

Moringa oleifera and Propolis in Cattle Nutrition: Characterization of Metabolic Activities in the Rumen In Vitro

 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Design and Treatments
2.2. RUSITEC Procedure
2.3. Sampling, Daily Measurements and Laboratory Analyses
2.4. Statistical Analysis
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dodd, D.; Mackie, R.I.; Cann, I.K. Xylan degradation, a metabolic property shared by rumen and human colonic Bacteroidetes. Mol. Microbiol. 2011, 79, 292–304. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jayasuriya, M.C.N. Use of crop residues and agro-industrial by-products in ruminant production systems in developing countries. BSAP Occas. Publ. 1993, 16, 47–55. [Google Scholar] [CrossRef]
- Xia, C.; Rahman, M.A.U.; Yang, H.; Shao, T.; Qiu, Q.; Su, H.; Cao, B. Effect of increased dietary crude protein levels on production performance, nitrogen utilisation, blood metabolites and ruminal fermentation of Holstein bulls. Asian-Australas. J. Anim. Sci. 2018, 31, 1643–1653. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Islam, Z.; Islam, S.M.; Hossen, F.; Mahtab-ul-Islam, K.; Hasan, M.; Karim, R. Moringa oleifera is a prominent source of nutrients with potential health benefits. Int. J. Food Sci. 2021, 2021, 6627265. [Google Scholar] [CrossRef] [PubMed]
- Su, B.; Chen, X. Current status and potential of Moringa oleifera leaf as an alternative protein source for animal feeds. Front. Vet. Sci. 2020, 7, 53–66. [Google Scholar] [CrossRef] [Green Version]
- Godino, M.; Arias, C.; Izquierdo, M.I. Moringa oleifera: Potential areas of cultivation on the Iberian Peninsula. Acta Hortic. 2017, 1158, 405–412. [Google Scholar] [CrossRef]
- Gopalakrishnan, L.; Doriya, K.; Kumar, D.S. Moringa oleifera: A review on nutritive importance and its medicinal application. Food Sci. Hum. Wellness 2016, 5, 49–56. [Google Scholar] [CrossRef] [Green Version]
- Patil, D.; Vaknin, Y.; Rytwo, G.; Lakemond, C.; Benjamin, O. Characterization of Moringa oleifera leaf and seed protein extract functionality in emulsion model system. Innov. Food Sci. Emerg. Technol. 2022, 75, 102903. [Google Scholar] [CrossRef]
- Kumar, N.; Pareek, S. Bioactive compounds of moringa (Moringa species). In Bioactive Compounds in Underutilized Vegetables and Legumes, 1st ed.; Murthy, H.N., Paek, K.Y., Eds.; Springer: Cham, Switzerland, 2021; pp. 1–22. [Google Scholar] [CrossRef]
- Soltan, Y.A.; Morsy, A.S.; Hashem, N.M.; Sallam, S.M. Utilization of Moringa oleifera in ruminant nutrition. In Proceedings of the Sustainable Development of Livestock’s Production Systems “(SDLPS)”, Alexandra University, Bab Sharqi, Egypt, 7–9 November 2017; pp. 7–9. [Google Scholar]
- Mohammed, S.; Manan, F.A. Analysis of total phenolics, tannins and flavonoids from Moringa oleifera seed extract. J. Chem. Pharm. Res. 2015, 7, 132–135. [Google Scholar]
- Karim, R.A.; Amin, M.R.; Moniruzzaman, M.; Sarker, M.B.; Kabir, A.K.M.A. Effect of Moringa oleifera leaf on the efficiency to increase protein supply to ruminants. BJAS 2015, 44, 46–51. [Google Scholar] [CrossRef]
- Olivares-Palma, S.M.; Meale, S.J.; Pereira, L.G.R.; Machado, F.S.; Carneiro, H.; Lopes, F.C.F.; Maurício, R.M.; Chaves, A.V. In vitro fermentation, digestion kinetics and methane production of oilseed press cakes from biodiesel production. Asian-Australas. J. Anim. Sci. 2013, 26, 1102–1112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morsy, T.A.; Gouda, G.A.; Kholif, A.E. In vitro fermentation and production of methane and carbon dioxide from rations containing Moringa oleifera leave silage as a replacement of soybean meal: In vitro assessment. Environ. Sci. Pollut. Res. 2022, 29, 69743–69752. [Google Scholar] [CrossRef] [PubMed]
- ÖZTURK, H.; Pekcan, M.; Sireli, M.; Fidanci, U.R. Effects of propolis on in vitro rumen microbial fermentation. Ankara Univ. Vet. Fak. 2010, 57, 217–221. [Google Scholar]
- Piccinelli, A.L.; Mencherini, T.; Celano, R.; Mouhoubi, Z.; Tamendjari, A.; Aquino, R.P.; Rastrelli, L. Chemical composition and antioxidant activity of Algerian propolis. J. Agric. Food Chem. 2013, 61, 5080–5088. [Google Scholar] [CrossRef] [PubMed]
- Ahangari, Z.; Naseri, M.; Vatandoost, F. Propolis: Chemical composition and its applications in endodontics. Iran. Endod. J. 2018, 13, 285–292. [Google Scholar] [CrossRef]
- Kumazawa, S. February. Bioactive compounds in bee propolis for drug discovery. AIP Conf. Proc. 2018, 1933, 030001. [Google Scholar] [CrossRef]
- Mahmood, M.; Petri, R.M.; Gavrău, A.; Zebeli, Q.; Khiaosa-ard, R. Betaine addition as a potent ruminal fermentation modulator under hyperthermal and hyperosmotic conditions in vitro. J. Sci. Food Agric. 2020, 100, 2261–2271. [Google Scholar] [CrossRef]
- Thomas, J.W. Protein requirements of milking cows. J. Dairy Sci. 1971, 54, 1629–1636. [Google Scholar] [CrossRef]
- Santos, N.W.; Yoshimura, E.H.; Machado, E.; Matumoto-Pintro, P.T.; Montanher, P.F.; Visentainer, J.V.; dos Santos, G.T.; Zeoula, L.M. Antioxidant effects of a propolis extract and vitamin E in blood and milk of dairy cows fed diet containing flaxseed oil. Livest. Sci. 2016, 191, 132–138. [Google Scholar] [CrossRef]
- BMSGPK. Verordnung der Bundesministerin für Gesundheit und Frauen uber die Mindestanforderungen fur die Haltung von Pferden und Pferdeartigen, Schweinen, Rindern, Schafen, Ziegen, Schalenwild, Lamas, Kaninchen, Hausgeflugel, Strausen und Nutzfischen (1. Tierhaltungsverordnung) StF: BGBl. II Nr. 485/2004; Bundeskanzleramt Osterreich: Vienna, Austria, 2004. [Google Scholar]
- VDLUFA. Die Chemische Untersuchung von Futtermitteln. In Handbuch der Landwirtschaftlichen Versuchs-und Untersuchungsmethodik (VDLUFA-Methodenbuch), Bd., Vol. III; VDLUFA-Verlag: Darmstadt, Germany, 2012. [Google Scholar]
- Weatherburn, M.W. Phenol-hypochlorite reaction for determination of ammonia. Anal. Chem. 1967, 39, 971–974. [Google Scholar] [CrossRef]
- Soliva, C.R.; Hess, H.D. Measuring methane emission of ruminants by in vitro and in vivo techniques. In Measuring Methane Production from Ruminants, 1st ed.; Makkar, H.P., Vercoe, P.E., Eds.; Springer: Dordrecht, Holland, 2007; pp. 15–31. [Google Scholar] [CrossRef]
- Benzie, I.F.; Strain, J.J. The ferric reducing ability of plasma (FRAP) as a measure of “antioxidant power”: The FRAP assay. Anal. Biochem. 1996, 239, 70–76. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weiss, W.P.; Tebbe, A.W. Estimating digestible energy values of feeds and diets and integrating those values into net energy systems. Transl. Anim. Sci. 2019, 3, 953–961. [Google Scholar] [CrossRef] [PubMed]
- El-Naggar, S.I. Impact of incorporating Moringa oleifera seed cake as protein source in growing lambs ration. CIGR J. 2017, 2017, 289–292. [Google Scholar]
- Kakengi, A.M.V.; Shem, M.N.; Sarwatt, S.V.; Fujihara, T. Can Moringa oleifera be used as a protein supplement for ruminants? Asian-Australas. J. Anim. Sci. 2005, 18, 42–47. [Google Scholar] [CrossRef]
- Lins, T.D.A.; Terry, S.A.; Silva, R.R.; Pereira, L.G.R.; Jancewicz, L.J.; He, M.L.; Wang, Y.; McAllister, T.A.; Chaves, A.V. Effects of the inclusion of Moringa oleifera seed on rumen fermentation and methane production in a beef cattle diet using the rumen simulation technique (Rusitec). Animal 2019, 13, 283–291. [Google Scholar] [CrossRef]
- Nocek, J.E.; Russell, J. Protein and energy as an integrated system. Relationship of ruminal protein and carbohydrate availability to microbial synthesis and milk production. J. Dairy Sci. 1988, 71, 2070–2107. [Google Scholar] [CrossRef]
- Aboamer, A.A.; Ebeid, H.M.; Shaaban, M.M.; Gaward, R.M.A.; Mostafa, M.M.; Abdalla, A.M. Effect of feeding moringa seed cake as an alternative protein source in lactating ewes rations. Int. J. Dairy Sci. 2020, 15, 80–87. [Google Scholar] [CrossRef] [Green Version]
- Aschenbach, J.R.; Kristensen, N.B.; Donkin, S.S.; Hammon, H.M.; Penner, G.B. Gluconeogenesis in dairy cows: The secret of making sweet milk from sour dough. IUBMB Life 2010, 62, 869–877. [Google Scholar] [CrossRef]
- Soliva, C.R.; Kreuzer, M.; Foidl, N.; Foidl, G.; Machmüller, A.; Hess, H.D. Feeding value of whole and extracted Moringa oleifera leaves for ruminants and their effects on ruminal fermentation in vitro. Anim. Feed Sci. Technol. 2005, 118, 47–62. [Google Scholar] [CrossRef]
- Etcheverry, P.; Grusak, M.A.; Fleige, L.E. Application of in vitro bioaccessibility and bioavailability methods for calcium, carotenoids, folate, iron, magnesium, polyphenols, zinc, and vitamins B6, B12, D, and E. Front. Physiol. 2012, 3, 317. [Google Scholar] [CrossRef] [Green Version]
- Grosshagauer, S.; Pirkwieser, P.; Kraemer, K.; Somoza, V. The future of Moringa foods: A food chemistry perspective. Front. Nutr. 2021, 8, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Guyader, J.; Ungerfeld, E.M.; Beauchemin, K.A. Redirection of metabolic hydrogen by inhibiting methanogenesis in the rumen simulation technique (RUSITEC). Front. Microbiol. 2017, 8, 393–409. [Google Scholar] [CrossRef] [Green Version]
- Vargas, J.E.; Andrés, S.; López-Ferreras, L.; Snelling, T.J.; Yáñez-Ruíz, D.R.; García-Estrada, C.; López, S. Dietary supplemental plant oils reduce methanogenesis from anaerobic microbial fermentation in the rumen. Sci. Rep. 2020, 10, 1613. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ibrahim, N.A.; Alimon, A.R.; Yaakub, H.; Samsudin, A.A.; Candyrine, S.C.L.; Wan Mohamed, W.N.; Mookiah, S. Effects of vegetable oil supplementation on rumen fermentation and microbial population in ruminant: A review. Trop. Anim. Health Prod. 2021, 53, 422. [Google Scholar] [CrossRef] [PubMed]
- Messana, J.D.; Berchielli, T.T.; Arcuri, P.B.; Reis, R.A.; Canesin, R.C.; Ribeiro, A.F.; Fernandes, J.J.D.R. Rumen fermentation and rumen microbes in Nellore steers receiving diets with different lipid contents. Rev. Bras. Zootec. 2013, 42, 204–212. [Google Scholar] [CrossRef] [Green Version]
- Vergara-Jimenez, M.; Almatrafi, M.M.; Fernandez, M.L. Bioactive components in Moringa oleifera leaves protect against chronic disease. Antioxidants 2017, 6, 91. [Google Scholar] [CrossRef] [Green Version]
- Oskoueian, E.; Abdullah, N.; Oskoueian, A. Effects of flavonoids on rumen fermentation activity, methane production, and microbial population. BioMed Res. Int. 2013, 2013, 349129. [Google Scholar] [CrossRef] [Green Version]
- Aboagye, I.A.; Oba, M.; Koenig, K.M.; Zhao, G.Y.; Beauchemin, K.A. Use of gallic acid and hydrolyzable tannins to reduce methane emission and nitrogen excretion in beef cattle fed a diet containing alfalfa silage. J. Anim. Sci. 2019, 97, 2230–2244. [Google Scholar] [CrossRef] [Green Version]
- Ku-Vera, J.C.; Jiménez-Ocampo, R.; Valencia-Salazar, S.S.; Montoya-Flores, M.D.; Molina-Botero, I.C.; Arango, J.; Gómez-Bravo, C.A.; Aguilar-Pérez, C.F.; Solorio-Sánchez, F.J. Role of secondary plant metabolites on enteric methane mitigation in ruminants. Front. Vet. Sci. 2020, 7, 584. [Google Scholar] [CrossRef]
- Morsy, A.S.; Soltan, Y.A.; El-Zaiat, H.M.; Alencar, S.M.D.; Abdalla, A.L. Bee propolis extract as a phytogenic feed additive to enhance diet digestibility, rumen microbial biosynthesis, mitigating methane formation and health status of late pregnant ewes. Anim. Feed Sci. Technol. 2021, 273, 114834. [Google Scholar] [CrossRef]
- Costa, J.B.G., Jr.; Zeoula, L.M.; Franco, S.L.; de Moura, L.P.; Valero, M.V.; Simioni, F.L.; da Paula, E.M.; Samensari, R.B. Effect of propolis product on digestibility and ruminal parameters in buffaloes consuming a forage-based diet. Ital. J. Anim. Sci. 2012, 11, e78. [Google Scholar] [CrossRef]
- Celińska-Janowicz, K.; Zaręba, I.; Lazarek, U.; Teul, J.; Tomczyk, M.; Pałka, J.; Miltyk, W. Constituents of propolis: Chrysin, caffeic acid, p-coumaric acid, and ferulic acid induce PRODH/POX-dependent apoptosis in human tongue squamous cell carcinoma cell (CAL-27). Front. Pharmacol. 2018, 9, 336. [Google Scholar] [CrossRef] [Green Version]
- Xu, J.; Ge, J.; He, X.; Sheng, Y.; Zheng, S.; Zhang, C.; Xu, W.; Huang, K. Caffeic acid reduces body weight by regulating gut microbiota in diet-induced-obese mice. J. Funct. Foods. 2020, 74, 104061. [Google Scholar] [CrossRef]
- Kačániová, M.; Rovná, K.; Arpášová, H.; Čuboň, J.; Hleba, L.; Pochop, J.; Kunová, S.; Haščík, P. In vitro and in vivo antimicrobial activity of propolis on the microbiota from gastrointestinal tract of chickens. J. Environ. 2012, 47, 1665–1671. [Google Scholar] [CrossRef]
- Penner, G.B.; Steele, M.A.; Aschenbach, J.R.; McBride, B.W. Ruminant Nutrition Symposium: Molecular adaptation of ruminal epithelia to highly fermentable diets. J. Anim. Sci. 2011, 89, 1108–1119. [Google Scholar] [CrossRef] [PubMed]


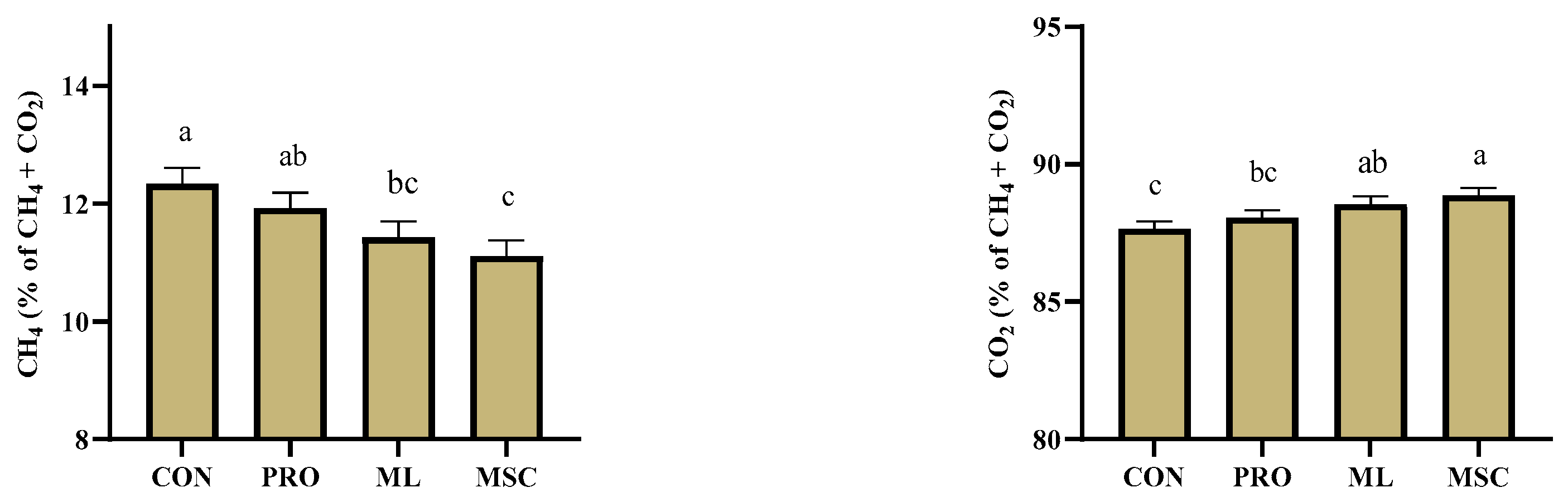{kind=link}
| Item | CON | PRO | ML | MSC |
|---|---|---|---|---|
| Ingredients | ||||
| Meadow hay | 503 | 502 | 465 | 483 |
| Concentrate 1 | 497 | 497 | 461 | 478 |
| Propolis | 0.0 | 1.0 | 0.0 | 0.0 |
| Moringa leaf powder | 0.0 | 0.0 | 74 | 0.0 |
| Moringa seed cake | 0.0 | 0.0 | 0.0 | 38 |
| Chemical composition | ||||
| Dry Matter | 894 | 894 | 897 | 895 |
| Organic Matter | 915 | 915 | 914 | 916 |
| Crude Protein | 99 | 99 | 112 | 114 |
| Ash | 85 | 85 | 86 | 84 |
| Neutral detergent fiber | 475 | 474 | 457 | 464 |
| Ether extract (crude fat) | 17 | 17 | 20 | 21 |
| Non-fiber carbohydrates | 325 | 325 | 324 | 318 |
| Item | CON | PRO | ML | MSC | SEM | p-Value |
|---|---|---|---|---|---|---|
| Dry matter | 42.9 | 43.0 | 41.7 | 42.8 | 2.0 | 0.735 |
| Organic matter | 40.5 | 40.6 | 39.7 | 40.3 | 1.9 | 0.909 |
| Crude protein | 43.8 | 42.9 | 41.5 | 43.4 | 2.4 | 0.881 |
| Ash | 69.13 a | 69.30 a | 64.11 b | 69.45 a | 2.69 | 0.019 |
| Neutral detergent fiber | 20.86 | 19.93 | 19.71 | 20.00 | 3.29 | 0.277 |
| Item | CON | PRO | ML | MSC | SEM 1 | p-Value |
|---|---|---|---|---|---|---|
| pH | 6.81 | 6.82 | 6.81 | 6.8 | 0.01 | 0.368 |
| Redox potential (mV) | −197 a | −196 a | −254 b | −263 b | 6.00 | <0.001 |
| Ammonia (mmol/L) | 3.75 c | 3.73 c | 4.26 b | 4.75 a | 0.12 | <0.001 |
| Total SCFAs 2 (mmol/L) | 69.6 xy | 67.2 y | 72.4 xy | 74.3 x | 5.79 | 0.064 |
| SCFAs profile (mol/100 mol) | ||||||
| Acetate | 49.5 a | 49.8 a | 49.7 a | 48.1 b | 0.40 | <0.001 |
| Propionate | 22.9 b | 22.1 b | 22.3 b | 25.3 a | 0.50 | <0.001 |
| Butyrate | 7.20 b | 7.66 a | 7.32 ab | 7.05 b | 0.19 | 0.008 |
| Isobutyrate | 0.66 ab | 0.67 a | 0.64 b | 0.64 b | 0.01 | 0.018 |
| Valerate | 9.0 | 8.90 | 9.28 | 9.34 | 0.20 | 0.184 |
| Isovalerate | 4.27 | 4.03 | 4.08 | 4.37 | 0.36 | 0.103 |
| Caproate | 4.34 b | 4.78 a | 4.64 ab | 3.63 b | 0.25 | <0.001 |
| Heptanoate | 2.29 ab | 2.36 b | 2.53 a | 2.01 b | 0.13 | <0.001 |
| Acetate to propionate | 2.20 a | 2.29 a | 2.25 a | 1.94 b | 0.05 | <0.001 |
| Fermentation gases | ||||||
| Total fermentation gas (mL/d) | 386 | 427 | 403 | 387 | 38 | 0.685 |
| Carbon dioxide (mL/d) | 312.0 | 347.0 | 331.0 | 318.0 | 33.00 | 0.738 |
| Methane (mL/d) | 44.0 | 47.0 | 42.7 | 39.4 | 3.8 | 0.225 |
| 3 MCR (% Gross energy intake) | 0.84 xy | 0.90 x | 0.75 xy | 0.72 y | 0.07 | 0.020 |
| Gross energy intake (MJ/d) | 0.207 a | 0.207 a | 0.226 b | 0.218 c | 0.0001 | <0.001 |
| Antioxidant capacity (µg Trolox/mL) 4 | 2.58 | 2.77 | 2.33 | 2.35 | 0.31 | 0.183 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mahmood, M.; Samli, H.E.; Sener-Aydemir, A.; Sharma, S.; Zebeli, Q.; Khiaosa-ard, R. Moringa oleifera and Propolis in Cattle Nutrition: Characterization of Metabolic Activities in the Rumen In Vitro. Metabolites 2022, 12, 1237. https://doi.org/10.3390/metabo12121237
Mahmood M, Samli HE, Sener-Aydemir A, Sharma S, Zebeli Q, Khiaosa-ard R. Moringa oleifera and Propolis in Cattle Nutrition: Characterization of Metabolic Activities in the Rumen In Vitro. Metabolites. 2022; 12(12):1237. https://doi.org/10.3390/metabo12121237
Chicago/Turabian StyleMahmood, Mubarik, Hasan Ersin Samli, Arife Sener-Aydemir, Suchitra Sharma, Qendrim Zebeli, and Ratchaneewan Khiaosa-ard. 2022. "Moringa oleifera and Propolis in Cattle Nutrition: Characterization of Metabolic Activities in the Rumen In Vitro" Metabolites 12, no. 12: 1237. https://doi.org/10.3390/metabo12121237
APA StyleMahmood, M., Samli, H. E., Sener-Aydemir, A., Sharma, S., Zebeli, Q., & Khiaosa-ard, R. (2022). Moringa oleifera and Propolis in Cattle Nutrition: Characterization of Metabolic Activities in the Rumen In Vitro. Metabolites, 12(12), 1237. https://doi.org/10.3390/metabo12121237