
Multi-Omic Analysis to Characterize Metabolic Adaptation of the E. coli Lipidome in Response to Environmental Stress

 ,
,  , , and
, , and 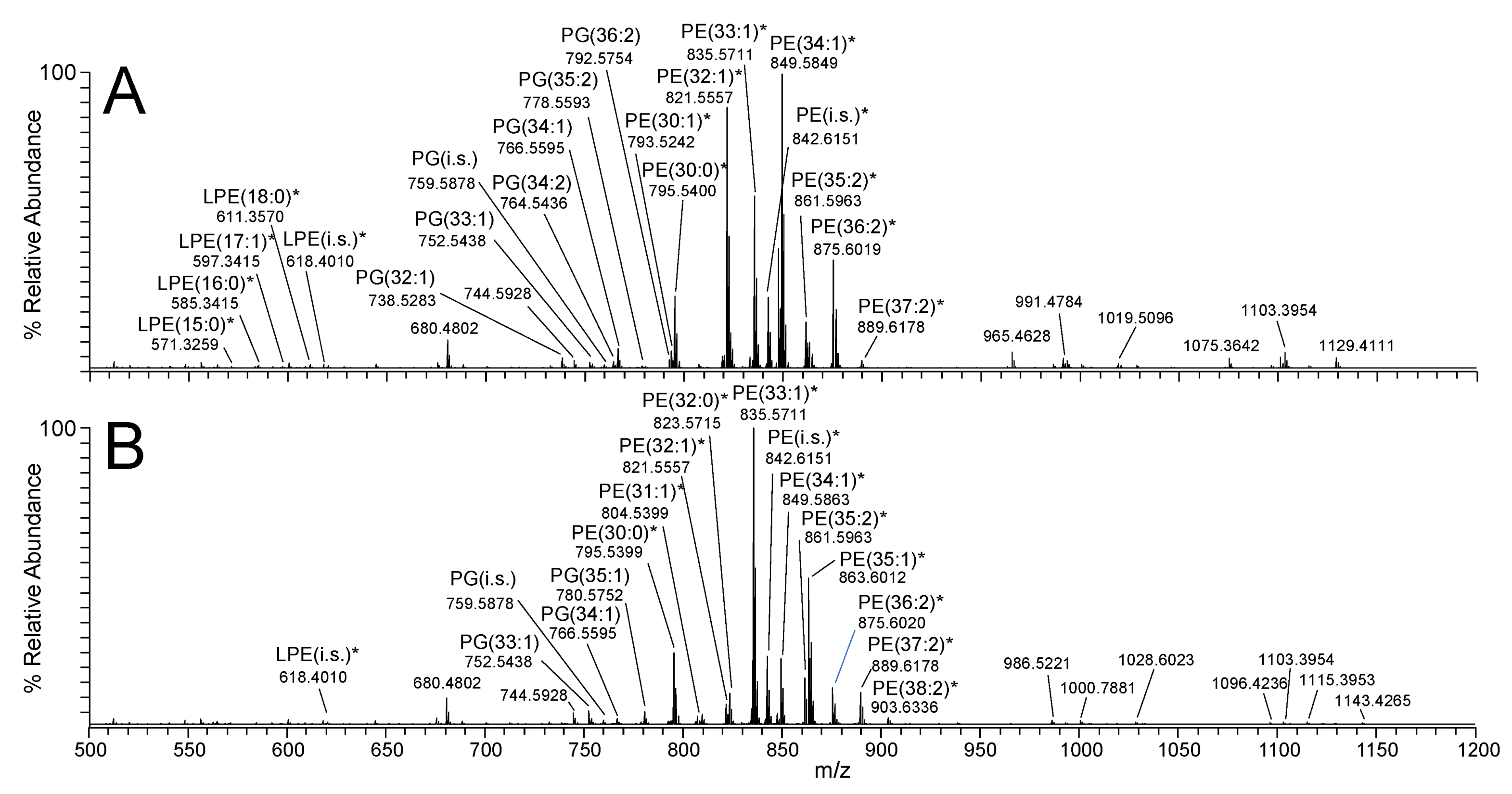{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
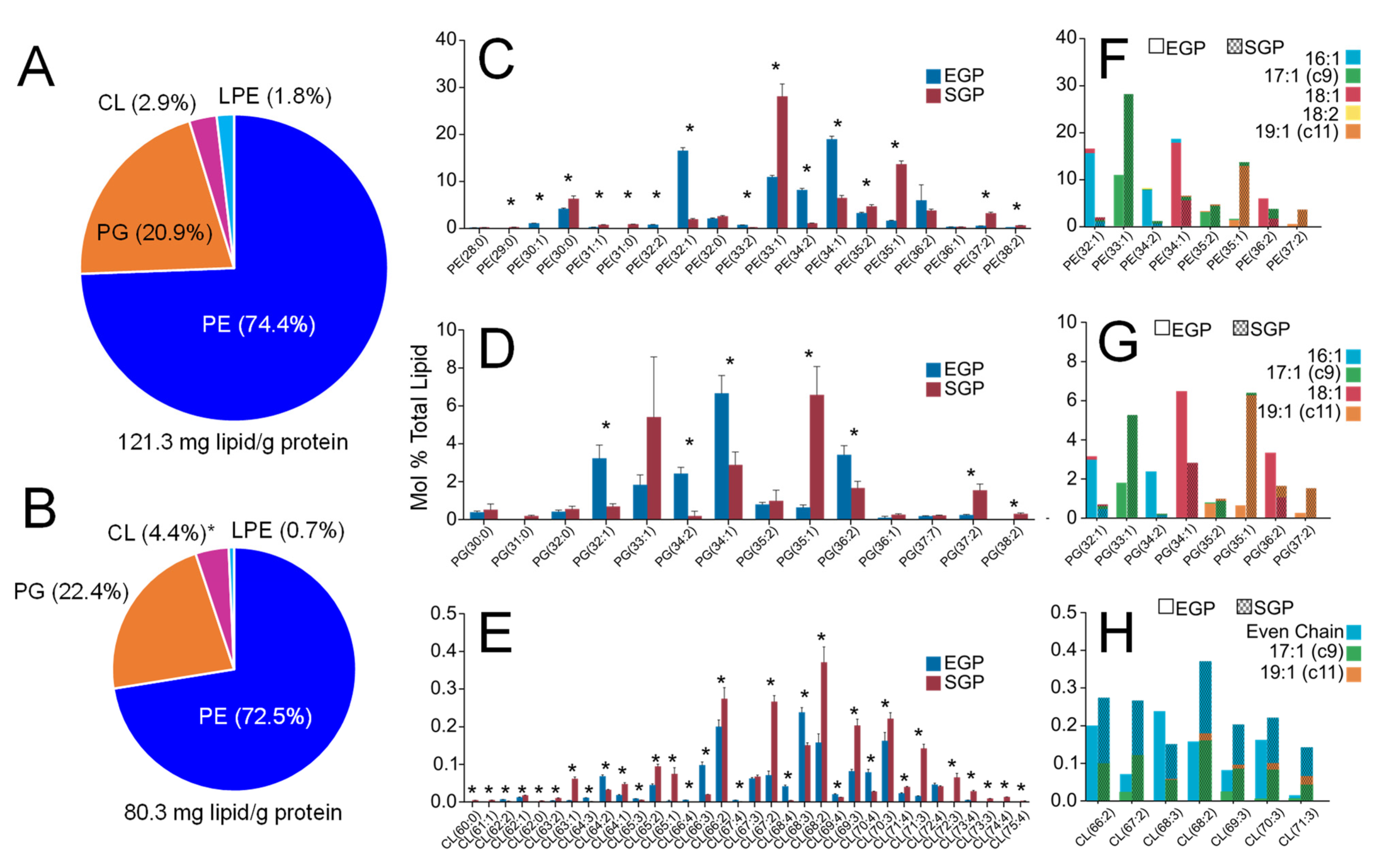
2.1. Characterisation of the E. coli Lipidome under Exponential Growth Phase (EGP) and Stationary Growth Phase (SGP) Conditions
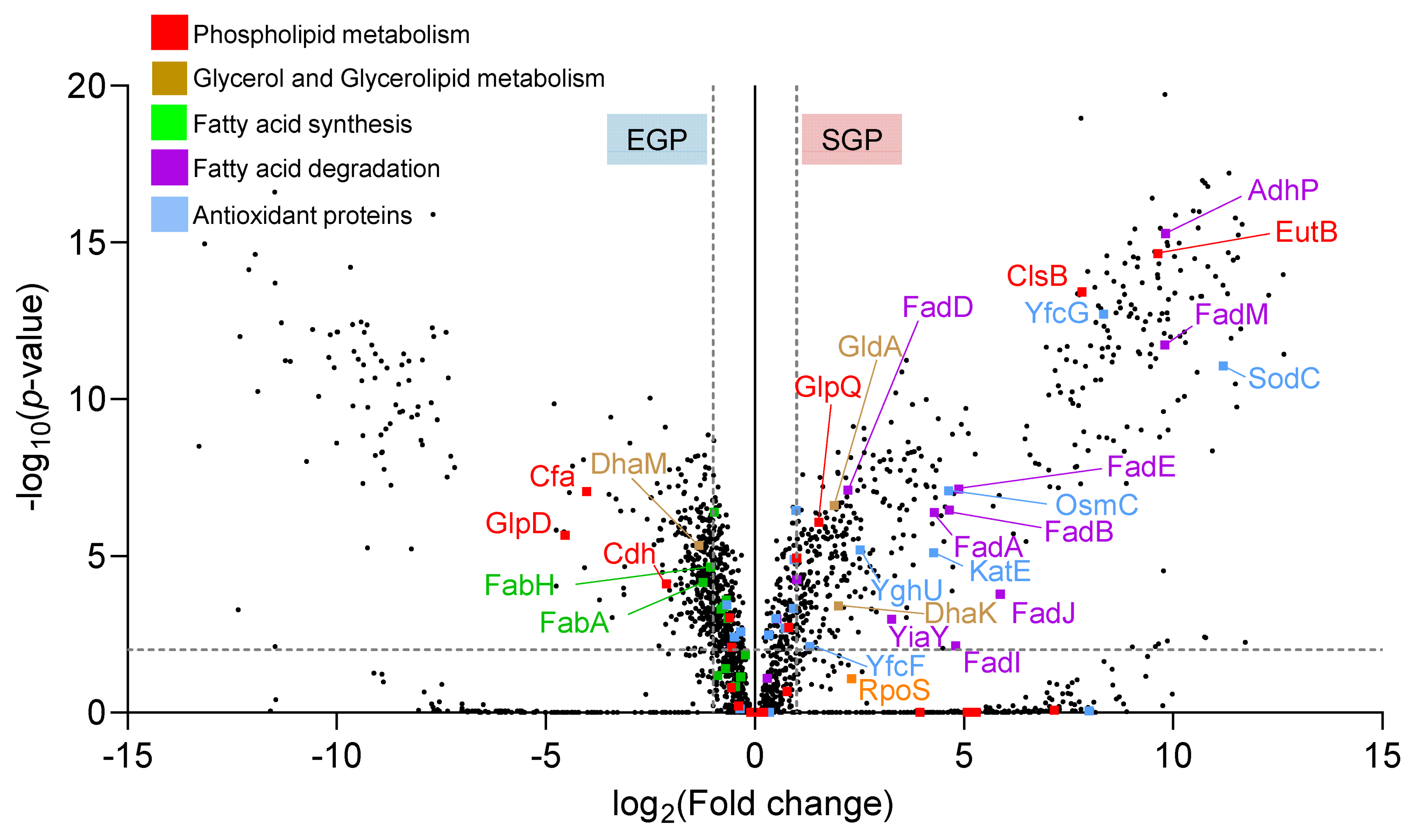
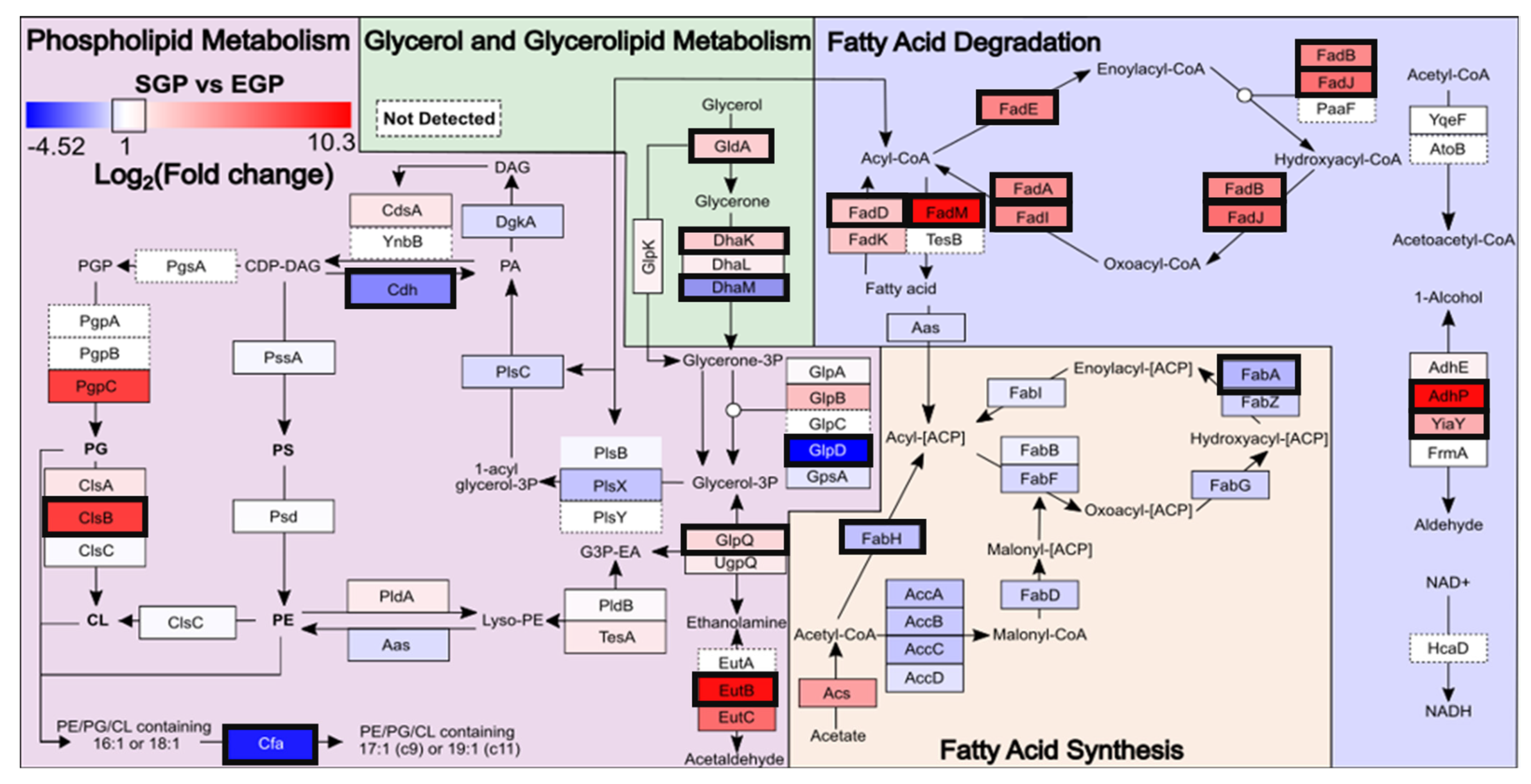
2.2. Proteomic Analysis of E. coli
3. Materials and Methods
3.1. Materials
3.2. E. coli Cell Culture
3.3. Monophasic Lipid Extraction and Derivatisation of Aminophospholipids and Plasmalogen-Containing Lipids
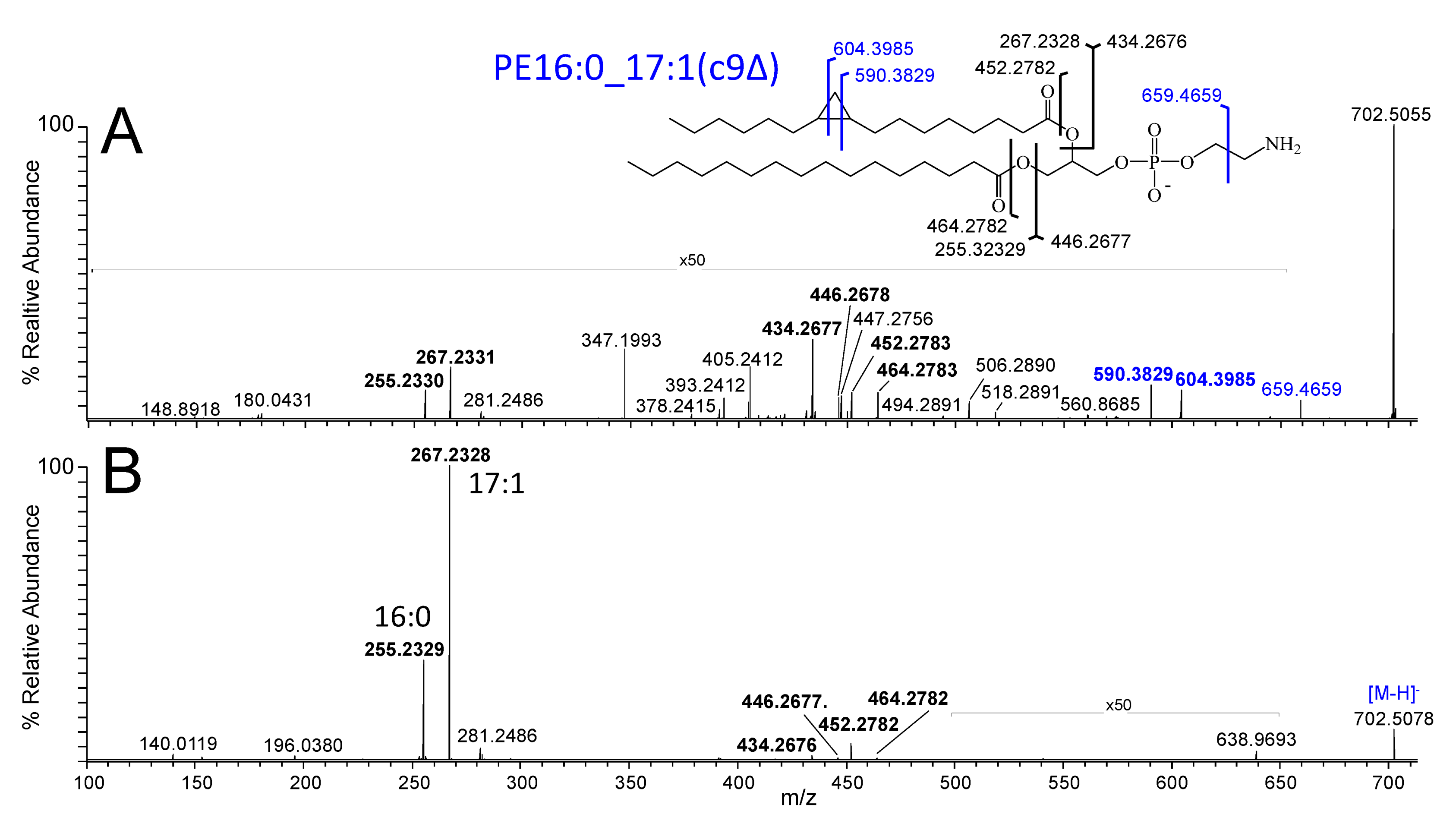
3.4. nESI-UHRAMS, CID-MS/MS and 213 nm UVPD-MS/MS Lipid Analysis
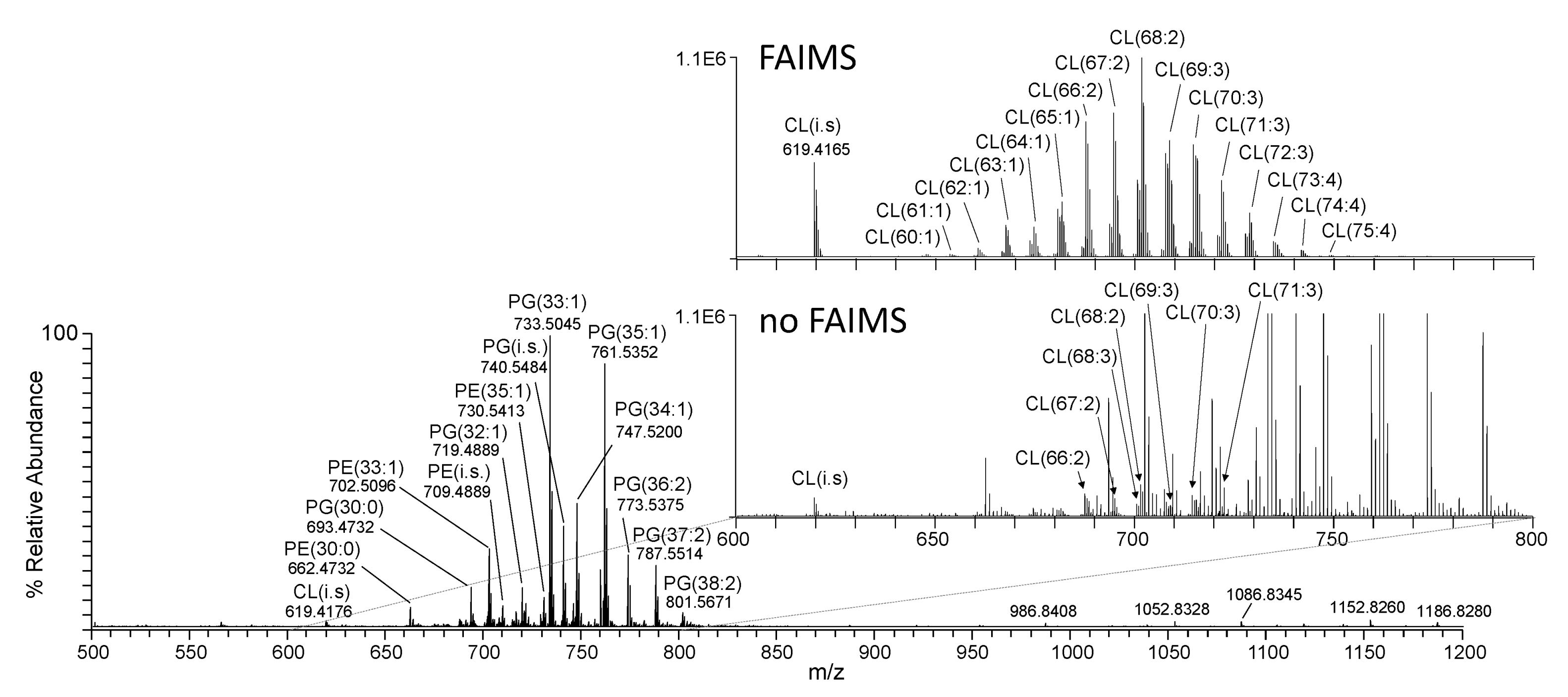
3.5. Cardiolipin Analysis Using FAIMS-MS
3.6. Lipid Identification and Quantification
3.7. Proteomic Sample Preparation
3.8. Data-Dependent Acquisition (DDA) Proteomic Analysis
3.9. Protein Identification and Label-Free Quantification
3.10. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Yano, Y.; Nakayama, A.; Ishihara, K.; Saito, H. Adaptive changes in membrane lipids of barophilic bacteria in response to changes in growth pressure. Appl. Environ. Microbiol. 1998, 64, 479–485. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hassan, N.; Anesio, A.M.; Rafiq, M.; Holtvoeth, J.; Bull, I.; Haleem, A.; Shah, A.A.; Hasan, F. Temperature driven membrane lipid adaptation in glacial psychrophilic bacteria. Front. Microbiol. 2020, 11, 824. [Google Scholar] [CrossRef] [PubMed]
- Janto, B.; Ahmed, A.; Ito, M.; Liu, J.; Hicks, D.B.; Pagni, S.; Fackelmayer, O.J.; Smith, T.A.; Earl, J.; Elbourne, L.D.; et al. Genome of alkaliphilic Bacillus pseudofirmus OF4 reveals adaptations that support the ability to grow in an external pH range from 7.5 to 11.4. Environ. Microbiol. 2011, 13, 3289–3309. [Google Scholar] [CrossRef] [Green Version]
- Kieft, T.L.; Wilch, E.; O’Connor, K.; Ringelberg, D.B.; White, D.C. Survival and phospholipid Fatty Acid profiles of surface and subsurface bacteria in natural sediment microcosms. Appl. Environ. Microbiol. 1997, 63, 1531–1542. [Google Scholar] [CrossRef] [Green Version]
- Levin, B.R.; Rozen, D.E. Non-inherited antibiotic resistance. Nat. Rev. Microbiol. 2006, 4, 556–562. [Google Scholar] [CrossRef] [PubMed]
- Gordia, S.; Gutierrez, C. Growth-phase-dependent expression of the osmotically inducible gene osmC of Escherichia coli K-12. Mol. Microbiol. 1996, 19, 729–736. [Google Scholar] [CrossRef]
- Sánchez Clemente, R.; Igeño, M.I.; Poblacion, A.; Guijo, M.; Merchán, F.; Blasco, R. Study of pH changes in media during bacterial growth of several environmental strains. Multidiscip. Digit. Publ. Inst. Proc. 2018, 2, 1297. [Google Scholar] [CrossRef] [Green Version]
- Jeucken, A.; Zhou, M.; Wösten, M.M.S.M.; Brouwers, J.F. Control of n-butanol induced lipidome adaptations in E. coli. Metabolites 2021, 11, 286. [Google Scholar] [CrossRef]
- Zhang, Y.M.; Rock, C.O. Membrane lipid homeostasis in bacteria. Nat. Rev. Microbiol. 2008, 6, 222–233. [Google Scholar] [CrossRef]
- Harayama, T.; Riezman, H. Understanding the diversity of membrane lipid composition. Nat. Rev. Mol. Cell Biol. 2018, 19, 281–296. [Google Scholar] [CrossRef]
- Rowlett, V.W.; Mallampalli, V.K.P.S.; Karlstaedt, A.; Dowhan, W.; Taegtmeyer, H.; Margolin, W.; Vitrac, H. Impact of membrane phospholipid alterations in Escherichia coli on cellular function and bacterial stress adaptation. J. Bacteriol. 2017, 19, e00849-e16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hiraoka, S.; Matsuzaki, H.; Shibuya, I. Active increase in cardiolipin synthesis in the stationary growth phase and its physiological significance in Escherichia coli. FEBS Lett. 1993, 336, 221–224. [Google Scholar] [CrossRef] [Green Version]
- Poger, D.; Pöyry, S.; Mark, A.E. Could cardiolipin protect membranes against the action of certain antimicrobial peptides? Aurein 1.2, a case study. ACS Omega 2018, 3, 16453–16464. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.Y.; Gänzle, M.G. Influence of cyclopropane fatty acids on heat, high pressure, acid and oxidative resistance in Escherichia coli. Int. J. Food Microbiol. 2016, 222, 16–22. [Google Scholar] [CrossRef] [PubMed]
- Beriashvili, D.; Taylor, R.; Kralt, B.; Abu Mazen, N.; Taylor, S.D.; Palmer, M. Mechanistic studies on the effect of membrane lipid acyl chain composition on daptomycin pore formation. Chem. Phys. Lipids 2018, 216, 73–79. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, L.H.; Zeng, X.A.; Wang, M.S.; Brennan, C.S.; Gong, D.M. Modification of membrane properties and fatty acids biosynthesis-related genes in Escherichia coli and Staphylococcus aureus: Implications for the antibacterial mechanism of naringenin. Biochim. Biophys. Acta-Biomembr. 2018, 1860, 481–490. [Google Scholar] [CrossRef]
- Lee, T.H.; Hofferek, V.; Separovic, F.; Reid, G.E.; Aguilar, M.I. The role of bacterial lipid diversity and membrane properties in modulating antimicrobial peptide activity and drug resistance. Curr. Opin. Chem. Biol. 2019, 52, 85–92. [Google Scholar] [CrossRef]
- Vranakis, I.; Goniotakis, I.; Psaroulaki, A.; Sandalakis, V.; Tselentis, Y.; Gevaert, K.; Tsiotis, G. Proteome studies of bacterial antibiotic resistance mechanisms. J. Proteom. 2014, 97, 88–99. [Google Scholar] [CrossRef]
- Miao, J.; Chen, F.; Duan, S.; Gao, X.; Liu, G.; Chen, Y.; Dixon, W.; Xiao, H.; Cao, Y. iTRAQ-based quantitative proteomic analysis of the antimicrobial mechanism of peptide F1 against Escherichia coli. J. Agric. Food Chem. 2015, 63, 7190–7197. [Google Scholar] [CrossRef]
- Zhang, D.F.; Li, H.; Lin, X.M.; Peng, X.X. Outer membrane proteomics of kanamycin-resistant Escherichia coli identified MipA as a novel antibiotic resistance-related protein. FEMS Microbiol. Lett. 2015, 362, fnv074. [Google Scholar] [CrossRef] [Green Version]
- Hines, K.M.; Waalkes, A.; Penewit, K.; Holmes, E.A.; Salipante, S.J.; Werth, B.J.; Xu, L. Characterization of the mechanisms of daptomycin resistance among gram-positive bacterial pathogens by multidimensional lipidomics. mSphere 2017, 2, e00492-e17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schmidt, R.; Yonghong, D.; Hoffmann, R. Phospholipid composition of the outer membrane of Escherichia coli influences its susceptibility against antimicrobial peptide apidaecin 1b. Diagn. Microbiol. Infect. Dis. 2018, 90, 316–323. [Google Scholar] [CrossRef] [PubMed]
- Hewelt-Belka, W.; Nakonieczna, J.; Belka, M.; Bączek, T.; Namieśnik, J.; Kot-Wasik, A. Untargeted lipidomics reveals differences in the lipid pattern among clinical isolates of Staphylococcus aureus resistant and sensitive to antibiotics. J. Proteome Res. 2016, 15, 914–922. [Google Scholar] [CrossRef] [PubMed]
- Chwastek, G.; Surma, M.A.; Rizk, S.; Grosser, D.; Lavrynenko, O.; Rucińska, M.; Jambor, H.; Sáenz, J. Principles of membrane adaptation revealed through environmentally induced bacterial lipidome remodeling. Cell Rep. 2020, 32, 108165. [Google Scholar] [CrossRef]
- Lee, T.H.; Hofferek, V.; Sani, M.A.; Separovic, F.; Reid, G.E.; Aguilar, M.I. The impact of antibacterial peptides on bacterial lipid membranes depends on stage of growth. Faraday Discuss. 2021, 232, 399–418. [Google Scholar] [CrossRef] [PubMed]
- Gidden, J.; Denson, J.; Liyanage, R.; Ivey, D.M.; Lay, J.O. Lipid compositions in Escherichia coli and Bacillus subtilis during growth as determined by MALDI-TOF and TOF/TOF mass spectrometry. Int. J. Mass Spectrom. 2009, 283, 178–184. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Low, S.X.; Aw, Z.Q.; Loo, B.Z.; Lee, K.C.; Oon, J.S.; Lee, C.H.; Ling, M.H. Viability of Escherichia coli ATCC 8739 in nutrient broth, luria-bertani broth and brain heart infusion over 11 weeks. Electron. Physician 2013, 5, 576–581. [Google Scholar]
- Xu, Y.; Zhao, Z.; Tong, W.; Ding, Y.; Liu, B.; Shi, Y.; Wang, J.; Sun, S.; Liu, M.; Wang, Y.; et al. An acid-tolerance response system protecting exponentially growing Escherichia coli. Nat. Commun. 2020, 11, 1496. [Google Scholar] [CrossRef] [Green Version]
- Romantsov, T.; Guan, Z.; Wood, J.M. Cardiolipin and the osmotic stress responses of bacteria. Biochim. Biophys. Acta 2009, 1788, 2092–2100. [Google Scholar] [CrossRef] [Green Version]
- Agrawal, A.; Rangarajan, N.; Weisshaar, J.C. Resistance of early stationary phase E. coli to membrane permeabilization by the antimicrobial peptide Cecropin, A. Biochim. Biophys. Acta Biomembr. 2019, 1861, 182990. [Google Scholar] [CrossRef]
- Jiang, X.; Duan, Y.; Zhou, B.; Guo, Q.; Wang, H.; Hang, X.; Zeng, L.; Jia, J.; Bi, H. The cyclopropane fatty acid synthase mediates antibiotic resistance and gastric colonization of helicobacter pylori. J. Bacteriol. 2019, 201, e00374-e19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jenkins, D.E.; Schultz, J.E.; Matin, A. Starvation-induced cross protection against heat or H2O2 challenge in Escherichia coli. J. Bacteriol. 1988, 170, 3910–3914. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jenkins, D.E.; Chaisson, S.A.; Matin, A. Starvation-induced cross protection against osmotic challenge in Escherichia coli. J. Bacteriol. 1990, 172, 2779–27781. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rowe, M.T.; Kirk, R.B. Cross-protection phenomenon in Escherichia coli strains harbouring cytotoxic necrotizing factors and cytolethal distending toxins. Lett. Appl. Microbiol. 2001, 32, 67–70. [Google Scholar] [CrossRef] [PubMed]
- Coman, C.; Solari, F.A.; Hentschel, A.; Sickmann, A.; Zahedi, R.P.; Ahrends, R. Simultaneous metabolite, protein, lipid extraction (SIMPLEX): A combinatorial multimolecular omics approach for systems biology. Mol. Cell Proteom. 2016, 15, 1453–1466. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakayasu, E.S.; Nicora, C.D.; Sims, A.C.; Burnum-Johnson, K.E.; Kim, Y.M.; Kyle, J.E.; Matzke, M.M.; Shukla, A.K.; Chu, R.K.; Schepmoes, A.A.; et al. MPLEx: A robust and universal protocol for single-sample integrative proteomic, metabolomic, and lipidomic analyses. mSystems 2016, 1, e00043-e16. [Google Scholar] [CrossRef] [Green Version]
- Kopczynski, D.; Coman, C.; Zahedi, R.P.; Lorenz, K.; Sickmann, A.; Ahrends, R. Multi-OMICS: A critical technical perspective on integrative lipidomics approaches. Biochim. Biophys. Acta Mol. Cell. Biol. Lipids 2017, 1862, 808–811. [Google Scholar] [CrossRef]
- Reich, S.; Nguyen, C.D.L.; Has, C.; Steltgens, S.; Soni, H.; Coman, C.; Freyberg, M.; Bichler, A.; Seifert, N.; Conrad, D.; et al. A multi-omics analysis reveals the unfolded protein response regulon and stress-induced resistance to folate-based antimetabolites. Nat. Commun. 2020, 11, 2936. [Google Scholar] [CrossRef]
- He, Y.; Rashan, E.H.; Linke, V.; Shishkova, E.; Hebert, A.S.; Jochem, A.; Westphall, M.S.; Pagliarini, D.J.; Overmyer, K.A.; Coon, J.J. Multi-omic single-shot technology for integrated proteome and lipidome analysis. Anal. Chem. 2021, 93, 4217–4222. [Google Scholar] [CrossRef]
- Sohlenkamp, C.; Geiger, O. Bacterial membrane lipids: Diversity in structures and pathways. FEMS Microbiol. Rev. 2016, 40, 133–159. [Google Scholar] [CrossRef] [Green Version]
- Jeucken, A.; Molenaar, M.R.; van de Lest, C.H.A.; Jansen, J.W.A.; Helms, J.B.; Brouwers, J.F. A comprehensive functional characterization of Escherichia coli lipid genes. Cell Rep. 2019, 27, 1597–1606. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lydic, T.A.; Busik, J.V.; Reid, G.E. A monophasic extraction strategy for the simultaneous lipidome analysis of polar and nonpolar retina lipids. J. Lipid Res. 2014, 55, 1797–1809. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hofferek, V.; Su, H.; Reid, G.E. Chemical derivatization-aided high resolution mass spectrometry for shotgun lipidome analysis. In Mass Spectrometry-Based Lipidomics: Methods and Protocols; Hsu, F.F., Ed.; Springer: New York, NY, USA, 2021; pp. 61–75. [Google Scholar]
- Fhaner, C.J.; Liu, S.; Ji, H.; Simpson, R.J.; Reid, G.E. Comprehensive lipidome profiling of isogenic primary and metastatic colon adenocarcinoma cell lines. Anal. Chem. 2012, 84, 8917–8926. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ryan, E.; Reid, G.E. Chemical derivatization and ultrahigh resolution and accurate mass spectrometry strategies for “shotgun” lipidome analysis. Acc. Chem. Res. 2016, 49, 1596–1604. [Google Scholar] [CrossRef] [PubMed]
- Feider, C.L.; Elizondo, N.; Eberlin, L.S. Ambient ionization and FAIMS mass spectrometry for enhanced imaging of multiply charged molecular ions in biological tissues. Anal. Chem. 2016, 88, 11533–11541. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, R.G.; Gao, J.; Siira, S.J.; Shearwood, A.M.; Ermer, J.A.; Hofferek, V.; Mathews, J.C.; Zheng, M.; Reid, G.E.; Rackham, O.; et al. Cardiolipin is required for membrane docking of mitochondrial ribosomes and protein synthesis. J. Cell Sci. 2020, 24, 133. [Google Scholar] [CrossRef] [PubMed]
- Hari, S.B.; Grant, R.A.; Sauer, R.T. Structural and functional analysis of E. coli cyclopropane fatty acid synthase. Structure 2018, 26, 1251–1258. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Poger, D.; Mark, A.E. A ring to rule them all: The effect of cyclopropane fatty acids on the fluidity of lipid bilayers. J. Phys. Chem. B 2015, 119, 5487–5495. [Google Scholar] [CrossRef]
- Wang, A.Y.; Cronan, J.E., Jr. The growth phase-dependent synthesis of cyclopropane fatty acids in Escherichia coli is the result of an RpoS(KatF)-dependent promoter plus enzyme instability. Mol. Microbiol. 1994, 11, 1009–1017. [Google Scholar] [CrossRef]
- Blevins, M.S.; Klein, D.R.; Brodbelt, J.S. Localization of cyclopropane modifications in bacterial lipids via 213 nm ultraviolet photodissociation mass spectrometry. Anal. Chem. 2019, 91, 6820–6828. [Google Scholar] [CrossRef]
- Macias, L.A.; Feider, C.L.; Eberlin, L.S.; Brodbelt, J.S. Hybrid 193 nm ultraviolet photodissociation mass spectrometry localizes cardiolipin unsaturations. Anal. Chem. 2019, 91, 12509–12516. [Google Scholar] [CrossRef] [PubMed]
- Klein, D.R.; Brodbelt, J.S. Structural characterization of phosphatidylcholines using 193 nm ultraviolet photodissociation mass spectrometry. Anal. Chem. 2017, 89, 1516–1522. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Williams, P.E.; Klein, D.R.; Greer, S.M.; Brodbelt, J.S. Pinpointing double bond and sn-positions in glycerophospholipids via hybrid 193 nm ultraviolet photodissociation (UVPD) mass spectrometry. J. Am. Chem Soc. 2017, 139, 15681–15690. [Google Scholar] [CrossRef] [PubMed]
- Tan, B.K.; Bogdanov, M.; Zhao, J.; Dowhan, W.; Raetz, C.R.H.; Guan, Z. Discovery of a cardiolipin synthase utilizing phosphatidylethanolamine and phosphatidylglycerol as substrates. Proc. Natl. Acad. Sci. USA 2012, 109, 16504–16509. [Google Scholar] [CrossRef] [Green Version]
- Haines, T.H.; Dencher, N.A. Cardiolipin: A proton trap for oxidative phosphorylation. FEBS Lett. 2002, 528, 35–39. [Google Scholar] [CrossRef] [Green Version]
- Weber, H.; Polen, T.; Heuveling, J.; Wendisch, V.F.; Hengge, R. Genome-wide analysis of the general stress response network in Escherichia coli: σS-dependent genes, promoters, and sigma factor selectivity. J. Bacteriol. 2005, 187, 1591–1603. [Google Scholar] [CrossRef] [Green Version]
- Chang, Y.Y.; Eichel, J.; Cronan, J.E., Jr. Metabolic instability of Escherichia coli cyclopropane fatty acid synthase is due to RpoH-dependent proteolysis. J. Bacteriol. 2000, 182, 4288–4294. [Google Scholar] [CrossRef] [Green Version]
- Gallant, J.A. Stringent control in E. coli. Annu. Rev. Genet. 1979, 13, 393–415. [Google Scholar] [CrossRef]
- Fujita, Y.; Matsuoka, H.; Hirooka, K. Regulation of fatty acid metabolism in bacteria. Mol. Microbiol. 2007, 66, 829–839. [Google Scholar] [CrossRef]
- Choi, K.H.; Heath, R.J.; Rock, C.O. Beta-ketoacyl-acyl carrier protein synthase III (FabH) is a determining factor in branched-chain fatty acid biosynthesis. J. Bacteriol. 2000, 182, 365–370. [Google Scholar] [CrossRef] [Green Version]
- Massey, L.K.; Sokatch, J.R.; Conrad, R.S. Branched-chain amino acid catabolism in bacteria. Bacteriol. Rev. 1976, 40, 42–54. [Google Scholar] [CrossRef] [PubMed]
- McDougald, D.; Gong, L.; Srinivasan, S.; Hild, E.; Thompson, L.; Takayama, K.; Rice, S.A.; Kjelleberg, S. Defences against oxidative stress during starvation in bacteria. Antonie Van Leeuwenhoek 2002, 81, 3–13. [Google Scholar] [CrossRef] [PubMed]
- Battesti, A.; Majdalani, N.; Gottesman, S. The RpoS-mediated general stress response in Escherichia coli. Annu. Rev. Microbiol. 2011, 65, 189–213. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aguilar-López, J.L.; Funes, S. Autophagy in stationary phase of growth. In Biogenesis of Fatty Acids, Lipids and Membranes; Geiger, O., Ed.; Springer International Publishing: Cham, Switzerland, 2019; pp. 849–866. [Google Scholar]
- Dukan, S.; Nyström, T. Oxidative stress defense and deterioration of growth-arrested Escherichia coli cells. J. Biol. Chem. 1999, 274, 26027–26032. [Google Scholar] [CrossRef] [Green Version]
- Mata, G.M.S.C.; Ferreira, G.M.; Spira, B. RpoS role in virulence and fitness in enteropathogenic Escherichia coli. PLoS ONE 2017, 12, e0180381. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kralj, T.; Nuske, M.; Hofferek, V.; Sani, M.-A.; Lee, T.-H.; Separovic, F.; Aguilar, M.-I.; Reid, G.E. Multi-Omic Analysis to Characterize Metabolic Adaptation of the E. coli Lipidome in Response to Environmental Stress. Metabolites 2022, 12, 171. https://doi.org/10.3390/metabo12020171
Kralj T, Nuske M, Hofferek V, Sani M-A, Lee T-H, Separovic F, Aguilar M-I, Reid GE. Multi-Omic Analysis to Characterize Metabolic Adaptation of the E. coli Lipidome in Response to Environmental Stress. Metabolites. 2022; 12(2):171. https://doi.org/10.3390/metabo12020171
Chicago/Turabian StyleKralj, Thomas, Madison Nuske, Vinzenz Hofferek, Marc-Antoine Sani, Tzong-Hsien Lee, Frances Separovic, Marie-Isabel Aguilar, and Gavin E. Reid. 2022. "Multi-Omic Analysis to Characterize Metabolic Adaptation of the E. coli Lipidome in Response to Environmental Stress" Metabolites 12, no. 2: 171. https://doi.org/10.3390/metabo12020171
APA StyleKralj, T., Nuske, M., Hofferek, V., Sani, M. -A., Lee, T. -H., Separovic, F., Aguilar, M. -I., & Reid, G. E. (2022). Multi-Omic Analysis to Characterize Metabolic Adaptation of the E. coli Lipidome in Response to Environmental Stress. Metabolites, 12(2), 171. https://doi.org/10.3390/metabo12020171