I.; Burton, D.; Donato, L.; Lueke, A.; Camilleri, M. Bile Acid Deficiency in a Subgroup of Patients with Irritable Bowel Syndrome with Constipation Based on Biomarkers in Serum and Fecal Samples. Clin. Gastroenterol. Hepatol. 2018, 16, 522–527. [Google Scholar] [CrossRef]
- Camilleri, M. Bile Acid diarrhea: Prevalence, pathogenesis, and therapy. Gut Liver 2015, 9, 332–339. [Google Scholar] [CrossRef]
- Wong, B.S.; Camilleri, M.; Carlson, P.; McKinzie, S.; Busciglio, I.; Bondar, O.; Dyer, R.B.; Lamsam, J.; Zinsmeister, A.R. Increased bile acid biosynthesis is associated with irritable bowel syndrome with diarrhea. Clin. Gastroenterol. Hepatol. 2012, 10, 1009–1015.e1003. [Google Scholar] [CrossRef] [Green Version]
- Shin, A.; Camilleri, M.; Vijayvargiya, P.; Busciglio, I.; Burton, D.; Ryks, M.; Rhoten, D.; Lueke, A.; Saenger, A.; Girtman, A. Bowel functions, fecal unconjugated primary and secondary bile acids, and colonic transit in patients with irritable bowel syndrome. Clin. Gastroenterol. Hepatol. 2013, 11, 1270–1275.e1271. [Google Scholar] [CrossRef] [Green Version]
- Duboc, H.; Rainteau, D.; Rajca, S.; Humbert, L.; Farabos, D.; Maubert, M.; Grondin, V.; Jouet, P.; Bouhassira, D.; Seksik, P. Increase in fecal primary bile acids and dysbiosis in patients with diarrhea-predominant irritable bowel syndrome. Neurogastroenterol. Motil. 2012, 24, 513. [Google Scholar] [CrossRef] [PubMed]
- Dior, M.; Delagrèverie, H.; Duboc, H.; Jouet, P.; Coffin, B.; Brot, L.; Humbert, L.; Trugnan, G.; Seksik, P.; Sokol, H.; et al. Interplay between bile acid metabolism and microbiota in irritable bowel syndrome. Neurogastroenterol. Motil. 2016, 28, 1330–1340. [Google Scholar] [CrossRef] [PubMed]
- Jeffery, I.B.; Das, A.; O’Herlihy, E.; Coughlan, S.; Cisek, K.; Moore, M.; Bradley, F.; Carty, T.; Pradhan, M.; Dwibedi, C.; et al. Differences in Fecal Microbiomes and Metabolomes of People With vs Without Irritable Bowel Syndrome and Bile Acid Malabsorption. Gastroenterology 2020, 158, 1016–1028.e1018. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Yang, W.; Chen, Y.; Huang, F.; Lu, L.; Lin, C.; Huang, T.; Ning, Z.; Zhai, L.; Zhong, L.L. A Clostridia-rich microbiota enhances bile acid excretion in diarrhea-predominant irritable bowel syndrome. J. Clin. Investig. 2019, 130, 438–450. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rao, A.S.; Wong, B.S.; Camilleri, M.; Odunsi-Shiyanbade, S.T.; McKinzie, S.; Ryks, M.; Burton, D.; Carlson, P.; Lamsam, J.; Singh, R.; et al. Chenodeoxycholate in females with irritable bowel syndrome-constipation: A pharmacodynamic and pharmacogenetic analysis. Gastroenterology 2010, 139, 1549–1558.e1541. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walters, J.R.; Pattni, S.S. Managing bile acid diarrhoea. Ther. Adv. Gastroenterol. 2010, 3, 349–357. [Google Scholar] [CrossRef] [Green Version]
- Pimentel, M.; Talley, N.J.; Quigley, E.M.; Hani, A.; Sharara, A.; Mahachai, V. Report from the multinational irritable bowel syndrome initiative 2012. Gastroenterology 2013, 144, e1–e5. [Google Scholar] [CrossRef]
- Heenan, P.; Creemers, R.H.; Sharma, S.; Keenan, J.; Bayer, S.; Young, W.; Cooney, J.; Armstrong, K.; Fraser, K.; Skidmore, P.M.; et al. Cohort Profile: The Christchurch IBS cOhort to investigate Mechanisms for gut Relief and improved Transit (COMFORT). Inflamm. Intest. Dis. 2020, 5, 132–143. [Google Scholar] [CrossRef]
- Chong, J.; Soufan, O.; Li, C.; Caraus, I.; Li, S.; Bourque, G.; Wishart, D.S.; Xia, J. MetaboAnalyst 4.0: Towards more transparent and integrative metabolomics analysis. Nucleic Acids Res. 2018, 46, W486–W494. [Google Scholar] [CrossRef] [Green Version]
- Chong, J.; Wishart, D.S.; Xia, J. Using MetaboAnalyst 4.0 for Comprehensive and Integrative Metabolomics Data Analysis. Curr. Protoc. Bioinform. 2019, 68, e86. [Google Scholar] [CrossRef] [PubMed]







| Name | Acronym |
|---|---|
| Beta-muricholic acid | βMCA |
| Cheno-deoxycholic acid | CDCA |
| Cholic acid | CA |
| Deuterated (d4) cholic acid (IS) | d4-CA |
| Glyco-cheno-deoxycholic acid | GCDCA |
| Glyco-cholic acid | GCA |
| Glyco-deoxycholic acid | GDCA |
| Glyco-hyo-deoxycholic acid | GHDCA |
| Glyco-litho cholic acid | GLCA |
| Glyco-urso-deoxycholic acid | GUDCA |
| Hyo-cholic acid | HCA |
| Hyo-deoxycholic acid | HDCA |
| Iso-lithocholic acid | ILA |
| Litho-cholic acid | LCA |
| Taurine | Taurine |
| Tauro-alpha-muricholic acid | TαMCA |
| Tauro-beta-muricholic acid | TβMCA |
| Tauro-cheno-deoxycholic acid | TCDCA |
| Tauro-cholic acid | TCA |
| Tauro-deoxycholic acid | TDCA |
| Tauro-hyo-deoxycholic acid | THDCA |
| Tauro-litho cholic acid | TLCA |
| Tauro-urso-deoxycholic acid | TUDCA |
| Urso-deoxycholic acid | UDCA |
| Control | IBS-C | FC | IBS-D | FD | IBS-M | p-Value | |
|---|---|---|---|---|---|---|---|
| Female (male) n | 52 (45) | 23 (1) | 25 (10) | 40 (12) | 11 (2) | 24 (5) | 0.0001 |
| Age (mean) | 54.4 | 53.5 | 59.1 | 52.8 | 58.4 | 50.5 | 0.128 |
| Fecal average dry weight (%) | 27.25 | 31.11 | 30.16 | 25.18 | 26.10 | 31.35 | 0.013 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
James, S.C.; Fraser, K.; Young, W.; Heenan, P.E.; Gearry, R.B.; Keenan, J.I.; Talley, N.J.; Joyce, S.A.; McNabb, W.C.; Roy, N.C. Concentrations of Fecal Bile Acids in Participants with Functional Gut Disorders and Healthy Controls. Metabolites 2021, 11, 612. https://doi.org/10.3390/metabo11090612
James SC, Fraser K, Young W, Heenan PE, Gearry RB, Keenan JI, Talley NJ, Joyce SA, McNabb WC, Roy NC. Concentrations of Fecal Bile Acids in Participants with Functional Gut Disorders and Healthy Controls. Metabolites. 2021; 11(9):612. https://doi.org/10.3390/metabo11090612
Chicago/Turabian StyleJames, Shanalee C., Karl Fraser, Wayne Young, Phoebe E. Heenan, Richard B. Gearry, Jacqueline I. Keenan, Nicholas J. Talley, Susan A. Joyce, Warren C. McNabb, and Nicole C. Roy. 2021. "Concentrations of Fecal Bile Acids in Participants with Functional Gut Disorders and Healthy Controls" Metabolites 11, no. 9: 612. https://doi.org/10.3390/metabo11090612
APA StyleJames, S. C., Fraser, K., Young, W., Heenan, P. E., Gearry, R. B., Keenan, J. I., Talley, N. J., Joyce, S. A., McNabb, W. C., & Roy, N. C. (2021). Concentrations of Fecal Bile Acids in Participants with Functional Gut Disorders and Healthy Controls. Metabolites, 11(9), 612. https://doi.org/10.3390/metabo11090612