

Effects of Proteases from Pineapple and Papaya on Protein Digestive Capacity and Gut Microbiota in Healthy C57BL/6 Mice and Dose-Manner Response on Mucosal Permeability in Human Reconstructed Intestinal 3D Tissue Model
 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Animal Experiment Design
2.3. Animal Material Collection
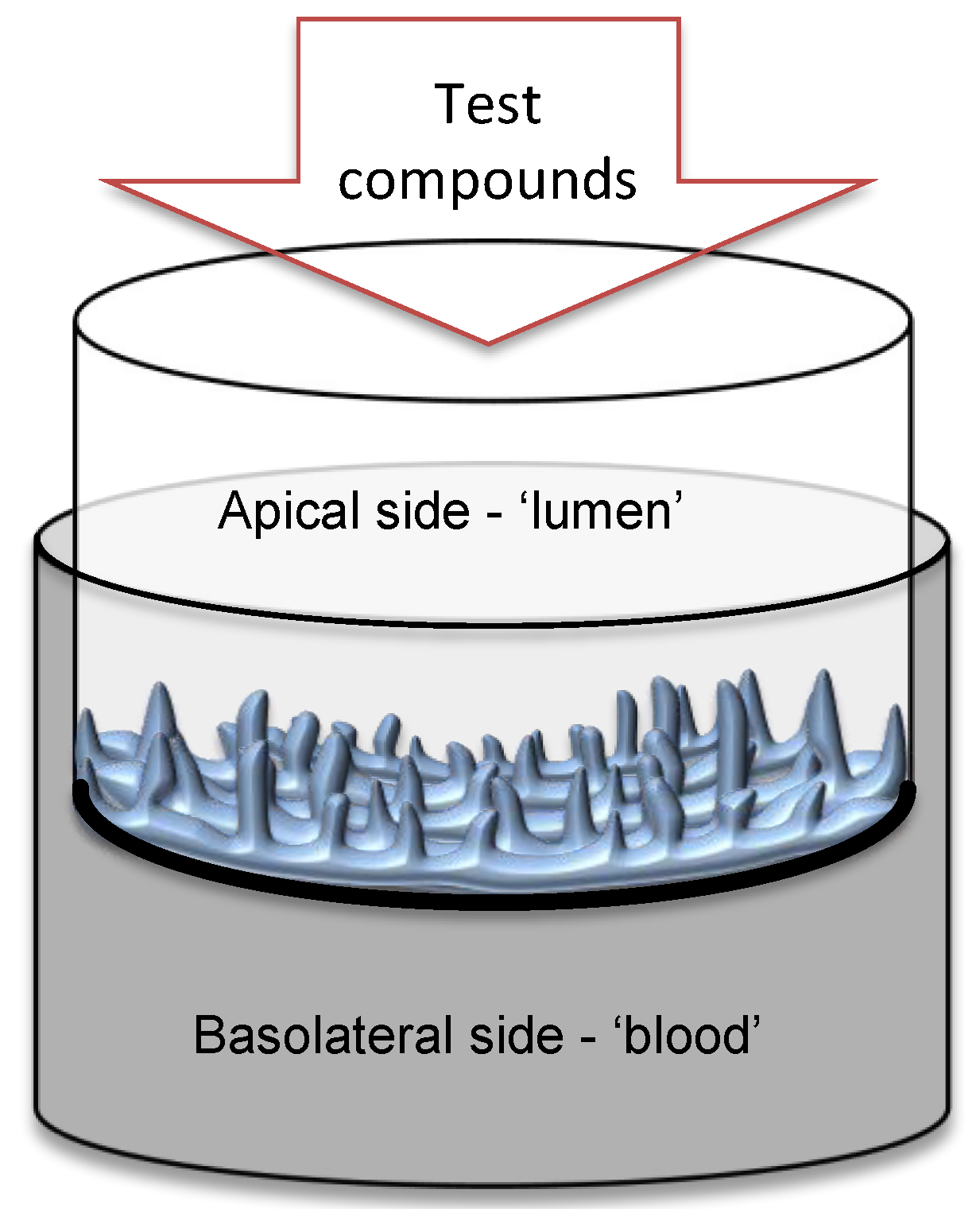
2.4. EpiIntestinal Human Tissue Model (SMI-100)
2.5. Tissue Experimental Design
2.6. TEER Measurement in the Tissue Model
2.7. MTT Assay
2.8. Passage of Lucifer Yellow (LY)
2.9. Histology and Microscopy
2.10. Gut Microbiome Sequencing
2.11. Enzymology and Protein Detection
2.12. Statistical Analysis
3. Results
3.1. In Vivo Study
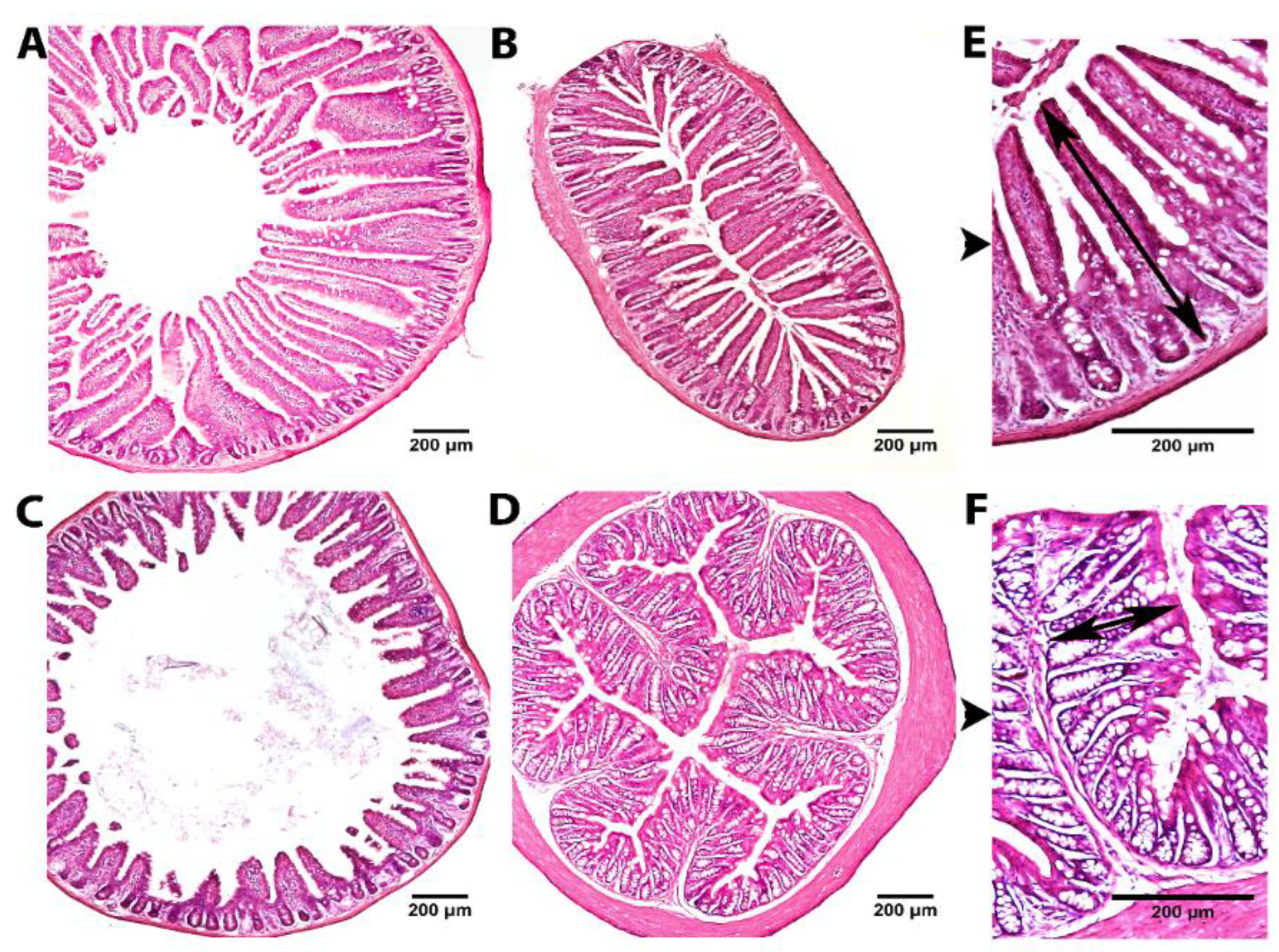
3.1.1. Histological Examination and Morphometric Analysis of the Intestinal Tissue
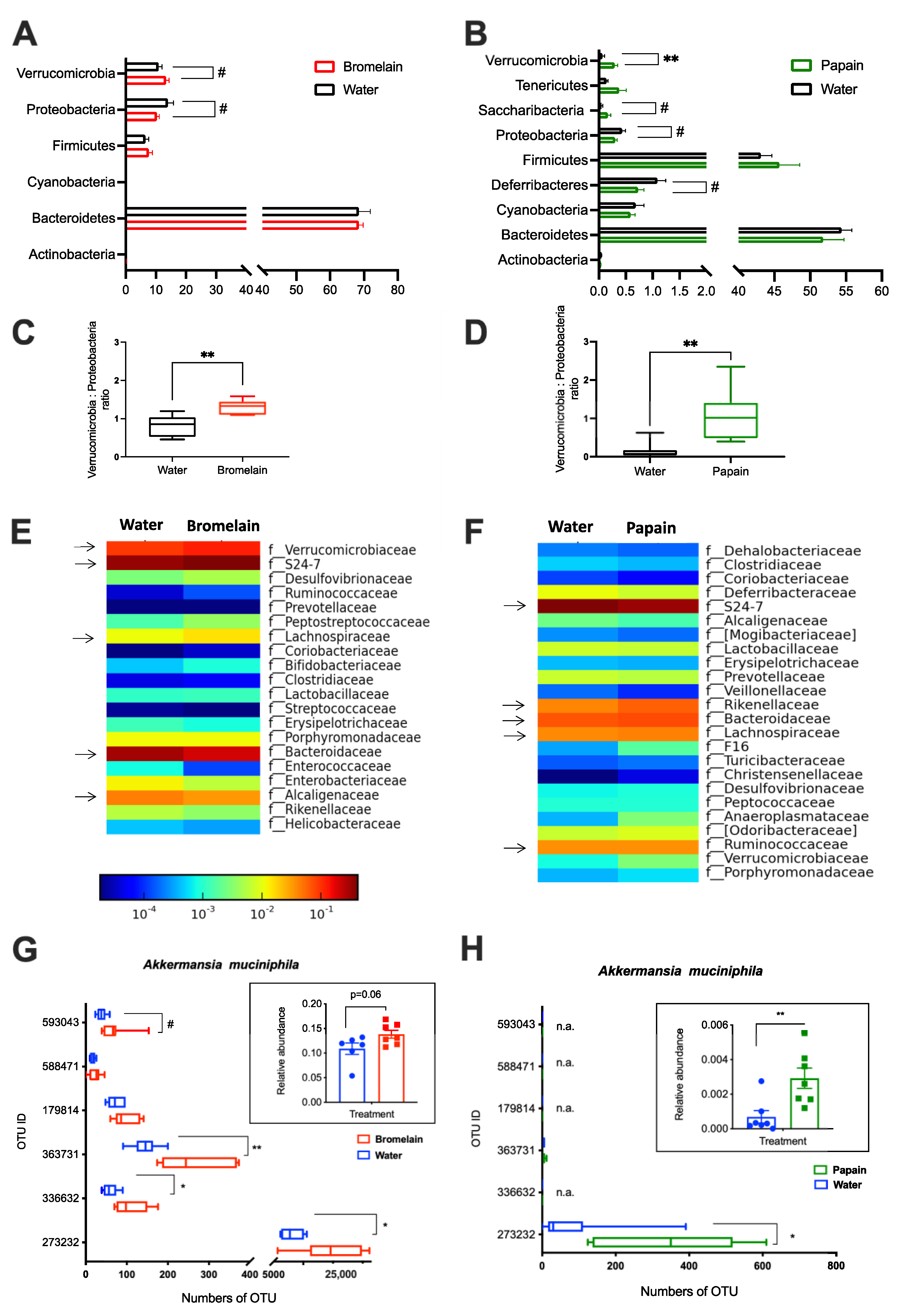
3.1.2. Microbiota Composition Analysis
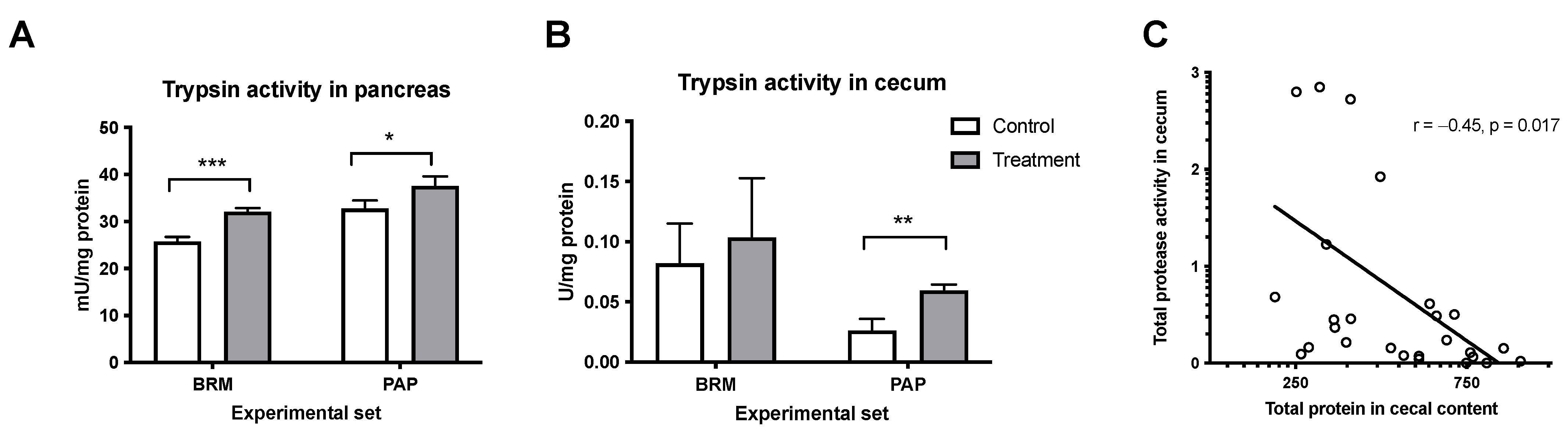
3.1.3. Effect of Plant Enzymes on Protein-Digestive Properties
3.2. In Vitro Study
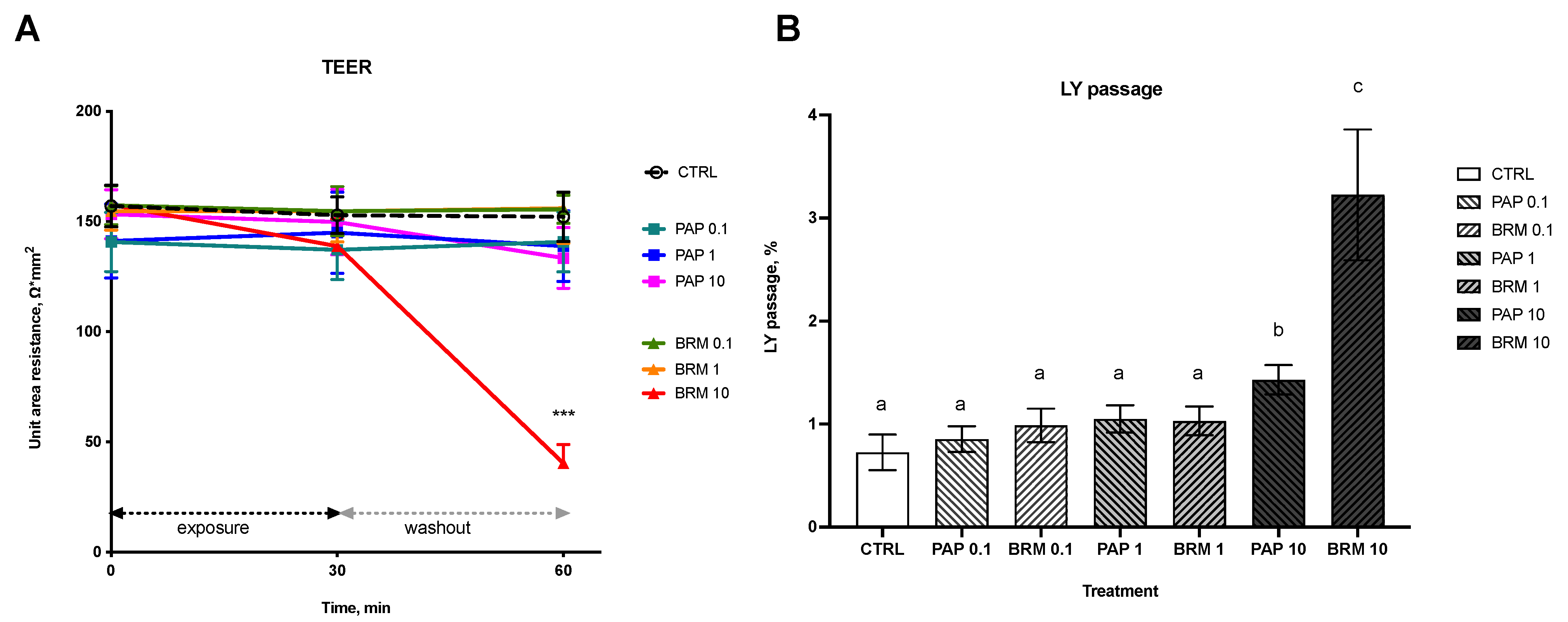
3.2.1. Intestinal Integrity
3.2.2. Intestinal Permeability and Cell Viability
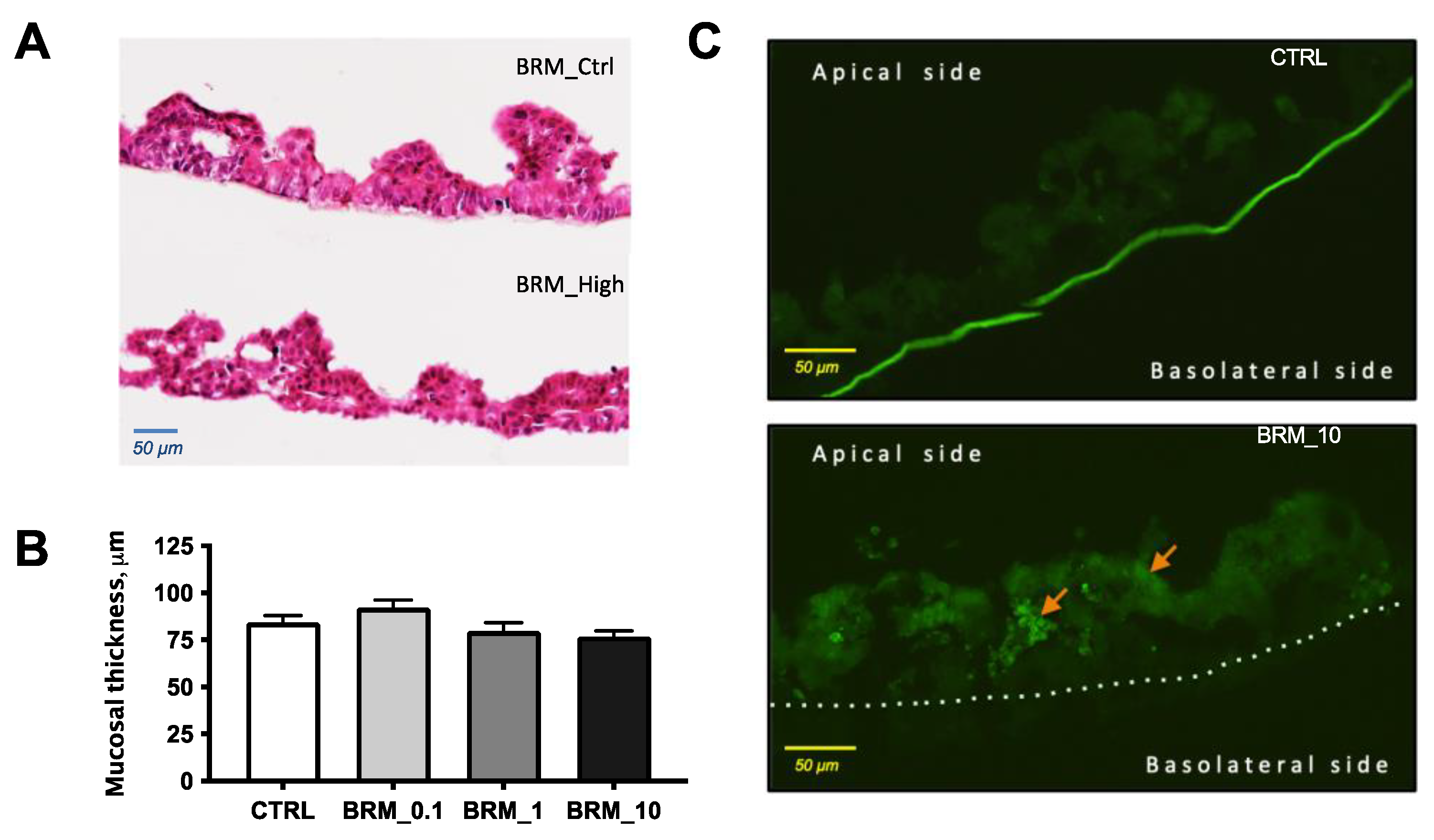
3.2.3. Histology of Reconstructed Tissues
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Borrelli, F.; Capasso, R.; Severino, B.; Fiorino, F.; Aviello, G.; De Rosa, G.; Mazzella, M.; Romano, B.; Capasso, F.; Fasolino, I.; et al. Inhibitory Effects of Bromelain, a Cysteine Protease Derived from Pineapple Stem (Ananas comosus), on Intestinal Motility in Mice. Neurogastroenterol. Motil. 2011, 23, 745-e331. [Google Scholar] [CrossRef]
- Tacias-Pascacio, V.G.; Morellon-Sterling, R.; Castañeda-Valbuena, D.; Berenguer-Murcia, Á.; Kamli, M.R.; Tavano, O.; Fernandez-Lafuente, R. Immobilization of Papain: A Review. Int. J. Biol. Macromol. 2021, 188, 94–113. [Google Scholar] [CrossRef]
- de Lencastre Novaes, L.C.; Jozala, A.F.; Lopes, A.M.; de Carvalho Santos-Ebinuma, V.; Mazzola, P.G.; Pessoa Junior, A. Stability, Purification, and Applications of Bromelain: A Review. Biotechnol. Prog. 2016, 32, 5–13. [Google Scholar] [CrossRef]
- Jančič, U.; Gorgieva, S. Bromelain and Nisin: The Natural Antimicrobials with High Potential in Biomedicine. Pharmaceutics 2021, 14, 76. [Google Scholar] [CrossRef]
- Pizzorno, J.E.; Murray, M.T. Textbook of Natural Medicine—E-Book; Elsevier Health Sciences: Amsterdam, The Netherlands, 2020; ISBN 978-0-323-52380-6. [Google Scholar]
- Bottega, R.; Persico, I.; De Seta, F.; Romano, F.; Di Lorenzo, G. Anti-Inflammatory Properties of a Proprietary Bromelain Extract (BromeyalTM) after in Vitro Simulated Gastrointestinal Digestion. Int. J. Immunopathol. Pharm. 2021, 35, 20587384211034690. [Google Scholar] [CrossRef]
- Tallei, T.E.; Fatimawali; Yelnetty, A.; Idroes, R.; Kusumawaty, D.; Emran, T.B.; Yesiloglu, T.Z.; Sippl, W.; Mahmud, S.; Alqahtani, T.; et al. An Analysis Based on Molecular Docking and Molecular Dynamics Simulation Study of Bromelain as Anti-SARS-CoV-2 Variants. Front. Pharmacol. 2021, 12, 717757. [Google Scholar] [CrossRef]
- Castell, J.V.; Friedrich, G.; Kuhn, C.S.; Poppe, G.E. Intestinal Absorption of Undegraded Proteins in Men: Presence of Bromelain in Plasma after Oral Intake. Am. J. Physiol. Gastrointest. Liver Physiol. 1997, 273, G139–G146. [Google Scholar] [CrossRef]
- Shiew, P.S.; Fang, Y.L.; Majid, F.A.A. In Vitro Study of Bromelain Activity in Artificial Stomach Juice and Blood. In Proceedings of the 3rd International Conference on Biotechnology for the Wellness Industry, Kuala Lumpur, Malaysia, 8–9 October 2010; pp. 8–9. [Google Scholar]
- Ataide, J.A.; Gérios, E.F.; Mazzola, P.G.; Souto, E.B. Bromelain-Loaded Nanoparticles: A Comprehensive Review of the State of the Art. Adv. Colloid Interface Sci. 2018, 254, 48–55. [Google Scholar] [CrossRef]
- Wen, S.; Huang, T.H.W.; Li, G.Q.; Yamahara, J.; Roufogalis, B.D.; Li, Y. Bromelain Improves Decrease in Defecation in Postoperative Rats: Modulation of Colonic Gene Expression of Inducible Nitric Oxide Synthase. Life Sci. 2006, 78, 995–1002. [Google Scholar] [CrossRef] [PubMed]
- Rathnavelu, V.; Alitheen, N.B.; Sohila, S.; Kanagesan, S.; Ramesh, R. Potential Role of Bromelain in Clinical and Therapeutic Applications. Biomed. Rep. 2016, 5, 283–288. [Google Scholar] [CrossRef]
- Glade, M.J.; Kendra, D.; Kaminski, M.V. Improvement in Protein Utilization in Nursing-Home Patients on Tube Feeding Supplemented with an Enzyme Product Derived from Aspergillus Niger and Bromelain. Nutrition 2001, 17, 348–350. [Google Scholar] [CrossRef]
- Fernández-Lucas, J.; Castañeda, D.; Hormigo, D. New Trends for a Classical Enzyme: Papain, a Biotechnological Success Story in the Food Industry. Trends Food Sci. Technol. 2017, 68, 91–101. [Google Scholar] [CrossRef]
- Eshamah, H.; Han, I.; Naas, H.; Acton, J.; Dawson, P. Antibacterial Effects of Natural Tenderizing Enzymes on Different Strains of Escherichia coli O157:H7 and Listeria monocytogenes on Beef. Meat Sci. 2014, 96, 1494–1500. [Google Scholar] [CrossRef] [PubMed]
- Osato, J.A.; Santiago, L.A.; Remo, G.M.; Cuadra, M.S.; Mori, A. Antimicrobial and Antioxidant Activities of Unripe Papaya. Life Sci. 1993, 53, 1383–1389. [Google Scholar] [CrossRef]
- Amini, N.; Setiasih, S.; Handayani, S.; Hudiyono, S.; Saepudin, E. Potential Antibacterial Activity of Partial Purified Bromelain from Pineapple Core Extracts Using Acetone and Ammonium Sulphate against Dental Caries-Causing Bacteria. AIP Conf. Proc. 2018, 2023, 020071. [Google Scholar] [CrossRef]
- Mynott, T.L.; Luke, R.K.; Chandler, D.S. Oral Administration of Protease Inhibits Enterotoxigenic Escherichia coli Receptor Activity in Piglet Small Intestine. Gut 1996, 38, 28–32. [Google Scholar] [CrossRef] [Green Version]
- Begum, M.; Li, H.L.; Hossain, M.M.; Kim, I.H. Dietary Bromelain-C.3.4.22.32 Supplementation Improves Performance and Gut Health in Sows and Piglets. Livest. Sci. 2015, 180, 177–182. [Google Scholar] [CrossRef]
- Saw, C.-Y.; Kao, W.-M.; Chen, P.-Y.; Chew, B.-S.; Liew, Y.; Chau, C.-F. Changes in Different Fecal Parameters with Administration of Bromelain and Inulin in a Rat Model. CyTA-J. Food 2018, 16, 980–985. [Google Scholar] [CrossRef]
- Mosmann, T. Rapid Colorimetric Assay for Cellular Growth and Survival: Application to Proliferation and Cytotoxicity Assays. J. Immunol Methods 1983, 65, 55–63. [Google Scholar] [CrossRef]
- Rastogi, H.; Pinjari, J.; Honrao, P.; Praband, S.; Somani, R. The Impact of Permeability Enhancers on Assessment for Monolayer of Colon Adenocarcinoma Cell Line (Caco-2) Used In In Vitro Permeability Assay. J. Drug Delivery Ther. 2013, 3, 20–29. [Google Scholar] [CrossRef]
- Ki, Y.; Kim, W.; Cho, H.; Ahn, K.; Choi, Y.; Kim, D. The Effect of Probiotics for Preventing Radiation-Induced Morphological Changes in Intestinal Mucosa of Rats. J. Korean Med. Sci. 2014, 29, 1372–1378. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kozich, J.J.; Westcott, S.L.; Baxter, N.T.; Highlander, S.K.; Schloss, P.D. Development of a Dual-Index Sequencing Strategy and Curation Pipeline for Analyzing Amplicon Sequence Data on the MiSeq Illumina Sequencing Platform. Appl. Environ. Microbiol. 2013, 79, 5112–5120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Peña, A.G.; Goodrich, J.K.; Gordon, J.I.; et al. QIIME Allows Analysis of High-Throughput Community Sequencing Data. Nat Methods 2010, 7, 335–336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- DeSantis, T.Z.; Hugenholtz, P.; Larsen, N.; Rojas, M.; Brodie, E.L.; Keller, K.; Huber, T.; Dalevi, D.; Hu, P.; Andersen, G.L. Greengenes, a Chimera-Checked 16S RRNA Gene Database and Workbench Compatible with ARB. Appl. Environ. Microbiol. 2006, 72, 5069–5072. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anson, M.L. The Estimation of Pepsin, Trypsin, Papain, and Cathepsin with Hemoglobin. J. Gen. Physiol. 1938, 22, 79–89. [Google Scholar] [CrossRef]
- Wang, X.; Hui, H.; Yu, A.; Jiang, Z.; Yu, H.; Zou, L.; Teng, Y. Characterization of Binding Interaction of Triclosan and Trypsin. Environ. Sci. Pollut. Res. 2020, 27, 13409–13416. [Google Scholar] [CrossRef]
- Pavan, R.; Jain, S.; Shraddha; Kumar, A. Properties and Therapeutic Application of Bromelain: A Review. Biotechnol. Res. Int. 2012, 2012, e976203. [Google Scholar] [CrossRef] [Green Version]
- Kane, S.; Goldberg, M.J. Use of Bromelain for Mild Ulcerative Colitis. Ann. Intern. Med. 2000, 132, 680. [Google Scholar] [CrossRef]
- Prykhodko, O.; Pierzynowski, S.G.; Nikpey, E.; Sureda, E.A.; Fedkiv, O.; Weström, B.R. Pancreatic and Pancreatic-Like Microbial Proteases Accelerate Gut Maturation in Neonatal Rats. PLoS ONE 2015, 10, e0116947. [Google Scholar] [CrossRef]
- Yuan, L.; Li, M.; Qiao, Y.; Wang, H.; Cui, L.; Wang, M. The Impact of Berberine on Intestinal Morphology, Microbes, and Immune Function of Broilers in Response to Necrotic Enteritis Challenge. BioMed Res. Int. 2021, 2021, e1877075. [Google Scholar] [CrossRef]
- Xu, Z.; Hu, C.; Xia, M.; Zhan, X.; Wang, M. Effects of Dietary Fructooligosaccharide on Digestive Enzyme Activities, Intestinal Microflora and Morphology of Male Broilers. Poult. Sci. 2003, 82, 1030–1036. [Google Scholar] [CrossRef] [PubMed]
- Jansen, C.; Ihse, I.; Axelson, J. Epidermal Growth Factor Induces Increased Mucosal Thickness of the Small Intestine in Mouse. ESR 1997, 29, 447–454. [Google Scholar] [CrossRef] [PubMed]
- Leeming, E.R.; Louca, P.; Gibson, R.; Menni, C.; Spector, T.D.; Le Roy, C.I. The Complexities of the Diet-Microbiome Relationship: Advances and Perspectives. Genome Med. 2021, 13, 10. [Google Scholar] [CrossRef]
- Campos, D.A.; Coscueta, E.R.; Vilas-Boas, A.A.; Silva, S.; Teixeira, J.A.; Pastrana, L.M.; Pintado, M.M. Impact of Functional Flours from Pineapple By-Products on Human Intestinal Microbiota. J. Funct. Foods 2020, 67, 103830. [Google Scholar] [CrossRef]
- Rizzatti, G.; Lopetuso, L.R.; Gibiino, G.; Binda, C.; Gasbarrini, A. Proteobacteria: A Common Factor in Human Diseases. BioMed Res. Int. 2017, 2017, e9351507. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vester-Andersen, M.K.; Mirsepasi-Lauridsen, H.C.; Prosberg, M.V.; Mortensen, C.O.; Träger, C.; Skovsen, K.; Thorkilgaard, T.; Nøjgaard, C.; Vind, I.; Krogfelt, K.A.; et al. Increased Abundance of Proteobacteria in Aggressive Crohn’s Disease Seven Years after Diagnosis. Sci. Rep. 2019, 9, 13473. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shin, N.-R.; Whon, T.W.; Bae, J.-W. Proteobacteria: Microbial Signature of Dysbiosis in Gut Microbiota. Trends Biotechnol. 2015, 33, 496–503. [Google Scholar] [CrossRef]
- Bárcena, C.; Valdés-Mas, R.; Mayoral, P.; Garabaya, C.; Durand, S.; Rodríguez, F.; Fernández-García, M.T.; Salazar, N.; Nogacka, A.M.; Garatachea, N.; et al. Healthspan and Lifespan Extension by Fecal Microbiota Transplantation into Progeroid Mice. Nat. Med. 2019, 25, 1234–1242. [Google Scholar] [CrossRef]
- de la Cuesta-Zuluaga, J.; Mueller, N.T.; Corrales-Agudelo, V.; Velásquez-Mejía, E.P.; Carmona, J.A.; Abad, J.M.; Escobar, J.S. Metformin Is Associated with Higher Relative Abundance of Mucin-Degrading Akkermansia muciniphila and Several Short-Chain Fatty Acid-Producing Microbiota in the Gut. Diabetes Care 2017, 40, 54–62. [Google Scholar] [CrossRef] [Green Version]
- Ottman, N.; Geerlings, S.Y.; Aalvink, S.; de Vos, W.M.; Belzer, C. Action and Function of Akkermansia muciniphila in Microbiome Ecology, Health and Disease. Best Pract. Res. Clin. Gastroenterol. 2017, 31, 637–642. [Google Scholar] [CrossRef]
- Derrien, M.; Belzer, C.; de Vos, W.M. Akkermansia Muciniphila and Its Role in Regulating Host Functions. Microb. Pathog. 2017, 106, 171–181. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, J.; Lin, S.; Vanhoutte, P.M.; Woo, C.W.; Xu, A. Akkermansia Muciniphila Protects Against Atherosclerosis by Preventing Metabolic Endotoxemia-Induced Inflammation in Apoe−/− Mice. Circulation 2016, 133, 2434–2446. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Everard, A.; Belzer, C.; Geurts, L.; Ouwerkerk, J.P.; Druart, C.; Bindels, L.B.; Guiot, Y.; Derrien, M.; Muccioli, G.G.; Delzenne, N.M.; et al. Cross-Talk between Akkermansia muciniphila and Intestinal Epithelium Controls Diet-Induced Obesity. Proc Natl Acad Sci U S A. 2013, 110, 9066–9071. [Google Scholar] [CrossRef] [Green Version]
- Hashimoto, N.; Hara, H. Dietary Amino Acids Promote Pancreatic Protease Synthesis at the Translation Stage in Rats. J. Nutr. 2003, 133, 3052–3057. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hale, L.P. Proteolytic Activity and Immunogenicity of Oral Bromelain within the Gastrointestinal Tract of Mice. Int. Immunopharmacol. 2004, 4, 255–264. [Google Scholar] [CrossRef] [PubMed]
- Mo, W.Y.; Lau, R.S.S.; Kwok, A.C.K.; Wong, M.H. Use of Soybean Meal and Papain to Partially Replace Animal Protein for Culturing Three Marine Fish Species: Fish Growth and Water Quality. Environ. Pollut. 2016, 219, 815–820. [Google Scholar] [CrossRef]
- Nishiyama, H.; Nagai, T.; Kudo, M.; Okazaki, Y.; Azuma, Y.; Watanabe, T.; Goto, S.; Ogata, H.; Sakurai, T. Supplementation of Pancreatic Digestive Enzymes Alters the Composition of Intestinal Microbiota in Mice. Biochem. Biophys. Res. Commun. 2018, 495, 273–279. [Google Scholar] [CrossRef] [Green Version]
- Guggi, D.; Bernkop-Schnürch, A. Improved Paracellular Uptake by the Combination of Different Types of Permeation Enhancers. Int. J. Pharm. 2005, 288, 141–150. [Google Scholar] [CrossRef]
- Grabovac, V.; Schmitz, T.; Föger, F.; Bernkop-Schnürch, A. Papain: An Effective Permeation Enhancer for Orally Administered Low Molecular Weight Heparin. Pharm. Res. 2007, 24, 1001–1006. [Google Scholar] [CrossRef]
- Ayehunie, S.; Landry, T.; Stevens, Z.; Armento, A.; Hayden, P.; Klausner, M. Human Primary Cell-Based Organotypic Microtissues for Modeling Small Intestinal Drug Absorption. Pharm. Res. 2018, 35, 72. [Google Scholar] [CrossRef]
- Ferreira, E.H.B.; Melo, K.C.; da Silva, C.O.; da Silva, J.S.; da Vilanova, L.S.M.; Siqueira, F.F.F.S.; Teixeira, S.A.M.; Siqueira, H.D.S.; Junior, R.N.C.M.; Cunha, H.G.S.S.; et al. Mucolytic action of bromelain: An integrative review. Res. Soc. Dev. 2021, 10, e557101321495. [Google Scholar] [CrossRef]
- Lorkowski, G. Gastrointestinal Absorption and Biological Activities of Serine and Cysteine Proteases of Animal and Plant Origin: Review on Absorption of Serine and Cysteine Proteases. Int. J. Physiol. Pathophysiol. Pharm. 2012, 4, 10–27. [Google Scholar]
- Annaházi, A.; Ferrier, L.; Bézirard, V.; Lévêque, M.; Eutamène, H.; Ait-Belgnaoui, A.; Coëffier, M.; Ducrotté, P.; Róka, R.; Inczefi, O.; et al. Luminal Cysteine-Proteases Degrade Colonic Tight Junction Structure and Are Responsible for Abdominal Pain in Constipation-Predominant IBS. Am. J. Gastroenterol. 2013, 108, 1322–1331. [Google Scholar] [CrossRef] [PubMed]
- Bock, U.; Kolac, C.; Borchard, G.; Koch, K.; Fuchs, R.; Streichhan, P.; Lehr, C.-M. Transport of Proteolytic Enzymes Across Caco-2 Cell Monolayers. Pharm. Res. 1998, 15, 1393–1400. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Mucosal thickness, μm | Bromelain | Papain | ||
|---|---|---|---|---|
| Control | Treatment | Control | Treatment | |
| Small intestine: | ||||
| Duodenum | 643.3 ± 25.7 | 719.4 ± 34.5 # | 601.2 ± 16.3 | 584.3 ± 14.9 |
| Jejunum | 458.2 ± 38.5 a | 453.4 ± 14.9 | 373.1 ± 13.9 b | 369.3 ± 14.3 |
| Ileum | 270.2 ± 28.8 | 356.2 ± 24.0 * | 313.1 ± 14.3 | 317.1 ± 11.6 |
| Large intestine: | ||||
| Upper colon | 194.2 ± 27.8 a | 187.1 ± 28.8 | 121.3 ± 14.1 b | 139.7 ± 15.63 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kostiuchenko, O.; Kravchenko, N.; Markus, J.; Burleigh, S.; Fedkiv, O.; Cao, L.; Letasiova, S.; Skibo, G.; Fåk Hållenius, F.; Prykhodko, O. Effects of Proteases from Pineapple and Papaya on Protein Digestive Capacity and Gut Microbiota in Healthy C57BL/6 Mice and Dose-Manner Response on Mucosal Permeability in Human Reconstructed Intestinal 3D Tissue Model. Metabolites 2022, 12, 1027. https://doi.org/10.3390/metabo12111027
Kostiuchenko O, Kravchenko N, Markus J, Burleigh S, Fedkiv O, Cao L, Letasiova S, Skibo G, Fåk Hållenius F, Prykhodko O. Effects of Proteases from Pineapple and Papaya on Protein Digestive Capacity and Gut Microbiota in Healthy C57BL/6 Mice and Dose-Manner Response on Mucosal Permeability in Human Reconstructed Intestinal 3D Tissue Model. Metabolites. 2022; 12(11):1027. https://doi.org/10.3390/metabo12111027
Chicago/Turabian StyleKostiuchenko, Olha, Nadiia Kravchenko, Jan Markus, Stephen Burleigh, Olexandr Fedkiv, Ling Cao, Silvia Letasiova, Galyna Skibo, Frida Fåk Hållenius, and Olena Prykhodko. 2022. "Effects of Proteases from Pineapple and Papaya on Protein Digestive Capacity and Gut Microbiota in Healthy C57BL/6 Mice and Dose-Manner Response on Mucosal Permeability in Human Reconstructed Intestinal 3D Tissue Model" Metabolites 12, no. 11: 1027. https://doi.org/10.3390/metabo12111027
APA StyleKostiuchenko, O., Kravchenko, N., Markus, J., Burleigh, S., Fedkiv, O., Cao, L., Letasiova, S., Skibo, G., Fåk Hållenius, F., & Prykhodko, O. (2022). Effects of Proteases from Pineapple and Papaya on Protein Digestive Capacity and Gut Microbiota in Healthy C57BL/6 Mice and Dose-Manner Response on Mucosal Permeability in Human Reconstructed Intestinal 3D Tissue Model. Metabolites, 12(11), 1027. https://doi.org/10.3390/metabo12111027