
Alkaloid Profiling, Anti-Enzymatic and Antiproliferative Activity of the Endemic Chilean Amaryllidaceae Phycella cyrtanthoides


 ,
,  and
and 
Abstract
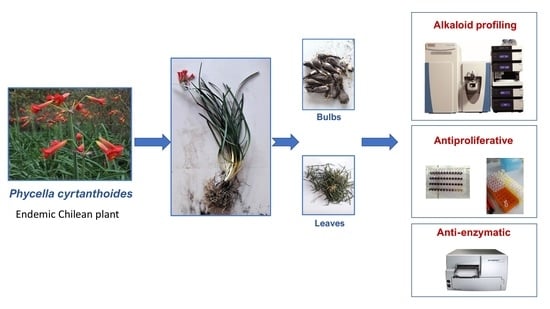
:
1. Introduction
2. Results and Discussion
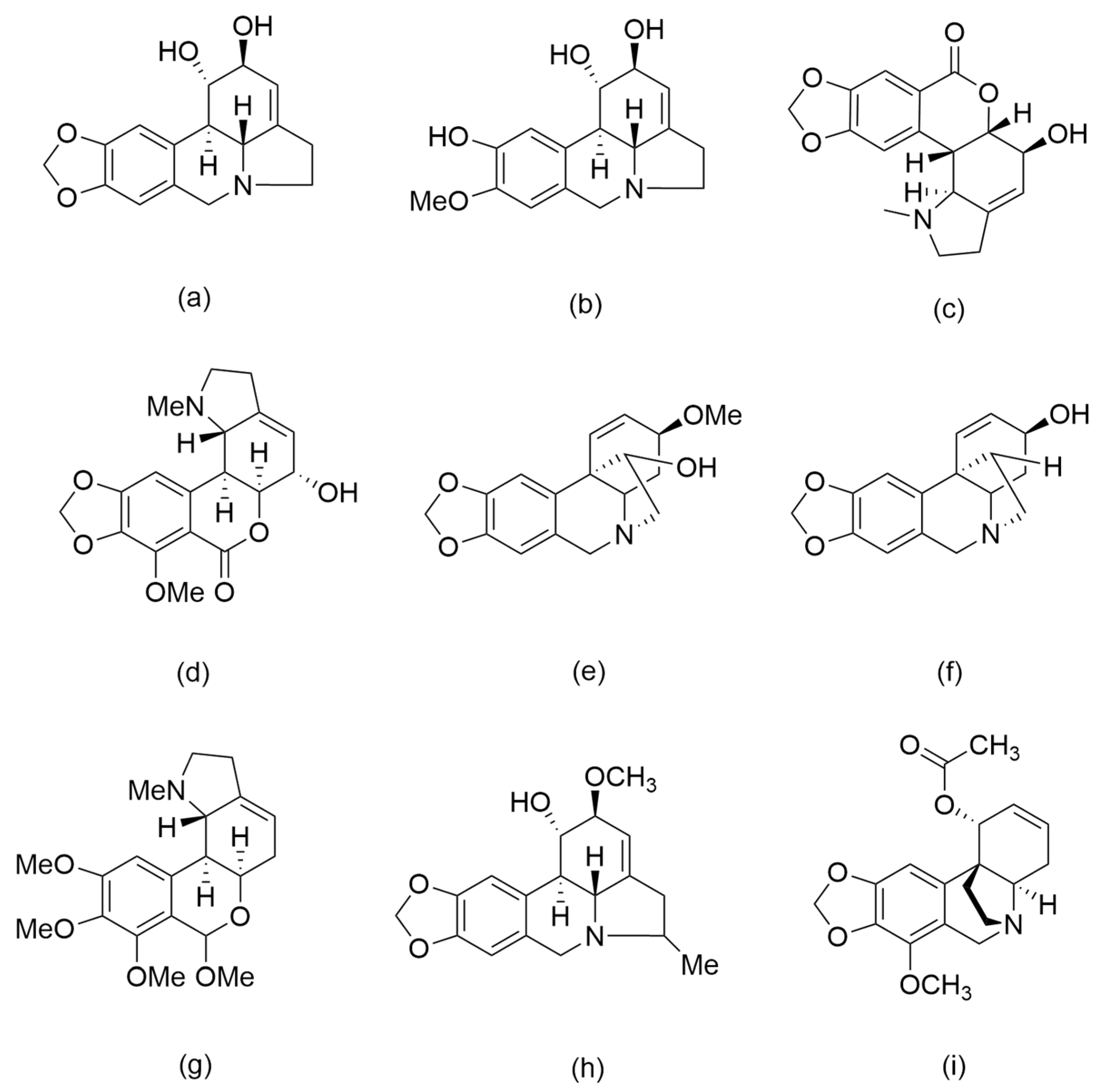
2.1. Alkaloid Profiling of Phycella Cyrtanthoides Extracts
2.2. Enzyme Inhibition Studies
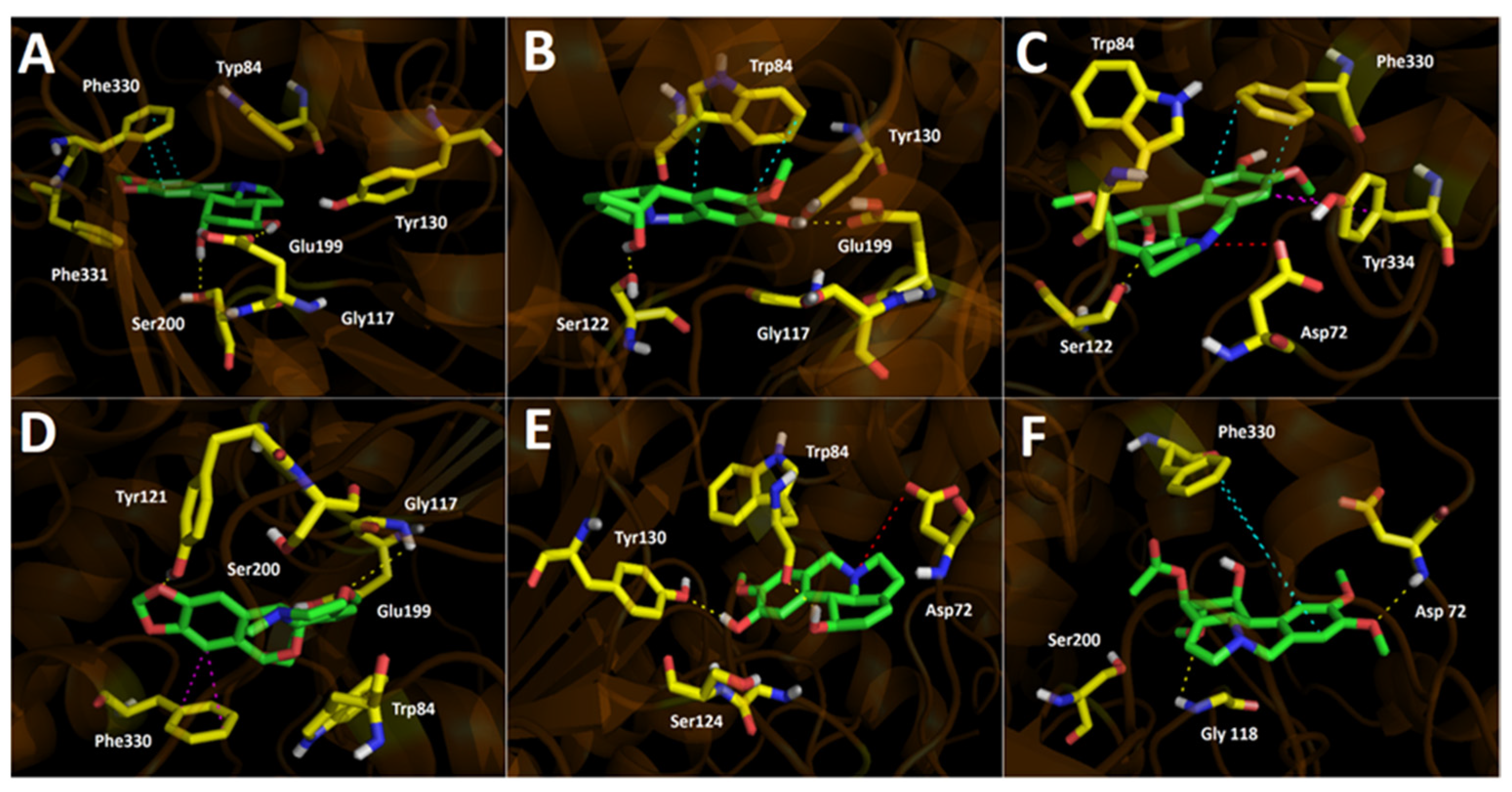
2.3. Docking Studies
2.3.1. Acetylcholinesterase (TcAChE) Docking Results
2.3.2. Butyrylcholinesterase (hBuChE) Docking Results
2.4. Antiproliferative Effects
3. Materials and Methods
3.1. Chemicals
3.2. Plant Material
3.3. Extraction
3.4. UHPLC–DAD–MS Instrument
3.5. LC Parameters and MS Parameters
3.6. Determination of Cholinesterase Inhibition
3.7. Determination of Tyrosinase Inhibition
3.8. Docking Assays
3.9. Antiproliferative Activity
3.10. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- He, M.; Qu, C.; Gao, O.; Hu, X.; Hong, X. Biological and pharmacological activities of Amaryllidaceae alkaloids. RSC Adv. 2015, 5, 16562–16574. [Google Scholar] [CrossRef]
- Riddle, J.M. Ancient and medieval chemotherapy for cancer. Isis 1985, 76, 319–330. [Google Scholar] [CrossRef] [PubMed]
- Van Goietsenoven, G.; Mathieu, V.; Lefranc, F.; Kornienko, A.; Evidente, A.; Kiss, R. Narciclasine as well as other Amaryllidaceae Isocarbostyrils are Promising GTP-ase Targeting Agents against Brain Cancers. Med. Res. Rev. 2013, 33, 439–455. [Google Scholar] [CrossRef] [PubMed]
- Heo, J.H.; Eom, B.H.; Ryu, H.W.; Kang, M.G.; Park, J.E.; Kim, D.Y.; Kim, J.H.; Park, D.; Oh, S.R.; Kim, H. Acetylcholinesterase and butyrylcholinesterase inhibitory activities of khellactone coumarin derivatives isolated from Peucedanum japonicum Thurnberg. Sci. Rep. 2020, 10, 21695. [Google Scholar] [CrossRef] [PubMed]
- Havelek, R.; Muthna, D.; Tomsik, P.; Kralovec, K.; Seifrtova, M.; Cahlikova, L.; Hostalkova, A.; Safratova, M.; Perwein, M.; Cermakova, E.; et al. Anticancer potential of Amaryllidaceae alkaloids evaluated by screening with a panel of human cells, real-time cellular analysis and Ehrlich tumor-bearing mice. Chem. Biol. Interact. 2017, 275, 121–132. [Google Scholar] [CrossRef]
- Yu, H.; Qiu, Y.; Pang, X.; Li, J.; Wu, S.; Yin, S.; Han, L.; Zhang, Y.; Jin, C.; Gao, X.; et al. Lycorine Promotes Autophagy and Apoptosis via TCRP1/Akt/mTOR Axis Inactivation in Human Hepatocellular Carcinoma. Mol. Cancer. Ther. 2017, 16, 2711–2723. [Google Scholar] [CrossRef] [Green Version]
- Cahlíková, L.; Kawano, I.; Řezáčová, M.; Blunden, G.; Hulcová, D.; Havelek, R. The Amaryllidaceae alkaloids haemanthamine, haemanthidine and their semisynthetic derivatives as potential drugs. Phytochem. Rev. 2021, 20, 303–323. [Google Scholar] [CrossRef]
- Griffin, C.; Sharda, N.; Sood, D.; Nair, J.; McNulty, J.; Pandey, S. Selective cytotoxicity of Pancratistatin-related natural Amaryllidaceae alkaloids: Evaluation of the activity of two new compounds. Cancer Cell Int. 2007, 7, 10. [Google Scholar] [CrossRef] [Green Version]
- Evidente, A.; Andolfi, A.; Abou-Donia, A.H.; Touema, S.M.; Hammoda, H.M.; Shawky, E.; Motta, A. (-)-Amarbellisine, a lycorine-type alkaloid from Amaryllis belladonna L. growing in Egypt. Phytochemistry 2004, 65, 2113–2118. [Google Scholar] [CrossRef]
- Deng, B.; Ye, L.; Yin, H.; Liu, Y.; Hu, S.; Li, B. Determination of pseudolycorine in the bulb of Lycoris radiata by capillary electrophoresis combined with online electrochemiluminescence using ultrasonic-assisted extraction. J. Chromatogr. B Analyt. Technol. Biomed. Life Sci. 2011, 879, 927–932. [Google Scholar] [CrossRef]
- Zhang, Y.; Chen, Z. Nonaqueous CE ESI-IT-MS analysis of Amaryllidaceae alkaloids. J. Sep. Sci. 2013, 36, 1078–1084. [Google Scholar] [CrossRef] [PubMed]
- Tallini, L.R.; Bastida, J.; Cortes, N.; Osorio, E.H.; Theoduloz, C.; Schmeda-Hirschmann, G. Cholinesterase inhibition activity, alkaloid profiling and molecular docking of Chilean Rhodophiala (Amaryllidaceae). Molecules 2018, 23, 1532. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Petruczynik, A.; Misiurek, J.; Tuzimski, T.; Uszyński, R.; Szymczak, G.; Chernetskyy, M.; Waksmundzka-Hajnos, M. Comparison of different HPLC systems for analysis of galantamine and lycorine in various species of Amaryllidaceae family. J. Liq. Chromatogr. Relat. Technol. 2016, 39, 574–579. [Google Scholar] [CrossRef]
- Ortiz, J.E.; Pigni, N.B.; Andujar, S.A.; Roitman, G.; Suvire, F.D.; Enriz, R.D.; Tapia, A.; Bastida, J.; Feresin, G.E. Alkaloids from Hippeastrum argentinum and Their Cholinesterase-Inhibitory Activities: An in Vitro and in Silico Study. J. Nat. Prod. 2016, 79, 1241–1248. [Google Scholar] [CrossRef]
- Karakoyun, Ç.; Bozkurt, B.; Çoban, G.; Masi, M.; Cimmino, A.; Evidente, A.; Unver Somer, N. A comprehensive study on Narcissus tazetta subsp. tazetta L.: Chemo-profiling, isolation, anticholinesterase activity and molecular docking of Amaryllidaceae alkaloids. S. Afr. J. Bot. 2020, 130, 148–154. [Google Scholar] [CrossRef]
- Li, A.; Du, Z.; Liao, M.; Feng, Y.; Ruan, H.; Jiang, H. Discovery and characterisation of lycorine-type alkaloids in Lycoris spp. (Amaryllidaceae) using UHPLC-QTOF-MS. Phytochem. Anal. 2019, 30, 268–277. [Google Scholar] [CrossRef]
- Jin, Z. Amaryllidaceae and Sceletium alkaloids. Nat. Prod. Rep. 2013, 30, 849–868. [Google Scholar] [CrossRef]
- Baeza, C.; Ruiz, E.; Almendras, F.; Peñailillo, P. Estudio comparativo del cariotipo en especies de Miltinea Ravenna, Phycella Lindl. y Rhodophiala C. Presl (Amaryllidaceae) de Chile. Rev. Fac. Cienc. Agrar. 2012, 44, 193–205. [Google Scholar]
- Cisternas, M.A.; Araneda, L.; García, N.; Baeza, C.M. Estudios cariotípicos en el género chileno Placea (Amaryllidaceae). Gayana Bot. 2010, 67, 198–205. [Google Scholar] [CrossRef] [Green Version]
- Urbina-Casanova, R.; Saldivia, P.; Scherson, R.A. Consideraciones sobre la sistemática de las familias y los géneros de plantas vasculares endémicos de Chile. Gayana Bot. 2015, 72, 272–295. [Google Scholar] [CrossRef] [Green Version]
- García, N.; Meerow, A.W.; Arroyo-Leuenberger, S.; Oliveira, R.S.; Dutilh, J.H.; Soltis, P.S.; Judd, W.S. Generic classification of Amaryllidaceae tribe Hippeastreae. TAXON 2019, 68, 481–498. [Google Scholar] [CrossRef]
- Trujillo-Chacón, L.M.; Alarcón-Enos, J.E.; Céspedes-Acuña, C.L.; Bustamante, L.; Baeza, M.; López, M.G.; Fernández-Mendívil, C.; Cabezas, F.; Pastene-Navarrete, E.R. Neuroprotective activity of isoquinoline alkaloids from of Chilean Amaryllidaceae plants against oxidative stress-induced cytotoxicity on human neuroblastoma SH-SY5Y cells and mouse hippocampal slice culture. Food Chem. Toxicol. 2019, 132, 110665. [Google Scholar] [CrossRef] [PubMed]
- Ortiz, J.E.; Berkov, S.; Pigni, N.B.; Theoduloz, C.; Roitman, G.; Tapia, A.; Bastida, J.; Feresin, G.E. Wild argentinian Amaryllidaceae, a new renewable source of the acetylcholinesterase inhibitor galanthamine and other alkaloids. Molecules 2012, 17, 13473–13482. [Google Scholar] [CrossRef] [Green Version]
- Shitara, N.; Hirasawa, Y.; Hasumi, S.; Sasaki, T.; Matsumoto, M.; Wong, C.P.; Kaneda, T.; Asakawa, Y.; Morita, H. Four new Amaryllidaceae alkaloids from Zephyranthes candida. J. Nat. Med. 2014, 68, 610–614. [Google Scholar] [CrossRef]
- Cortes, N.; Sierra, K.; Alzate, F.; Osorio, E.H.; Osorio, E. Alkaloids of Amaryllidaceae as Inhibitors of Cholinesterases (AChEs and BChEs): An Integrated Bioguided Study. Phytochem. Anal. 2018, 29, 217–227. [Google Scholar] [CrossRef] [PubMed]
- Katoch, D.; Kumar, D.; Padwad, Y.S.; Singh, B.; Sharma, U. Pseudolycorine N-oxide, a new N-oxide from Narcissus tazetta. Nat. Prod. Res. 2019, 34, 2051–2058. [Google Scholar] [CrossRef]
- Chen, G.-L.; Tian, Y.-Q.; Wu, J.-L.; Li, N.; Guo, M.-Q. Antiproliferative activities of Amaryllidaceae alkaloids from Lycoris radiata targeting DNA topoisomerase I. Sci. Rep. 2016, 6, 38284. [Google Scholar] [CrossRef] [Green Version]
- De Andrade, J.P.; Guo, Y.; Font-Bardia, M.; Calvet, T.; Dutilh, J.; Viladomat, F.; Codina, C.; Nair, J.J.; Zuanazzi, J.A.S.; Bastida, J. Crinine-type alkaloids from Hippeastrum aulicum and H. calyptratum. Phytochemistry 2014, 103, 188–195. [Google Scholar] [CrossRef]
- Zhang, X.; Huang, H.; Liang, X.; Huang, H.; Dai, W.; Shen, Y.; Yan, S.; Zhang, W. Analysis of Amaryllidaceae alkaloids from Crinum by high-performance liquid chromatography coupled with electrospray ionization tandem mass spectrometry. Rapid Commun. Mass Spectrom. 2009, 23, 2903–2916. [Google Scholar] [CrossRef]
- Giordani, R.B.; Vieira, P.D.B.; Weizenmann, M.; Rosemberg, D.B.; Souza, A.P.; Bonorino, C.; De Carli, G.A.; Bogo, M.R.; Zuanazzi, J.A.; Tasca, T. Candimine-induced cell death of the amitochondriate parasite Trichomonas vaginalis. J. Nat. Prod. 2010, 73, 2019–2023. [Google Scholar] [CrossRef]
- Cortes, N.; Castañeda, C.; Osorio, E.H.; Cardona-Gomez, G.P.; Osorio, E. Amaryllidaceae alkaloids as agents with protective effects against oxidative neural cell injury. Life Sci. 2018, 203, 54–65. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.D.; Zhang, Y.; He, H.P.; Li, S.F.; Tang, G.H.; Chen, D.Z.; Cao, M.M.; Di, Y.T.; Hao, X.J. A new Amaryllidaceae alkaloid from the bulbs of Lycoris radiata. Chin. J. Nat. Med. 2013, 11, 406–410. [Google Scholar] [CrossRef] [PubMed]
- Masi, M.; Van slambrouck, S.; Gunawardana, S.; van Rensburg, M.J.; James, P.C.; Mochel, J.G.; Heliso, P.S.; Albalawi, A.S.; Cimmino, A.; van Otterlo, W.A.L.; et al. Alkaloids isolated from Haemanthus humilis Jacq., an indigenous South African Amaryllidaceae: Anticancer activity of coccinine and montanine. S. Afr. J. Bot. 2019, 126, 277–281. [Google Scholar] [CrossRef]
- Tarakemeh, A.; Azizi, M.; Rowshan, V.; Salehi, H.; Spina, R.; Dupire, F.; Arouie, H.; Laurain-Mattar, D. Screening of Amaryllidaceae alkaloids in bulbs and tissue cultures of Narcissus papyraceus and four varieties of N. tazetta. J. Pharm. Biomed. Anal. 2019, 172, 230–237. [Google Scholar] [CrossRef]
- Quirion, J.C.; Husson, H.P.; Weniger, B.; Jimenez, F.; Zanoni, T.A. (-)-3-O-Acetylnarcissidine, a New Alkaloid from Hippeastrum puniceum. J. Nat. Prod. 2004, 54, 1112–1114. [Google Scholar] [CrossRef]
- De Andrade, J.P.; Pigni, N.B.; Torras-Claveria, L.; Berkov, S.; Codina, C.; Viladomat, F.; Bastida, J. Bioactive alkaloid extracts from Narcissus broussonetii: Mass spectral studies. J. Pharm. Biomed. Anal. 2012, 70, 13–25. [Google Scholar] [CrossRef]
- Pigni, N.B.; Ríos-Ruiz, S.; Martínez-Francés, V.; Nair, J.J.; Viladomat, F.; Codina, C.; Bastida, J. Alkaloids from Narcissus serotinus. J. Nat. Prod. 2012, 75, 1643–1647. [Google Scholar] [CrossRef]
- Masi, M.; Frolova, L.V.; Yu, X.; Mathieu, V.; Cimmino, A.; De Carvalho, A.; Kiss, R.; Rogelj, S.; Pertsemlidis, A.; Kornienko, A.; et al. Jonquailine, a new pretazettine-type alkaloid isolated from Narcissus jonquilla quail, with activity against drug-resistant cancer. Fitoterapia 2015, 102, 41–48. [Google Scholar] [CrossRef] [Green Version]
- Kaya, G.I.; Unver, N.; Gözler, B.; Bastida, J. (-)-Capnoidine and (+)-bulbocapnine from an Amaryllidaceae species, Galanthus nivalis subsp. cilicicus. Biochem. Syst. Ecol. 2004, 32, 1059–1062. [Google Scholar] [CrossRef]
- Tallini, L.R.; de Andrade, J.P.; Kaiser, M.; Viladomat, F.; Nair, J.J.; Zuanazzi, J.A.S.; Bastida, J. Alkaloid Constituents of the Amaryllidaceae Plant Amaryllis belladonna L. Molecules 2017, 22, 1437. [Google Scholar] [CrossRef] [Green Version]
- Al Mamun, A.; Maříková, J.; Hulcová, D.; Janoušek, J.; Šafratová, M.; Nováková, L.; Kučera, T.; Hrabinová, M.; Kuneš, J.; Korábečný, J.; et al. Amaryllidaceae Alkaloids of Belladine-Type from Narcissus pseudonarcissus cv. Carlton as New Selective Inhibitors of Butyrylcholinesterase. Biomolecules 2020, 10, 800. [Google Scholar] [CrossRef] [PubMed]
- Van Rensburg, E.; Zietsman, P.C.; Bonnet, S.L.; Wilhelm, A. Alkaloids from the Bulbs of Boophone disticha. Nat. Prod. Commun. 2017, 12, 1431–1433. [Google Scholar] [CrossRef] [Green Version]
- Ghosal, S.; Rao, P.H.; Saini, K.S. Natural Occurrence of 11-O-Acetylambelline and 11-O-Acetyl-l,2-β-epoxyambelline in Crinum latifolium: Immuno-regulant Alkaloids. Pharm. Res. 1985, 2, 251–252. [Google Scholar] [CrossRef]
- Cespedes, C.L.; Balbontin, C.; Avila, J.G.; Dominguez, M.; Alarcon, J.; Paz, C.; Burgos, V.; Ortiz, L.; Peñaloza-Castro, I.; Seigler, D.S.; et al. Inhibition on cholinesterase and tyrosinase by alkaloids and phenolics from Aristotelia chilensis leaves. Food Chem. Toxicol. 2017, 109, 984–995. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Galleguillos, C.; Quesada-Romero, L.; Puerta, A.; Padrón, J.M.; Souza, E.; Romero-Parra, J.; Simirgiotis, M.J. Uhplc-ms chemical fingerprinting and antioxidant, antiproliferative, and enzyme inhibition potential of Gaultheria pumila berries. Metabolites 2021, 11, 523. [Google Scholar] [CrossRef]
- Abebe, B.; Tadesse, S.; Hymete, A.; Bisrat, D. Antiproliferative Effects of Alkaloids from the Bulbs of Crinum abyscinicum Hochst. ExA. Rich. Evid. Based Complement. Altern. Med. 2020, 2020, 2529730. [Google Scholar] [CrossRef]
- Lamoral-Theys, D.; Andolfi, A.; Van Goietsenoven, G.; Cimmino, A.; Le Calvé, B.; Wauthoz, N.; Mégalizzi, V.; Gras, T.; Bruyère, C.; Dubois, J.; et al. Lycorine, the Main Phenanthridine Amaryllidaceae Alkaloid, Exhibits Significant Antitumor Activity in Cancer Cells That Display Resistance to Proapoptotic Stimuli: An Investigation of Structure−Activity Relationship and Mechanistic Insight. J. Med. Chem. 2009, 52, 6244–6256. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carvalho, K.R.; Silva, A.B.; Torres, M.C.M.; Pinto, F.C.L.; Guimarães, L.A.; Rocha, D.D.; Silveira, E.R.; Costa-Lotufo, L.V.; Braz-Filho, R.; Pessoa, O.D.L. Cytotoxic Alkaloids from Hippeastrum solandriflorum Lindl. J. Braz. Chem. Soc. 2015, 26, 1976–1980. [Google Scholar] [CrossRef]
- Reis, A.; Magne, K.; Massot, S.; Tallini, L.R.; Scopel, M.; Bastida, J.; Ratet, P.; Zuanazzi, J.A.S. Amaryllidaceae alkaloids: Identification and partial characterization of montanine production in Rhodophiala bifida plant. Sci. Rep. 2019, 9, 8471. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barrientos, R.; Fernández-Galleguillos, C.; Pastene, E.; Simirgiotis, M.; Romero-Parra, J.; Ahmed, S.; Echeverría, J. Metabolomic Analysis, Fast Isolation of Phenolic Compounds, and Evaluation of Biological Activities of the Bark from Weinmannia trichosperma Cav. (Cunoniaceae). Front. Pharmacol. 2020, 11, 780. [Google Scholar] [CrossRef] [PubMed]
- Larrazábal-Fuentes, M.J.; Fernández-Galleguillos, C.; Palma-Ramírez, J.; Romero-Parra, J.; Sepúlveda, K.; Galetovic, A.; González, J.; Paredes, A.; Bórquez, J.; Simirgiotis, M.J.; et al. Chemical Profiling, Antioxidant, Anticholinesterase, and Antiprotozoal Potentials of Artemisia copa Phil. (Asteraceae). Front. Pharmacol. 2020, 11, 1911. [Google Scholar] [CrossRef]
- Barrientos, R.E.; Ahmed, S.; Cortés, C.; Fernández-Galleguillos, C.; Romero-Parra, J.; Simirgiotis, M.J.; Echeverría, J. Chemical Fingerprinting and Biological Evaluation of the Endemic Chilean Fruit Greigia sphacelata (Ruiz and Pav.) Regel (Bromeliaceae) by UHPLC-PDA-Orbitrap-Mass Spectrometry. Molecules 2020, 25, 3750. [Google Scholar] [CrossRef]
- Petersson, G.A.; Bennett, A.; Tensfeldt, T.G.; Al-Laham, M.A.; Shirley, W.A.; Mantzaris, J. A complete basis set model chemistry. I. The total energies of closed-shell atoms and hydrides of the first-row elements. J. Chem. Phys. 1988, 89, 2193–2218. [Google Scholar] [CrossRef]
- McLean, A.D.; Chandler, G.S. Contracted Gaussian basis sets for molecular calculations. I. Second row atoms, Z = 11–18. J. Chem. Phys. 2008, 72, 5639. [Google Scholar] [CrossRef]
- Greenblatt, H.M.; Kryger, G.; Lewis, T.; Silman, I.; Sussman, J.L. Structure of acetylcholinesterase complexed with (−)-galanthamine at 2.3 Å resolution. FEBS Lett. 1999, 463, 321–326. [Google Scholar] [CrossRef] [Green Version]
- Nachon, F.; Carletti, E.; Ronco, C.; Trovaslet, M.; Nicolet, Y.; Jean, L.; Renard, P.Y. Crystal structures of human cholinesterases in complex with huprine W and tacrine: Elements of specificity for anti-Alzheimer’s drugs targeting acetyl- and butyryl-cholinesterase. Biochem. J. 2013, 453, 393–399. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berman, H.M.; Westbrook, J.; Feng, Z.; Gilliland, G.; Bhat, T.N.; Weissig, H.; Shindyalov, I.N.; Bourne, P.E. The Protein Data Bank. Nucleic Acids Res. 2000, 28, 235–242. [Google Scholar] [CrossRef] [PubMed] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Peak | UV Max | Tentative Identification Name (AA-Type) | Elemental Composition [M+H] + | Rt | Theoretical Mass (m/z) | Measured Mass (m/z) | Accuracy (ppm) | MSn Ions (ppm) | Organs |
|---|---|---|---|---|---|---|---|---|---|
| 1 | 236–281 | 2-Hydroxyalbomaculine (homolycorine-type) | C19H24O6N+ | 0.86 | 362.15981 | 362.13348 | −72.719 | 274.14774, 251.09735, 214.06267, 199.07582, 165.07063, 121.06544, 107.04959 | L |
| 2 | 231–248-271–307 | Tazettine isomer (tazettine-type) | C18H22O5N+ | 1.37 | 332.14925 | 332.15756 | 25.020 | 316.12589, 274.14835, 247.12236, 228.14095, 181.06543, 152.06271, 115.05478, 107.04965 | L |
| 3 | 233–247-306 | Powelline (crinine-type) | C17H20O4N+ | 2.06 | 302.13868 | 302.14523 | 21.663 | 284.13364, 274.14798, 251.09767, 228.06763, 181.06557, 165.07068, 127.05473, 115.05476, | L |
| 4 | 236–286 | Lycorine * (lycorine-type) | C16H18O4N+ | 3.07 | 288.12303 | 288.12869 | 19.628 | 286.11295, 239.05540, 194.11844, 166.12335, 147.04440, 119.04981, 103.05488 | B; L |
| 5 | 235–283 | Pseudolycorine (lycorine-type) | C16H20O4N+ | 3.11 | 290.13923 | 290.14432 | 19.423 | 244.09990, 214.08778, 177.26830, 153.07031, 152.06248, 147.04427, 119.04977, 112.91312 | B; L |
| 6 | 233–278 | Hippeastrine isomers (homolycorine-type) | C17H18O5N+ | 3.25 | 316.11850 | 316.12579 | 25.689 | 298.11389, 290.14404, 280.10153, 274.14795, 191.03445, 166.12321, 121.06518 | B; L |
| 7 | 231–276 | Hippeastrine isomers (homolycorine-type) | C17H18O5N+ | 4.54 | 316.11850 | 316.12579 | 24.803 | 290.14407, 274.14798, 191.03456, 166.12329, 121.06531 | L |
| 8 | 232–278 | Hippeastrine isomers (homolycorine-type) | C17H18O5N+ | 4.59 | 316.11850 | 316.12616 | 25.973 | 291.14764, 274.14828, 191.03468, 247.12253, 166.12341, 121.06541 | B |
| 9 | 233–289 | Pluviine (lycorine- type) | C17H22O3N+ | 5.35 | 288.15942 | 288.12869 | 19.628 | 275.25861, 270.11700, 240.06802, 194.09659, 165.07004, 147.04448, 119.04977 | B |
| 10 | 232–289 | Dihydrolycorine (lycorine- type) | C16H20O4N+ | 6.59 | 290.13923 | 290.14432 | 19.423 | 272.13245, 257.15308, 220.12132, 167.11861, 152.06258, 149.06068 | B |
| 11 | 233–290 | 3-O-methyl-epimacowine (crinine-type) | C16H18O4N+ | 6.73 | 288.12358 | 288.12872 | 19.732 | 193.65796, 179.76768, 153.07051, 128.55969, 115.05492, 109.22836 | B |
| 12 | 231–277-310 | 11-O-methylcrinamine (crinine-type) | C18H22O4N+ | 6.84 | 316.15433 | 316.12589 | −89.979 | 631.24774 (2M+), 274.14807, 228.14088, 182.11839, 121.06535 | L |
| 13 | 231–277 | 3-Hydroxydihydrocaranine (lycorine- type) | C16H20O4N+ | 7.74 | 290.13868 | 290.14420 | 19.940 | 268.67581, 239.88618, 167.34416, 137.10796, 111.88570, 107.48717 | B |
| 14 | 232–278 | epi-Zephyranthine isomers (lycorine- type) | C16H20O4N+ | 8.34 | 290.13868 | 290.14413 | 18.768 | 272.13232, 262.11163, 244.09996, 214.08743, 181.06534, 169.06546, 147.04451, 120.08131, 118.06568 | L |
| 15 | 232–278 | epi-Zephyranthine isomers (lycorine- type) | C16H20O4N+ | 8.81 | 290.13868 | 290.14435 | 19.526 | 272.13245, 268.74606, 181.17052, 170.64424, 148.16725, 129.39676, 118.06547 | B |
| 16 | 232–275 | Hippeastrine isomers (homolycorine-type) | C17H18O5N+ | 9.67 | 316.11850 | 316.12579 | 24.80 | 298.11267, 290.14447, 274.14795, 191.03447, 166.12320, 121.06516 | L |
| 17 | 232–275 | Hippeastrine isomers (homolycorine-type) | C17H18O5N+ | 10.09 | 316.11850 | 316.12610 | −22.476 | 288.12851, 274.14813, 191.03467, 166.12337, 137.10789, 124.07613 | B |
| 18 | 235–282-314 | Candimine (homolycorine-type) | C18H20O6N+ | 10.75 | 346.12906 | 346.13831 | 28.301 | 316.12576, 288.12836, 274.14810, 228.14084, 155.15477, 138.05533, 121.06532 | B; L |
| 19 | 234–385 | 8-O-demethylmaritidine (haemanthamine-type) | C16H20O3N+ | 11.39 | 274.14377 | 274.14804 | 15.576 | 267.06967, 223.07715, 191.03459, 177.01894, 149.02383, 121.06531, 107.04959 | L |
| 20 | 232–273 | Haemanthamine (haemanthamine-type) | C17H20O4N+ | 11.78 | 302.13923 | 302.14548 | 22.490 | 289.13168, 272.13251, 228.14088, 183.57773, 180.10258, 167.11855, 161.10791, 144.08138 | B; L |
| 21 | 235–282 | 5-Methyl-epimethylpseudolycorine (lycorine-type) | C18H24O4N+ | 11.89 | 318.17053 | 318.17819 | 25.788 | 287.12823, 162.06857, 147.04459, 125.98681, 115.05488, 103.05471 | B |
| 22 | 241–325 | 2α-Methoxy-6-O-ethyloduline (homolycorine-type) | C20H26O5N+ | 12.16 | 360.18110 | 360.19092 | 28.792 | 330.14169, 274.14807, 270.11676, 228.14082, 153.10274, 151.07596, 121.06521 | L |
| 23 | 240–291 | Vittatine (haemanthamine- type) | C16H18O3N+ | 12.43 | 272.12867 | 272.13257 | 16.242 | 268.10175, 247.12251, 199.21875, 180.10249, 167.99812, 153.13918, 121.06526, 115.05503 | B; L |
| 24 | 233–288 | 10-Norpluviine (lycorine-tipe) | C16H20O3N+ | 12.63 | 274.14432 | 274.14813 | 15.904 | 274.14813, 256.13666, 228.14076, 175.03946, 147.04443, 121.06533, 118.06563, 102.03407 | L |
| 25 | 236–287 | Kirkine (lycorine-tipe) | C16H20O3N+ | 12.73 | 274.14432 | 274.14822 | 16.232 | 270.11679, 256.13672, 231.15173, 228.07106, 197.16562, 175.03972, 120.08125, 118.06559 | B |
| 26 | 232–272 | Albomaculine (homolycorine-type) | C19H24O5N+ | 12.87 | 346.16490 | 346.17456 | 27.994 | 320.15652, 304.16083, 274.14795, 193.05043, 180.10242, 178.06317, 152.06268, 103.05467 | L |
| 27 | 232–256-309 | 3-Epimacronine (tazettine-type) | C18H20O5N+ | 13.05 | 330.13415 | 330.14191 | 25.173 | 326.09451, 316.12592, 247.12227, 231.15181, 202.13469, 167.15497, 144.08133, 111.09221 | B |
| 28 | 234–252 | 10-O-methylpseudolycorine (lycorine-type) | C17H22O4N+ | 13.12 | 304.15488 | 304.16098 | 21.848 | 304.16074, 276.12741, 258.11655, 193.05020, 178.06328, 165.07066, 147.04482, 125.08409, 118.06564 | L |
| 29 | 232–271 | Aknadicine | C19H24O5N+ | 13.29 | 346.16545 | 346.17526 | 29.929 | 316.12640, 304.16125, 193.05049, 178.06313, 125.08411, 121.06541, 110.06061 | L |
| 30 | 248–271 | 3-O-Acetylnarcissidine (lycorine-type) | C20H26O6N+ | 13.43 | 376.17601 | 376.18796 | 33.217 | 316.12589, 304.16135, 258.11682, 193.05042, 165.07048, 153.07048, 147.04456, 125.08416, 118.06568 | L |
| 31 | 238–284 | 10-O-Dimethylgalanthine (lycorine-type) | C17H22O4N+ | 13.48 | 304.15488 | 304.16119 | 22.539 | 274.14847, 266.08557, 258.11697, 191.15506, 167.15486, 125.98666, 118.06574 | B |
| 32 | 248–271 | 7-Methoxy-O-methyllycorenine (homolycorine-type) | C20H28O5N+ | 13.75 | 362.19675 | 362.20648 | 29.405 | 346.13885, 330.14188, 247.12267, 221.16689, 191.15508, 167.01363 | B; L |
| 33 | 238–271 | Unknown alkaloid | C24H26O4N+ | 13.97 | 392.18618 | 392.18298 | −6.769 | 376.18741, 344.15909, 304.16098, 252.10530, 212.14474, 180.10254 | L |
| 34 | 249–278 | Unknown alkaloid | C20H24O4N+ | 14.05 | 342.17053 | 342.17990 | 28.977 | 337.19955, 316.12595, 259.18481, 247.12242, 194.11845, 144.08136 | B |
| 35 | 241–287 | Unknown alkaloid | C19H22O5N+ | 14.24 | 344.16183 | 344.15912 | 28.681 | 337.19943, 316.16238, 282.11783, 227.08345, 191.14377, 110.02042 | L |
| 36 | 248–284 | Homolycorine (homolycorine-type) | C18H22O4N+ | 14.29 | 316.15488 | 316.16257 | −22.206 | 312.16776, 284.18124, 272.13272, 251.15782, 201.13953, 181.17062, 125.98682, 110.02058 | B |
| 37 | 249–284 | Unknown alkaloid | C14H30O9N+ | 14.42 | 356.19151 | 356.19595 | 12.471 | 226.28473, 201.05020, 143.05002, 115.05481, 108.08139 | B; L |
| 38 | 249–282 | Unknown alkaloid | C18H34O7N+ | 14.60 | 376.23298 | 376.22391 | −24.105 | 356.19583, 322.15192, 240.15221, 181.17058, 167.01340, 125.98679 | B |
| 39 | 242 | Unknown alkaloid | C18H34O8N+ | 14.68 | 392.22789 | 392.21906 | −22.522 | 374.17166, 346.17444, 290.15924, 197.11798, 150.09190, 121.06519 | B; L |
| 40 | 243 | Unknown alkaloid | C24H24O5N+ | 14.85 | 406.16490 | 406.16412 | −7.395 | 392.21921, 346.10187, 290.15942, 274.14810, 211.17101, 197.11813, 179.10728 | L |
| 41 | 253–282 | 5-methyl-2-epihippamine isomers (lycorine-type) | C18H22O4N+ | 14.99 | 316.15433 | 316.12613 | 25.878 | 284.18137, 272.13257, 254.16881, 247.12253, 197.08179, 144.08141, 125.98682, 111.09224 | B; L |
| 42 | 252–276 | Unknown alkaloid | C19H30O6N+ | 15.37 | 368.20676 | 368.19748 | 32.171 | 316.12601, 249.14214, 228.14104, 209.20259, 167.01352, 110.02060 | B |
| 43 | 252–278 | Unknown alkaloid | C28H33O5N+ | 15.53 | 463.23532 | 463.24121 | 12.705 | 449.26157, 431.21402, 346.13846, 249.14217, 225.12744, 163.07600 | B |
| 44 | 251–301 | 3-O-methylnarcissidine (lycorine-type) | C19H26O5N+ | 15.85 | 348.18055 | 348.19052 | 28.636 | 346.17471, 274.14804, 197.11795, 171.14987, 138.09195, 121.06521 | L |
| 45 | 250–280 | Unknown alkaloid | C26H32O3N+ | 15.96 | 406.23767 | 406.23715 | −1.281 | 346.13843, 316.12604, 247.12244, 203.11884, 144.08141, | B |
| 46 | 252–297 | 2-O-acetyl-4-O-methyllicorine (lycorine-type) | C19H22O6N+ | 16.33 | 360.14471 | 360.15427 | 28.061 | 314.14539, 267.12448, 247.12241, 211.07701, 171.14987, 121.06534, 103.05473 | L |
| 47 | 253–283 | Jonquailine (tazettine-type) | C19H24O5N+ | 16.45 | 346.16490 | 346.13846 | −76.384 | 321.20441, 314.14566, 288.05582, 247.12248, 171.14989, 102.03439 | B |
| 48 | 261–305 | Unknown alkaloid | C19H22O6N+ | 16.63 | 360.14416 | 360.15417 | 27.783 | 344.12250, 326.14676, 316.12579, 304.16077, 247.12231, 180.10242, 164.10765 | L |
| 49 | 252–282 | Unknown alkaloid | C27H33O3N+ | 16.76 | 419.24550 | 419.24805 | 6.093 | 398.21057, 346.13849, 316.12604, 268.13763, 210.12157, 121.06532 | B |
| 50 | 250–275 | Bulbocapnine (isoquinoline alkaloid) | C19H20O4N+ | 16.85 | 326.13923 | 326.14673 | 24.668 | 282.08136, 274.14810, 240.13322, 225.15038, 207.13930, 138.09189, 121.06530 | L |
| 51 | 253–278 | Unknown alkaloid | C21H32O4N+ | 17.18 | 362.23258 | 362.22394 | −23.866 | 360.21735, 316.12598, 247.12244, 167.01346, 102.03439 | B |
| 52 | 254–297 | Unknown alkaloid | C18H26O7N+ | 17.39 | 368.17038 | 368.17639 | 16.328 | 346.17471, 316.12595, 304.16098, 274.14807, 164.10770, 102.03436 | L |
| 53 | 253–302 | Nerinine (homolycorine-type) | C19H25O5N+ | 18.34 | 347.17272 | 347.17801 | 59.168 | 346.17453, 316.12576, 247.14180, 197.11798, 152.10750, 102.03431 | L |
| 54 | 242–277 | Unknown alkaloid | C24H16O3N+ | 18.82 | 366.11247 | 366.10648 | 28.525 | 344.12265, 316.12579, 274.14801, 256.06381, 167.01337, 110.02050, 102.03431 | L |
| 55 | 285–310 | 1-O-acetylcaranine (lycorine-type) | C18H20O4N+ | 19.24 | 314.13868 | 314.10910 | −94.186 | 304.16092, 278.08636, 270.15344, 252.10562, 247.12234, 226.18208, 191.03456, 102.03433 | L |
| 56 | 266–309-356 | Unknown alkaloid | C23H20O3N+ | 19.41 | 358.14377 | 358.13867 | −14.240 | 336.09265, 314.10925, 278.08649, 191.03464, 102.03435 | L |
| 57 | 251–282 | Unknown alkaloid | C14H21O3N+ | 20.32 | 251.15159 | 251.15793 | 25.223 | 250.14595, 228.14110, 186.09221, 167.01358, 102.03442 | B |
| 58 | 253–327 | Carltonine A (belladine-type) | C27H33O3N2+ | 20.44 | 433.24857 | 433.26575 | 39.654 | 399.18460, 388.15140, 376.18735, 316.12582, 247.12230 167.01335, 122.54755 | L |
| 59 | 251–278 | Unknown alkaloid | C27H38O6N+ | 20.79 | 472.26936 | 472.28061 | 23.811 | 449.26141, 429.29208, 346.13840, 247.12242 144.08139 | B; L |
| 60 | 245–284 | Unknown alkaloid | C24H22O4N+ | 21.06 | 388.15433 | 388.15140 | −7.561 | 330.10541, 316.12585 274.14807, 225.55421, 187.12723, 167.01343 | B; L |
| 61 | 251–285 | Unknown alkaloid | C15H27O3N+ | 21.27 | 269.19909 | 269.20554 | 25.983 | 267.19000, 247.12218, 235.17180 184.10028, 150.09189, 121.06524 | B; L |
| 62 | 252–281 | Unknown alkaloid | C20H33O6N+ | 21.38 | 383.23024 | 383.23532 | 13.258 | 352.34933, 316.12595, 269.20566, 228.14091, 221.15578, 144.08133 | B |
| 63 | 252–297 | 11-Oxo-haemanthamine (haemanthamine-type) | C17H18O4N+ | 21.46 | 300.12303 | 300.12955 | 21.709 | 277.19623, 269.20547, 240.25401, 239.25075, 211.08781, 180.10248, 102.03432 | L |
| 64 | 253–282 | Unknown alkaloid | C14H21O3N+ | 21.89 | 251.15159 | 251.15799 | 25.462 | 247.12265, 212.14493, 186.12865, 167.01361, 144.08147, 102.03445 | B |
| 65 | 255–302 | 1-O-acetylbuphanamine (crinine-type) | C19H22O5N+ | 22.23 | 344.14925 | 344.15903 | −14.851 | 294.11795, 274.14795, 197.11797, 181.12305, 174.12837, 138.09187 | B; L |
| 66 | 257 | 11-Acetylambelline (crinine-type) | C20H24O6N+ | 22.33 | 374.16036 | 374.17181 | 32.060 | 358.13858, 324.13116, 304.16089, 197.11806. 174.12848, 151.11229, 121.06525 | B; L |
| 67 | 255–306 | Maritidine (haemanthamine type) | C17H22O3N+ | 22.78 | 288.15942 | 288.12863 | 27.502 | 244.13684, 216.14043, 191.03470, 167.01357, 122.54771, 102.03446 | B; L |
| 68 | 257–296 | 9-Norpluviine (lycorine-type) | C16H20O3N+ | 23.05 | 274.14432 | 274.14798 | 15.357 | 274.14798, 256.26694, 230.25032, 228.27116, 191.03436, 174.12823, 147.18108, 121.06519, 102.03426 | L |
| 69 | 258 | 5-Methylipseudolycorine (lycorine-type) | C17H22O4N+ | 24.22 | 304.15433 | 304.16098 | 21.848 | 258.28345, 242.28685, 228.27103, 174.12837, 151.11229, 102.03432 | L |
| 70 | 257–292 | Unknown alkaloid | C24H26O3N+ | 24.37 | 376.19072 | 376.18744 | −8.720 | 352.34924, 274.14795, 258.28336, 174.12845, 166.11258, 146.09694, 132.08127 | L |
| Assay | AChE Inhibition IC50 (µg/mL) | BChE Inhibition IC50 (µg/mL) | Tyrosinase Inhibition IC50 (µg/mL) |
|---|---|---|---|
| P. cyrtanthoides bulbs | 4.29 ± 0.04 | 18.32 ± 0.03 | ND |
| P. cyrtanthoides leaves | 8.66 ± 0.03 | 37.70 ± 0.02 | ND |
| Galanthamine | 0.55 ± 0.03 | 3.82 ± 0.02 | - |
| Kojic acid | - | - | 0.76 ± 0.05 |
| Compound | Binding Energy (kcal/mol) Acetylcholinesterase | Binding Energy (kcal/mol) Butyrylcholinesterase |
|---|---|---|
| 3-hydroxydihydrocaranine (13) | −8.67 | −8.06 |
| Kirkine (25) | −8.42 | −8.33 |
| 10-O-dimethylgalanthine (31) | −8.27 | −7.1 |
| 2-α-methoxy-6-O-ethyloduline (22) | −9.38 | −8.21 |
| 10-norpluviine (24) | −8.56 | −7.48 |
| 3-O-acetylnarcissidine (30) | −8.88 | −7.68 |
| Galanthamine | −11.81 | −9.5 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fernández-Galleguillos, C.; Romero-Parra, J.; Puerta, A.; Padrón, J.M.; Simirgiotis, M.J. Alkaloid Profiling, Anti-Enzymatic and Antiproliferative Activity of the Endemic Chilean Amaryllidaceae Phycella cyrtanthoides. Metabolites 2022, 12, 188. https://doi.org/10.3390/metabo12020188
Fernández-Galleguillos C, Romero-Parra J, Puerta A, Padrón JM, Simirgiotis MJ. Alkaloid Profiling, Anti-Enzymatic and Antiproliferative Activity of the Endemic Chilean Amaryllidaceae Phycella cyrtanthoides. Metabolites. 2022; 12(2):188. https://doi.org/10.3390/metabo12020188
Chicago/Turabian StyleFernández-Galleguillos, Carlos, Javier Romero-Parra, Adrián Puerta, José M. Padrón, and Mario J. Simirgiotis. 2022. "Alkaloid Profiling, Anti-Enzymatic and Antiproliferative Activity of the Endemic Chilean Amaryllidaceae Phycella cyrtanthoides" Metabolites 12, no. 2: 188. https://doi.org/10.3390/metabo12020188
APA StyleFernández-Galleguillos, C., Romero-Parra, J., Puerta, A., Padrón, J. M., & Simirgiotis, M. J. (2022). Alkaloid Profiling, Anti-Enzymatic and Antiproliferative Activity of the Endemic Chilean Amaryllidaceae Phycella cyrtanthoides. Metabolites, 12(2), 188. https://doi.org/10.3390/metabo12020188