
Effect of Supplementation with Olive Leaf Extract Enriched with Oleuropein on the Metabolome and Redox Status of Athletes’ Blood and Urine—A Metabolomic Approach


 , , ,
, , ,  ,
, 
Abstract
:
1. Introduction
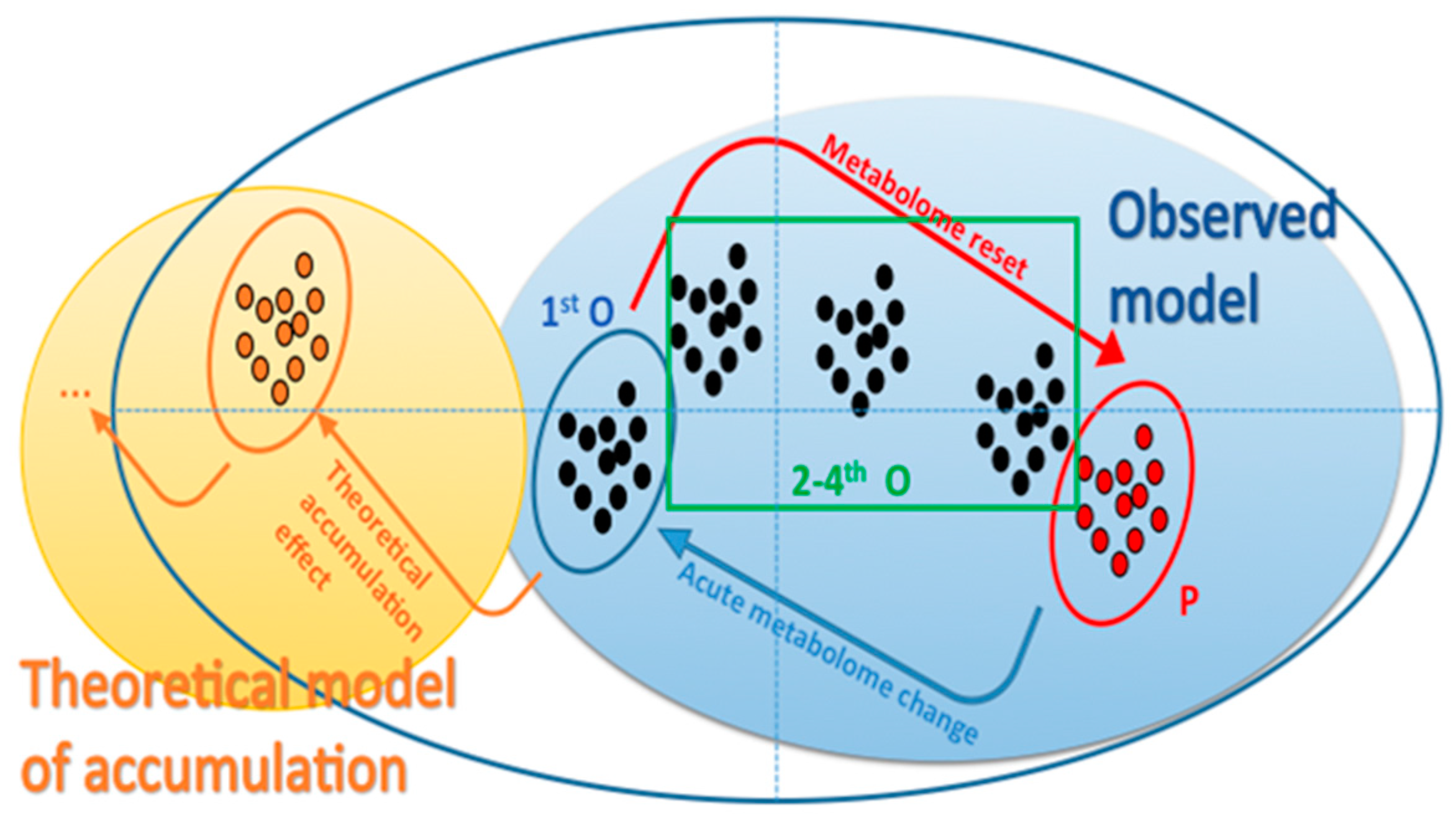
2. Results and Discussion
2.1. Biochemical Analyses
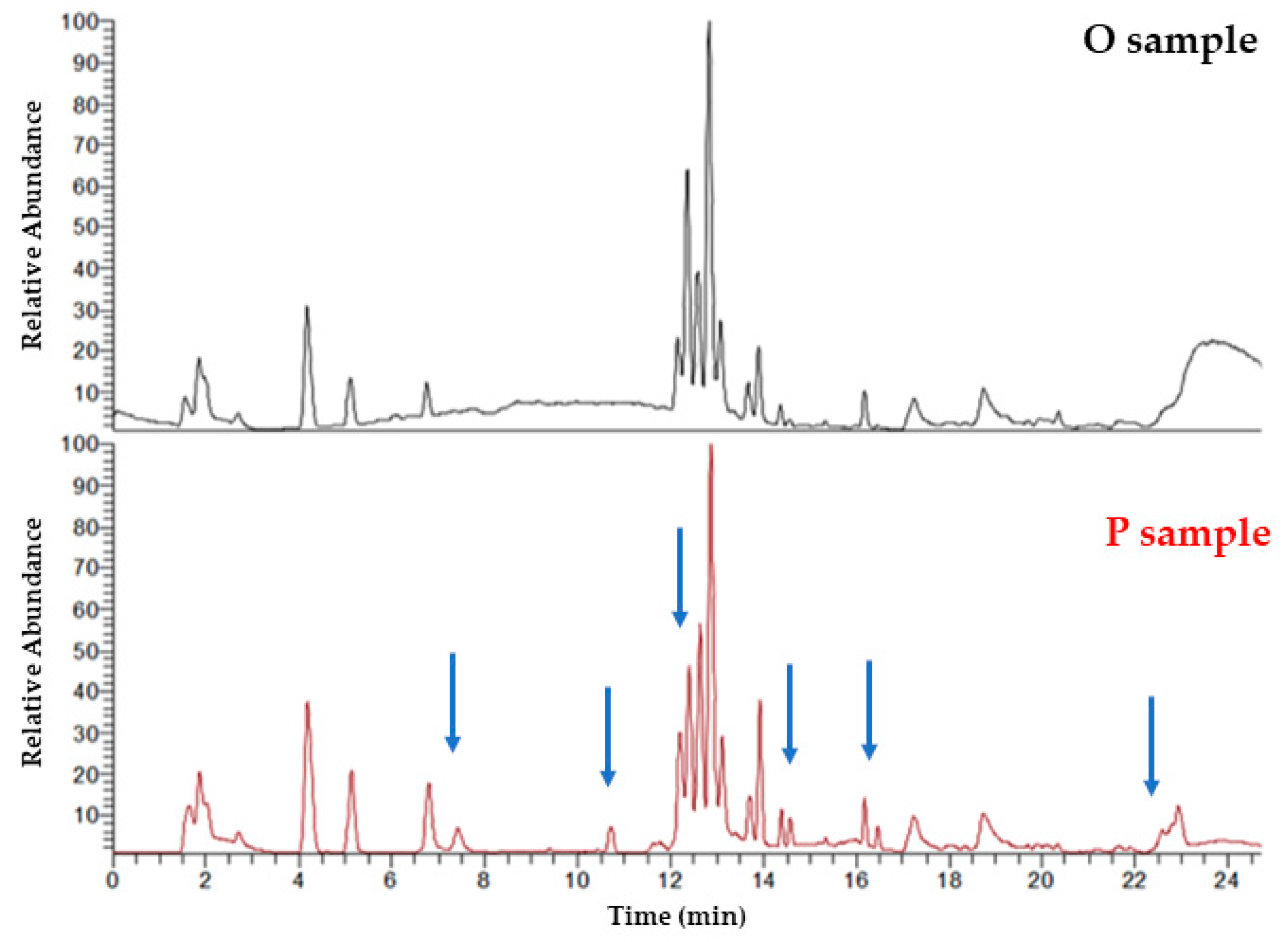
2.2. UHPLC-HRMS Analysis—Data Preprocessing
2.2.1. Peak Picking Procedure
2.2.2. Metaboanalyst Process
2.3. MVA Analysis
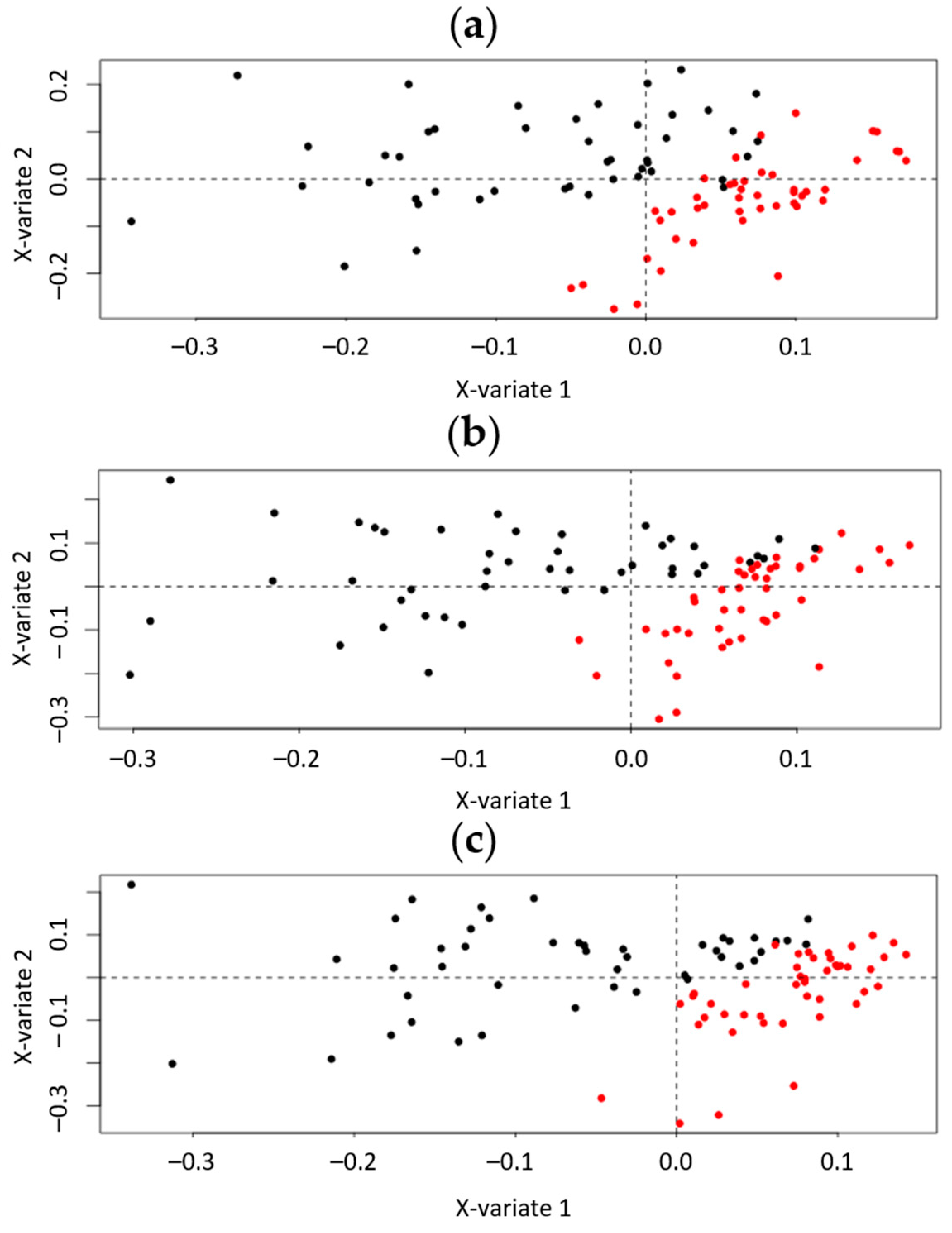
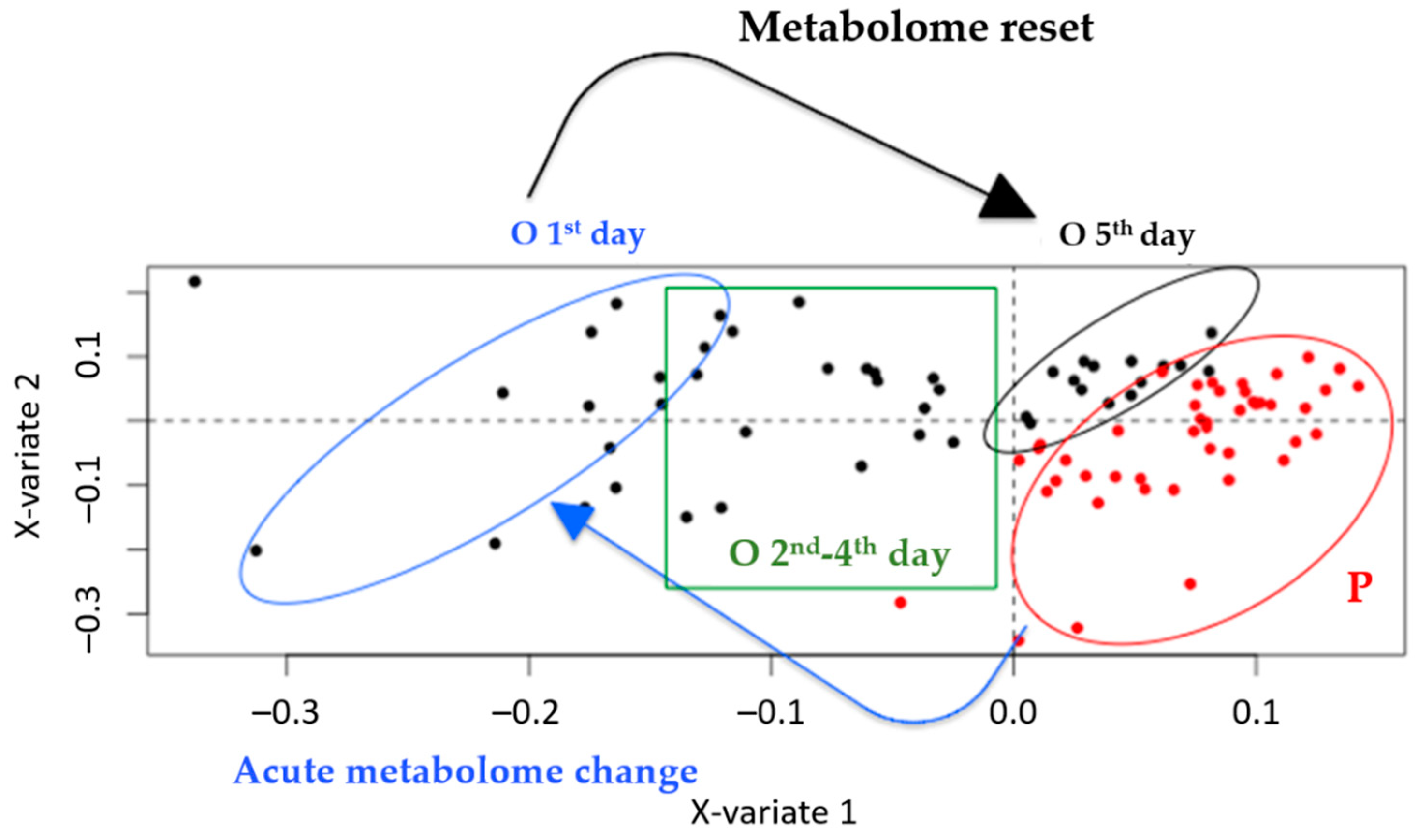
2.3.1. PLS-DA and OPLS-DA Models
2.3.2. PLS-DA and OPLS-DA Models
2.4. Metabolites
2.5. Biological Evaluation
3. Materials and Methods
3.1. Crossover Study Design
3.2. Biofluid Collection
3.3. Samples for the Metabolomic Study
3.4. Chemicals, Reagents and Instrumentation
3.5. Sample Pretreatment
3.6. Biochemical Analyses
3.7. UHPLC-HRMS Analysis
3.7.1. Serum Samples’ Acquisition
3.7.2. Urine Samples’ Acquisition
3.7.3. Quality Control Samples
3.7.4. Mass Spectrometry Data Processing
3.7.5. Statistical Data Analysis
3.7.6. Multilevel Sparse Partial Least Squares–Discriminant Analysis
3.7.7. Variable Selection—Variable Importance in the Projection
3.7.8. Sparse Loadings
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Japón-Luján, R.; de Castro, M.D.L. Superheated liquid extraction of oleuropein and related biophenols from olive leaves. J. Chromatogr. A 2006, 1136, 185–191. [Google Scholar] [CrossRef]
- Romani, A.; Mulinacci, N.; Pinelli, P.; Vincieri, F.F.; Cimato, A. Polyphenolic Content in Five Tuscany Cultivars of Olea europaea L. J. Agric. Food Chem. 1999, 47, 964–967. [Google Scholar] [CrossRef]
- Renaud, S.; De Lorgeril, M.; Delaye, J.; Guidollet, J.; Jacquard, F.; Mamelle, N.; Martin, J.-L.; Monjaud, I.; Salen, P.; Toubol, P. Cretan Mediterranean diet for prevention of coronary heart disease. Am. J. Clin. Nutr. 1995, 61, 1360S–1367S. [Google Scholar] [CrossRef] [PubMed]
- Galli, C.; Visioli, F. Antioxidant and other activities of phenolics in olives/olive oil, typical components of the mediterranean diet. Lipids 1999, 34, S23–S26. [Google Scholar] [CrossRef]
- Kontogianni, V.G.; Gerothanassis, I.P. Phenolic compounds and antioxidant activity of olive leaf extracts. Nat. Prod. Res. 2012, 26, 186–189. [Google Scholar] [CrossRef]
- Pereira, A.P.; Ferreira, I.C.F.R.; Marcelino, F.; Valentão, P.; Andrade, P.B.; Seabra, R.; Estevinho, L.; Bento, A.; Pereira, J.A. Phenolic Compounds and Antimicrobial Activity of Olive (Olea europaea L. Cv. Cobrançosa) Leaves. Molecules 2007, 12, 1153–1162. [Google Scholar] [CrossRef] [PubMed]
- Omar, S.H. Cardioprotective and neuroprotective roles of oleuropein in olive. Saudi Pharm. J. 2010, 18, 111–121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Keys, A.; Mienotti, A.; Karvonen, M.J.; Aravanis, C.; Blackburn, H.; Buzina, R.; Djordjevic, B.S.; Dontas, A.S.; Fidanza, F.; Keys, M.H.; et al. The diet and 15-year death rate in the seven countries study. Am. J. Epidemiol. 1986, 124, 903–915. [Google Scholar] [CrossRef] [PubMed]
- Andreadou, I.; Iliodromitis, E.K.; Mikros, E.; Constantinou, M.; Agalias, A.; Magiatis, P.; Skaltsounis, A.L.; Kamber, E.; Tsantili-Kakoulidou, A.; Kremastinos, D.T. The Olive Constituent Oleuropein Exhibits Anti-Ischemic, Antioxidative, and Hypolipidemic Effects in Anesthetized Rabbits. J. Nutr. 2006, 136, 2213–2219. [Google Scholar] [CrossRef] [Green Version]
- Visioli, F.; Galli, C. Antiatherogenic components of olive oil. Curr. Atheroscler. Rep. 2001, 3, 64–67. [Google Scholar] [CrossRef]
- Scoditti, E.; Calabriso, N.; Massaro, M.; Pellegrino, M.; Storelli, C.; Martines, G.; De Caterina, R.; Carluccio, M.A. Mediterranean diet polyphenols reduce inflammatory angiogenesis through MMP-9 and COX-2 inhibition in human vascular endothelial cells: A potentially protective mechanism in atherosclerotic vascular disease and cancer. Arch. Biochem. Biophys. 2012, 527, 81–89. [Google Scholar] [CrossRef] [PubMed]
- Poudyal, H.; Campbell, F.; Brown, L. Olive Leaf Extract Attenuates Cardiac, Hepatic, and Metabolic Changes in High Carbohydrate–, High Fat–Fed Rats. J. Nutr. 2010, 140, 946–953. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jemai, H.; El Feki, A.; Sayadi, S. Antidiabetic and Antioxidant Effects of Hydroxytyrosol and Oleuropein from Olive Leaves in Alloxan-Diabetic Rats. J. Agric. Food Chem. 2009, 57, 8798–8804. [Google Scholar] [CrossRef] [PubMed]
- Impellizzeri, D.; Esposito, E.; Mazzon, E.; Paterniti, I.; Di Paola, R.; Bramanti, P.; Morittu, V.M.; Procopio, A.; Britti, D.; Cuzzocrea, S. The effects of oleuropein aglycone, an olive oil compound, in a mouse model of carrageenan-induced pleurisy. Clin. Nutr. 2011, 30, 533–540. [Google Scholar] [CrossRef] [PubMed]
- Impellizzeri, D.; Esposito, E.; Mazzon, E.; Paterniti, I.; Di Paola, R.; Bramanti, P.; Morittu, V.M.; Procopio, A.; Perri, E.; Britti, D.; et al. The effects of a polyphenol present in olive oil, oleuropein aglycone, in an experimental model of spinal cord injury in mice. Biochem. Pharmacol. 2012, 83, 1413–1426. [Google Scholar] [CrossRef]
- Agostoni, C.V. Scientific Opinion on the substantiation of health claims related to polyphenols in olive and protection of LDL particles from oxidative damage (ID 1333, 1638, 1639, 1696, 2865), maintenance of normal blood HDL-cholesterol concentrations (ID 1639), mainte. EFSA J. 2012, 9, 2033. [Google Scholar] [CrossRef]
- European Medicines Agency. Assessment Report on Olea europaea L, Folium. Based on Article 16d(1), Article 16f and Article 16h of Directive 2001/83/EC as Amended (Traditional Use); Assessment report on Olea europaea L. folium; EMA/HMPC/430506/2009; European Medicines Agency: London, UK, 2012.
- Cazzola, R.; Russo-Volpe, S.; Cervato, G.; Cestaro, B. Biochemical assessments of oxidative stress, erythrocyte membrane fluidity and antioxidant status in professional soccer players and sedentary controls. Eur. J. Clin. Investig. 2003, 33, 924–930. [Google Scholar] [CrossRef]
- Tasat, D.R.; Mancuso, R.; Evelson, P.A.; Polo, J.M.; Llesuy, S.; Molinari, B. Radiation effects on oxidative metabolism in young and aged rat alveolar macrophages. Cell. Mol. Biol. 2002, 48, 529–535. [Google Scholar]
- Radak, Z.; Chung, H.Y.; Koltai, E.; Taylor, A.W.; Goto, S. Exercise, oxidative stress and hormesis. Ageing Res. Rev. 2008, 7, 34–42. [Google Scholar] [CrossRef]
- Radak, Z.; Chung, H.Y.; Goto, S. Systemic adaptation to oxidative challenge induced by regular exercise. Free Radic. Biol. Med. 2008, 44, 153–159. [Google Scholar] [CrossRef]
- Brites, F.D.; Evelson, P.A.; Christiansen, M.G.; Nicol, M.F.; Basílico, M.J.; Wikinski, R.W.; Llesuy, S.F. Soccer players under regular training show oxidative stress but an improved plasma antioxidant status. Clin. Sci. 1999, 96, 381. [Google Scholar] [CrossRef]
- Kłapcińska, B.; Kempa, K.; Sobczak, A.; Sadowska-Krępa, E.; Jagsz, S.; Szołtysek, I. Evaluation of Autoantibodies Against Oxidized LDL (oLAB) and Blood Antioxidant Status in Professional Soccer Players. Int. J. Sports Med. 2005, 26, 71–78. [Google Scholar] [CrossRef] [PubMed]
- Teixeira, V.H.; Valente, H.F.; Casal, S.I.; Marques, F.P.; Moreira, P.A. Blood antioxidant and oxidative stress biomarkers acute responses to a 1000-m kayak sprint in elite male kayakers. J. Sports Med. Phys. Fit. 2013, 53, 71–79. [Google Scholar]
- Seifi-Skishahr, F.; Siahkohian, M.; Nakhostin-Roohi, B. Influence of aerobic exercise at high and moderate intensities on lipid peroxidation in untrained men. J. Sports Med. Phys. Fit. 2008, 48, 515–521. [Google Scholar]
- World Health Organization. Global Recommendations on Physical Activity for Health; WHO: Genève, Switzerland, 2010; pp. 24–27. ISBN 9789241599979.
- Gomez-Cabrera, M.C.; Domenech, E.; Viña, J. Moderate exercise is an antioxidant: Upregulation of antioxidant genes by training. Free Radic. Biol. Med. 2008, 44, 126–131. [Google Scholar] [CrossRef]
- Gomez-Cabrera, M.-C.; Domenech, E.; Romagnoli, M.; Arduini, A.; Borras, C.; Pallardo, F.V.; Sastre, J.; Viña, J. Oral administration of vitamin C decreases muscle mitochondrial biogenesis and hampers training-induced adaptations in endurance performance. Am. J. Clin. Nutr. 2008, 87, 142–149. [Google Scholar] [CrossRef] [Green Version]
- Margaritis, I.; Palazzetti, S.; Rousseau, A.-S.; Richard, M.-J.; Favier, A. Antioxidant Supplementation and Tapering Exercise Improve Exercise-Induced Antioxidant Response. J. Am. Coll. Nutr. 2003, 22, 147–156. [Google Scholar] [CrossRef]
- Rokitzki, L.; Hinkel, S.; Klemp, C.; Cufi, D.; Keul, J. Dietary, Serum and Urine Ascorbic Acid Status in Male Athletes. Int. J. Sports Med. 1994, 15, 435–440. [Google Scholar] [CrossRef]
- Clarkson, P.M.; Thompson, H.S. Antioxidants: What role do they play in physical activity and health? Am. J. Clin. Nutr. 2000, 72, 637S–646S. [Google Scholar] [CrossRef] [Green Version]
- De Bock, M.; Derraik, J.G.; Brennan, C.M.; Biggs, J.B.; Morgan, P.E.; Hodgkinson, S.C.; Hofman, P.L.; Cutfield, W.S. Olive (Olea europaea L.) Leaf Polyphenols Improve Insulin Sensitivity in Middle-Aged Overweight Men: A Randomized, Placebo-Controlled, Crossover Trial. PLoS ONE 2013, 8, e57622. [Google Scholar] [CrossRef]
- Oliveras-López, M.-J.; Molina, J.J.M.; Mir, M.V.; Rey, E.F.; Martin, F.; de la Serrana, H.L.-G. Extra virgin olive oil (EVOO) consumption and antioxidant status in healthy institutionalized elderly humans. Arch. Gerontol. Geriatr. 2013, 57, 234–242. [Google Scholar] [CrossRef] [PubMed]
- Ruano, J.; Lopez-Miranda, J.; de la Torre, R.; Delgado-Lista, J.; Fernández, J.; Caballero, J.; Covas, M.I.; Jiménez, Y.; Pérez-Martínez, P.; Marín, C.; et al. Intake of phenol-rich virgin olive oil improves the postprandial prothrombotic profile in hypercholesterolemic patients. Am. J. Clin. Nutr. 2007, 86, 341–346. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salvini, S.; Sera, F.; Caruso, D.; Giovannelli, L.; Visioli, F.; Saieva, C.; Masala, G.; Ceroti, M.; Giovacchini, V.; Pitozzi, V.; et al. Daily consumption of a high-phenol extra-virgin olive oil reduces oxidative DNA damage in postmenopausal women. Br. J. Nutr. 2006, 95, 742–751. [Google Scholar] [CrossRef] [PubMed]
- Franconi, F.; Coinu, R.; Carta, S.; Urgeghe, P.P.; Ieri, F.; Mulinacci, A.N.; Romani, A. Antioxidant Effect of Two Virgin Olive Oils Depends on the Concentration and Composition of Minor Polar Compounds. J. Agric. Food Chem. 2006, 54, 3121–3125. [Google Scholar] [CrossRef]
- Jacomelli, M.; Pitozzi, V.; Zaid, M.; Larrosa, M.; Tonini, G.; Martini, A.; Urbani, S.; Taticchi, A.; Servili, M.; Dolara, P.; et al. Dietary extra-virgin olive oil rich in phenolic antioxidants and the aging process: Long-term effects in the rat. J. Nutr. Biochem. 2010, 21, 290–296. [Google Scholar] [CrossRef] [PubMed]
- Bertram, H.C.; Knudsen, K.E.B.; Serena, A.; Malmendal, A.; Nielsen, N.C.; Fretté, X.C.; Andersen, H.J. NMR-based metabonomic studies reveal changes in the biochemical profile of plasma and urine from pigs fed high-fibre rye bread. Br. J. Nutr. 2006, 95, 955–962. [Google Scholar] [CrossRef] [Green Version]
- Pohjanen, E.; Thysell, E.; Jonsson, P.; Eklund, C.; Silfver, A.; Carlsson, I.-B.; Lundgren, K.; Moritz, T.; Svensson, M.B.; Antti, H. A Multivariate Screening Strategy for Investigating Metabolic Effects of Strenuous Physical Exercise in Human Serum. J. Proteome Res. 2007, 6, 2113–2120. [Google Scholar] [CrossRef] [PubMed]
- van Velzen, E.J.J.; Westerhuis, J.A.; van Duynhoven, J.P.M.; van Dorsten, F.A.; Hoefsloot, H.C.J.; Jacobs, D.M.; Smit, S.; Draijer, R.; Kroner, C.I.; Smilde, A.K. Multilevel Data Analysis of a Crossover Designed Human Nutritional Intervention Study. J. Proteome Res. 2008, 7, 4483–4491. [Google Scholar] [CrossRef]
- Jansen, T.; Kristensen, K.; Payne, M.; Edwards, M.; Schrum, C.; Pitois, S. Long-Term Retrospective Analysis of Mackerel Spawning in the North Sea: A New Time Series and Modeling Approach to CPR Data. PLoS ONE 2012, 7, e38758. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rantalainen, M.; Cloarec, O.; Ebbels, T.M.; Lundstedt, T.; Nicholson, J.K.; Holmes, E.; Trygg, J. Piecewise multivariate modelling of sequential metabolic profiling data. BMC Bioinform. 2008, 9, 105. [Google Scholar] [CrossRef] [Green Version]
- Gibney, M.J.; Walsh, M.; Brennan, L.; Roche, H.M.; German, B.; van Ommen, B. Metabolomics in human nutrition: Opportunities and challenges. Am. J. Clin. Nutr. 2005, 82, 497–503. [Google Scholar] [CrossRef] [PubMed]
- Solanky, K.S.; Bailey, N.J.; Beckwith-Hall, B.M.; Bingham, S.; Davis, A.; Holmes, E.; Nicholson, J.; Cassidy, A. Biofluid 1H NMR-based metabonomic techniques in nutrition research—Metabolic effects of dietary isoflavones in humans. J. Nutr. Biochem. 2005, 16, 236–244. [Google Scholar] [CrossRef] [PubMed]
- Rezzi, S.; Ramadan, Z.; Fay, L.B.; Kochhar, S. Nutritional Metabonomics: Applications and Perspectives. J. Proteome Res. 2007, 6, 513–525. [Google Scholar] [CrossRef] [PubMed]
- van der Greef, J.; Martin, S.; Juhasz, P.; Adourian, A.; Plasterer, T.; Verheij, E.R.; McBurney, R.N. The Art and Practice of Systems Biology in Medicine: Mapping Patterns of Relationships. J. Proteome Res. 2007, 6, 1540–1559. [Google Scholar] [CrossRef] [PubMed]
- Lenz, E.; Bright, J.; Wilson, I.; Hughes, A.; Morrisson, J.; Lindberg, H.; Lockton, A. Metabonomics, dietary influences and cultural differences: A 1H NMR-based study of urine samples obtained from healthy British and Swedish subjects. J. Pharm. Biomed. Anal. 2004, 36, 841–849. [Google Scholar] [CrossRef] [PubMed]
- Stacklies, W.; Redestig, H.; Scholz, M.; Walther, D.; Selbig, J. pcaMethods a bioconductor package providing PCA methods for incomplete data. Bioinformatics 2007, 23, 1164–1167. [Google Scholar] [CrossRef] [PubMed]
- Chaouloff, F. Effects of acute physical exercise on central serotonergic systems. Med. Sci. Sports Exerc. 1997, 29, 58–62. [Google Scholar] [CrossRef]
- Badawy, A. Novel nutritional treatment for manic and psychotic disorders: A review of tryptophan and tyrosine depletion studies and the potential of protein-based formulations using glycomacropeptide. Psychopharmacology 2013, 228, 347–358. [Google Scholar] [CrossRef]
- Segura, R.; Ventura, J. Effect of L-Tryptophan Supplementation on Exercise Performance. Int. J. Sports Med. 1988, 9, 301–305. [Google Scholar] [CrossRef]
- Koopman, N.; Katsavelis, D.; Hove, A.S.T.; Brul, S.; de Jonge, W.J.; Seppen, J. The Multifaceted Role of Serotonin in Intestinal Homeostasis. Int. J. Mol. Sci. 2021, 22, 9487. [Google Scholar] [CrossRef]
- Deacon, A.C. The Measurement of 5-Hydroxyindoleacetic Acid in Urine. Ann. Clin. Biochem. 1994, 31, 215–232. [Google Scholar] [CrossRef] [PubMed]
- Meeusen, R.; Thorré, K.; Chaouloff, F.; Sarre, S.; De Meirleir, K.; Ebinger, G.; Michotte, Y. Effects of tryptophan and/or acute running on extracellular 5-HT and 5-HIAA levels in the hippocampus of food-deprived rats. Brain Res. 1996, 740, 245–252. [Google Scholar] [CrossRef]
- Sugimoto, T.; Uchitomi, R.; Hatazawa, Y.; Miura, S.; Kamei, Y. Metabolomic analysis on blood of transgenic mice overexpressing PGC-1α in skeletal muscle. Biosci. Biotechnol. Biochem. 2021, 85, 579–586. [Google Scholar] [CrossRef] [PubMed]
- Gamble, L.-J.; Frye, C.; Hansen, C.; Locasale, J.; Liu, X.; Davis, M.; Wakshlag, J. Serum metabolomics of Alaskan sled dogs during endurance racing. Comp. Exerc. Physiol. 2018, 14, 149–159. [Google Scholar] [CrossRef]
- Lawler, N.G.; Abbiss, C.R.; Gummer, J.P.A.; Broadhurst, D.I.; Govus, A.D.; Fairchild, T.J.; Thompson, K.G.; Garvican-Lewis, L.A.; Gore, C.J.; Maker, G.L.; et al. Characterizing the plasma metabolome during 14 days of live-high, train-low simulated altitude: A metabolomic approach. Exp. Physiol. 2019, 104, 81–92. [Google Scholar] [CrossRef] [PubMed]
- Harshman, S.W.; Pitsch, R.L.; Smith, Z.K.; O’Connor, M.L.; Geier, B.A.; Qualley, A.V.; Schaeublin, N.M.; Fischer, M.V.; Eckerle, J.J.; Strang, A.J.; et al. The proteomic and metabolomic characterization of exercise-induced sweat for human performance monitoring: A pilot investigation. PLoS ONE 2018, 13, e0203133. [Google Scholar] [CrossRef] [Green Version]
- Wyatt, M.; Greathouse, K.L. Targeting Dietary and Microbial Tryptophan-Indole Metabolism as Therapeutic Approaches to Colon Cancer. Nutrients 2021, 13, 1189. [Google Scholar] [CrossRef]
- Huć, T.; Nowinski, A.; Drapala, A.; Konopelski, P.; Ufnal, M. Indole and indoxyl sulfate, gut bacteria metabolites of tryptophan, change arterial blood pressure via peripheral and central mechanisms in rats. Pharmacol. Res. 2018, 130, 172–179. [Google Scholar] [CrossRef]
- Hara, M.; Iigo, M.; Ohtani-Kaneko, R.; Nakamura, N.; Suzuki, T.; Reiter, R.J.; Hirata, K. Administration of Melatonin and Related Indoles Prevents Exercise-Induced Cellular Oxidative Changes in Rats. Neurosignals 1997, 6, 90–100. [Google Scholar] [CrossRef]
- Williams, B.B.; Van Benschoten, A.H.; Cimermancic, P.; Donia, M.S.; Zimmermann, M.; Taketani, M.; Ishihara, A.; Kashyap, P.C.; Fraser, J.; Fischbach, M.A. Discovery and Characterization of Gut Microbiota Decarboxylases that Can Produce the Neurotransmitter Tryptamine. Cell Host Microbe 2014, 16, 495–503. [Google Scholar] [CrossRef] [Green Version]
- Corkery, J.M.; Durkin, E.; Elliott, S.; Schifano, F.; Ghodse, A.H. The recreational tryptamine 5-MeO-DALT (N,N-diallyl-5-methoxytryptamine): A brief review. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2012, 39, 259–262. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Panoutsopoulos, G. Phenylacetaldehyde Oxidation by Freshly Prepared and Cryopreserved Guinea Pig Liver Slices: The Role of Aldehyde Oxidase. Int. J. Toxicol. 2005, 24, 103–109. [Google Scholar] [CrossRef] [PubMed]
- Szabo, A.D.; Billett, E.; Turner, J. Phenylethylamine, a possible link to the antidepressant effects of exercise? Br. J. Sports Med. 2001, 35, 342–343. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cao, B.; Liu, S.; Yang, L.; Chi, A. Changes of Differential Urinary Metabolites after High-Intensive Training in Teenage Football Players. BioMed Res. Int. 2020, 2020, 2073803. [Google Scholar] [CrossRef] [Green Version]
- Timon, R.; Olcina, G.; Muñoz, D.; Maynar, J.I.; Caballero, M.J.; Maynar, M. Determination of Urine Steroid Profile in Untrained Men to Evaluate Recovery After a Strength Training Session. J. Strength Cond. Res. 2008, 22, 1087–1093. [Google Scholar] [CrossRef]
- Mingrone, G.; Castagneto, M. Medium-Chain, Even-Numbered Dicarboxylic Acids as Novel Energy Substrates: An Update. Nutr. Rev. 2006, 64, 449–456. [Google Scholar] [CrossRef]
- Shigenaga, M.K.; Gimeno, C.J.; Ames, B.N. Urinary 8-hydroxy-2′-deoxyguanosine as a biological marker of in vivo oxidative DNA damage. Proc. Natl. Acad. Sci. USA 1989, 86, 9697–9701. [Google Scholar] [CrossRef] [Green Version]
- Dunn, W.B.; Broadhurst, D.; Begley, P.; Zelena, E.; Francis-McIntyre, S.; Anderson, N.; Brown, M.; Knowles, J.D.; Halsall, A.; Haselden, J.N.; et al. Procedures for large-scale metabolic profiling of serum and plasma using gas chromatography and liquid chromatography coupled to mass spectrometry. Nat. Protoc. 2011, 6, 1060–1083. [Google Scholar] [CrossRef]
- Want, E.J.; Wilson, I.; Gika, H.G.; Theodoridis, G.; Plumb, R.S.; Shockcor, J.P.; Holmes, E.; Nicholson, J. Global metabolic profiling procedures for urine using UPLC–MS. Nat. Protoc. 2010, 5, 1005–1018. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| ID | m/z | tR (min) | Complex/ Dimer/ Product/ Precursor | Experimental Deconvoluted m/z (Precursor) | Theoretical m/z (Precursor) | Δm (ppm) | RDB | Pseudo-MS/MS ions | Putative Identification | Molecular Formula | Exact Mass | Trend | Fold Change +/− SD | Dataset |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| M1 | 188.0704 | 1.02 | Product | 205.0970 | 205.0972 | −0.9 | 6.5 | - | L-tryptophan | C11H12N2O2 | 204.0899 | ↑ a | 2.6 (0.4) | Urine ESI(+) |
| M2 | 188.0704 | 1.46 | Precursor | 188.0704 | 188.0703 | −1.0 | 7.5 | - | Indoleacrylic acid | C11H9NO2 | 187.1947 | ↑ | 1.9 (0.5) | Urine ESI(+) |
| M3 | 161.1070 | 3.96 | Precursor | 161.1070 | 161.1073 | −1.9 | 5.5 | - | Tryptamine | C10H12NO2 | 160.2157 | ↑ | 1.9 (0.3) | Urine ESI(+) |
| M4 | 206.0810 | 4.88 | Precursor | 206.0810 | 206.0812 | 0.2 | 6.5 | - | 5-methoxy-indolelactic acid | C11H11NO3 | 205.0738 | ↑ | 1.9 (0.3) | Urine ESI(+) |
| M5 | 221.1534 | 4.77 | Product | 286.2013 | 286.2013 | 0 | 2.5 | - | 2-octenoylcarnitine | C15H27NO4 | 285.3792 | ↑ | 1.6 (0.2) | Urine ESI(+) |
| M6 | 279.1338 | 3.02 | Precursor | 279.1338 | 279.1339 | −0.5 | 6.5 | - | L-phenylalanyl-L-hydroxyproline | C14H18N2O4 | 278.3037 | ↑ | 1.1 (0.3) | Urine ESI(+) |
| M7 | 316.2481 | 5.67 | Precursor | 316.2481 | 316.2482 | −0.4 | 1.5 | - | Decanoylcarnitine | C17H33NO4 | 315.4482 | ↑ | 1.6 (0.3) | Urine ESI(+) |
| M8 | 251.1277 | 5.80 | Precursor | 251.1277 | 251.1278 | –0.3 | 5.5 | - | Ubiquinone-1 | C14H18O4 | 250.2903 | ↑ | 1.1 (0.2) | Urine ESI(+) |
| M9 | 314.2325 | 5.30 | Precursor | 314.2325 | 314.2326 | −0.4 | 2.5 | - | 9-decenoylcarnitine | C17H31NO4 | 313.4323 | ↑ | 1.5 (0.4) | Urine ESI(+) |
| M10 | 302.2325 | 5.23 | Precursor | 302.2325 | 302.2326 | −0.6 | 1.5 | - | 2.6 dimethylheptanoyl carnitine | C16H31NO4 | 301.4216 | ↑ | 1.6 (0.3) | Urine ESI(+) |
| M11 | 130.0494 | 4.30 | Product | 147.0760 | 147.0764 | −2.7 | 1.5 | - | L-glutamine | C5H10N2O3 | 146.1445 | ↓ | Urine ESI(+) | |
| M12 | 293.1471 | 4.30 | Dimer (2M + H) | - | 1.1 (0.4) | |||||||||
| M13 | 147.0760 | 4.30 | Precursor | - | ||||||||||
| M14 | 286.2012 | 4.55 | Precursor | 286.2012 | 286.2013 | −0.5 | 2.5 | - | 2-octenoylcarnitine | C15H27NO4 | 285.3792 | ↓ | 1.7 (0.3) | Urine ESI(+) |
| M15 | 330.2274 | 4.24 | Precursor | 330.2274 | 330.2275 | −0.6 | 2.5 | - | 6-keto-decanoylcarnitine | C17H31NO5 | 329.4317 | ↑ | 1.6 (0.4) | Urine ESI(+) |
| M16 | 105.0329 | 1.71 | Product | 118.0646 | 118.0651 | −4.6 | 5.5 | - | Indole | C8H8N | 117.1479 | ↓ | 1.9 (0.3) | Urine ESI(+) |
| M17 | 137.0593 | 4.49 | Precursor | 137.0593 | 137.0597 | −3 | 4.5 | Phenylacetic acid | C8H8O2 | 136.0524 | ↑ | 1.8 (0.3) | Urine ESI(+) | |
| M18 | 349.1121 | 2.96 | Precursor | 349.1121 | 349.1104 | 4.8 | 8.5 | 215/183/171/157/133 | Estrone-sulfate | C18H22O5S | 350.4290 | ↑ | 1.8 (0.2) | Urine ESI(−) |
| M19 | 201.1134 | 3.58 | Precursor | 201.1134 | 201.1121 | 6.2 | 2.5 | 183/157 | Sebacic acid | C10H18O4 | 202.2475 | ↓ | 1.6 (0.3) | Urine ESI(−) |
| M20 | 389.0989 | 0.90 | Dimer (2M + H) | 194.0452 | 194.0448 | 2.1 | 6.5 | 178/96 | Hydroxyhippuric acid | C9H9NO4 | 195.1721 | ↓ | 1.2 (0.4) | Urine ESI(−) |
| M21 | 357.1092 | 1.71 | Dimer (2M + H) | 178.0501 | 178.0499 | 1.3 | 6.5 | 134/96 | Hippuric acid | C9H9NO3 | 179.1727 | ↑ | 1.2 (0.4) | Urine ESI(−) |
| M22 | 758.5666 | 20.1 | Precursor | 758.5666 | 758.5694 | −3.7 | 3.5 | - | Glycerophospholipid Skeleton | C42H80NO8P | 757.5621 | ↑ | 1.1 (0.2) | Serum ESI(+) |
| M23 | 158.0806 | 10.4 | Precursor | 158.0806 | 158.0812 | −3.1 | 2.5 | 79/80/97/98/99 | Tiglylglycine | C7H11NO3 | 157.1671 | ↓ | 1.1 (0.4) | Serum ESI(+) |
| M24 | 414.2993 | 10.4 | Precursor | 414.2993 | 414.3003 | −2.3 | 7.5 | - | N-docosahexaenoyl GABA | C26H39NO3 | 413.2924 | ↓ | 1.3 (0.2) | Serum ESI(+) |
| M25 | 465.3042 | 24.1 | Product | 465.3042 | 465.3033 | 1.9 | 5.5 | 97/385 | Cholesterol sulfate | C27H46O4S | 466.7170 | ↓ | 1.3 (0.3) | Serum ESI(−) |
| M26 | 167.0214 | 0.9 | Product | 167.0214 | 167.0200 | 6.2 | 6.5 | 96/124 | Uric acid | C5H5O3N4 | 168.1103 | ↓ | 1.1 (0.3) | Serum ESI(−) |
| M27 | 391.2853 | 9.9 | Product | 391.2853 | 391.2843 | 1.2 | 5.5 | – | Bile acid | C24H40O4 | 392.2926 | ↓ | 1.1 (0.3) | Serum ESI(−) |
| M28 | 103.0399 | 1.1 | Product | 103.0399 | 103.0390 | 3.7 | 1.5 | – | Hydroxybutyric acid | C4H8O3 | 104.1045 | ↓ | 1.2 (0.4) | Serum ESI(−) |
| M29 | 135.0302 | 1.0 | Product | 135.0302 | 135.0288 | 6.1 | 1.5 | – | Erythronic acid/Threonic acid | C4H8O5 | 136.1033 | ↓ | 1.3 (0.2) | Serum ESI(−) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lemonakis, N.; Mougios, V.; Halabalaki, M.; Dagla, I.; Tsarbopoulos, A.; Skaltsounis, A.-L.; Gikas, E. Effect of Supplementation with Olive Leaf Extract Enriched with Oleuropein on the Metabolome and Redox Status of Athletes’ Blood and Urine—A Metabolomic Approach. Metabolites 2022, 12, 195. https://doi.org/10.3390/metabo12020195
Lemonakis N, Mougios V, Halabalaki M, Dagla I, Tsarbopoulos A, Skaltsounis A-L, Gikas E. Effect of Supplementation with Olive Leaf Extract Enriched with Oleuropein on the Metabolome and Redox Status of Athletes’ Blood and Urine—A Metabolomic Approach. Metabolites. 2022; 12(2):195. https://doi.org/10.3390/metabo12020195
Chicago/Turabian StyleLemonakis, Nikolaos, Vassilis Mougios, Maria Halabalaki, Ioanna Dagla, Anthony Tsarbopoulos, Alexios-Leandros Skaltsounis, and Evagelos Gikas. 2022. "Effect of Supplementation with Olive Leaf Extract Enriched with Oleuropein on the Metabolome and Redox Status of Athletes’ Blood and Urine—A Metabolomic Approach" Metabolites 12, no. 2: 195. https://doi.org/10.3390/metabo12020195
APA StyleLemonakis, N., Mougios, V., Halabalaki, M., Dagla, I., Tsarbopoulos, A., Skaltsounis, A. -L., & Gikas, E. (2022). Effect of Supplementation with Olive Leaf Extract Enriched with Oleuropein on the Metabolome and Redox Status of Athletes’ Blood and Urine—A Metabolomic Approach. Metabolites, 12(2), 195. https://doi.org/10.3390/metabo12020195