
N-Chlorotaurine Reduces the Lung and Systemic Inflammation in LPS-Induced Pneumonia in High Fat Diet-Induced Obese Mice


 ,
, 
Abstract
:1. Introduction
2. Results
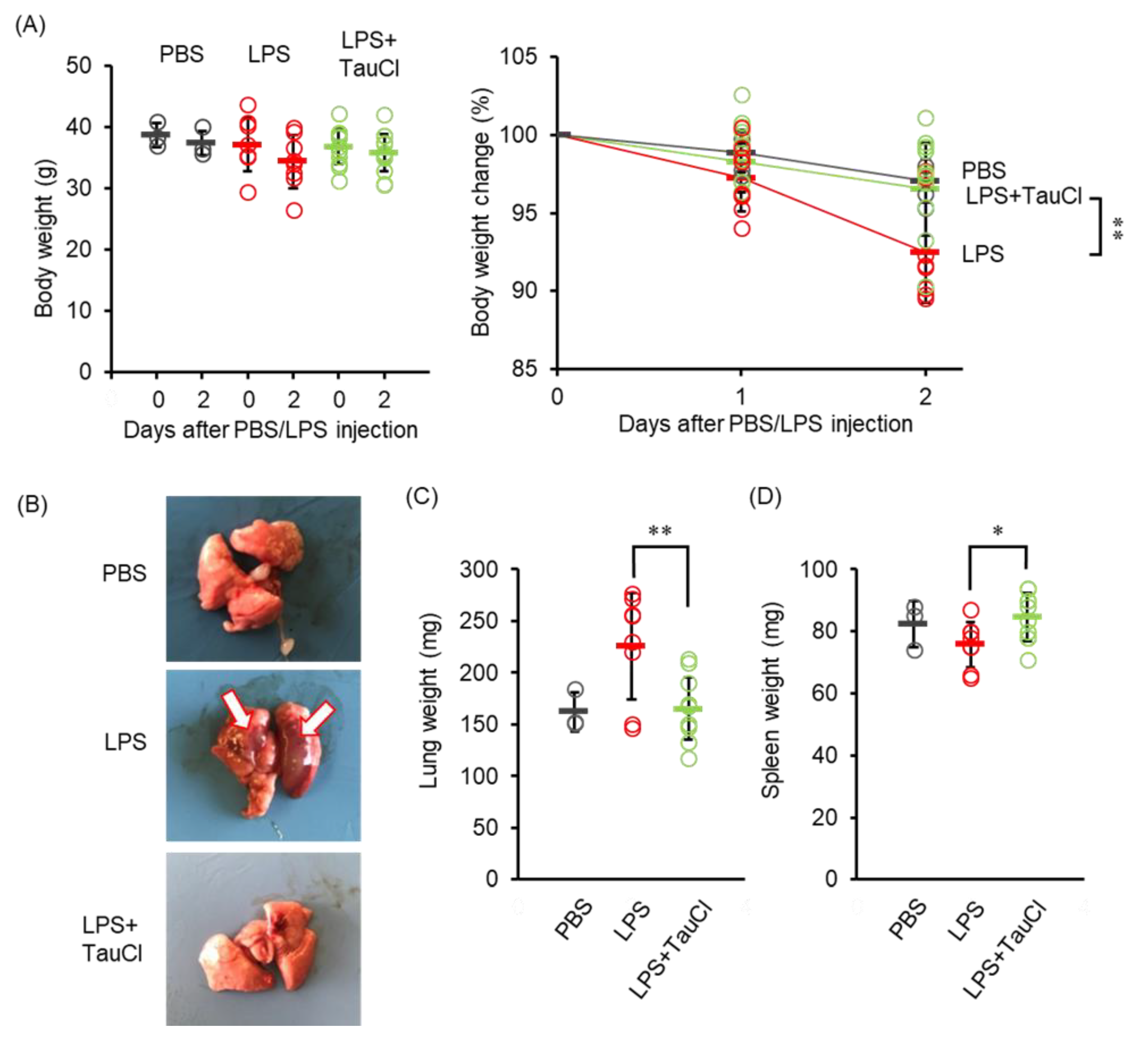
2.1. Effects on Lung Inflammation
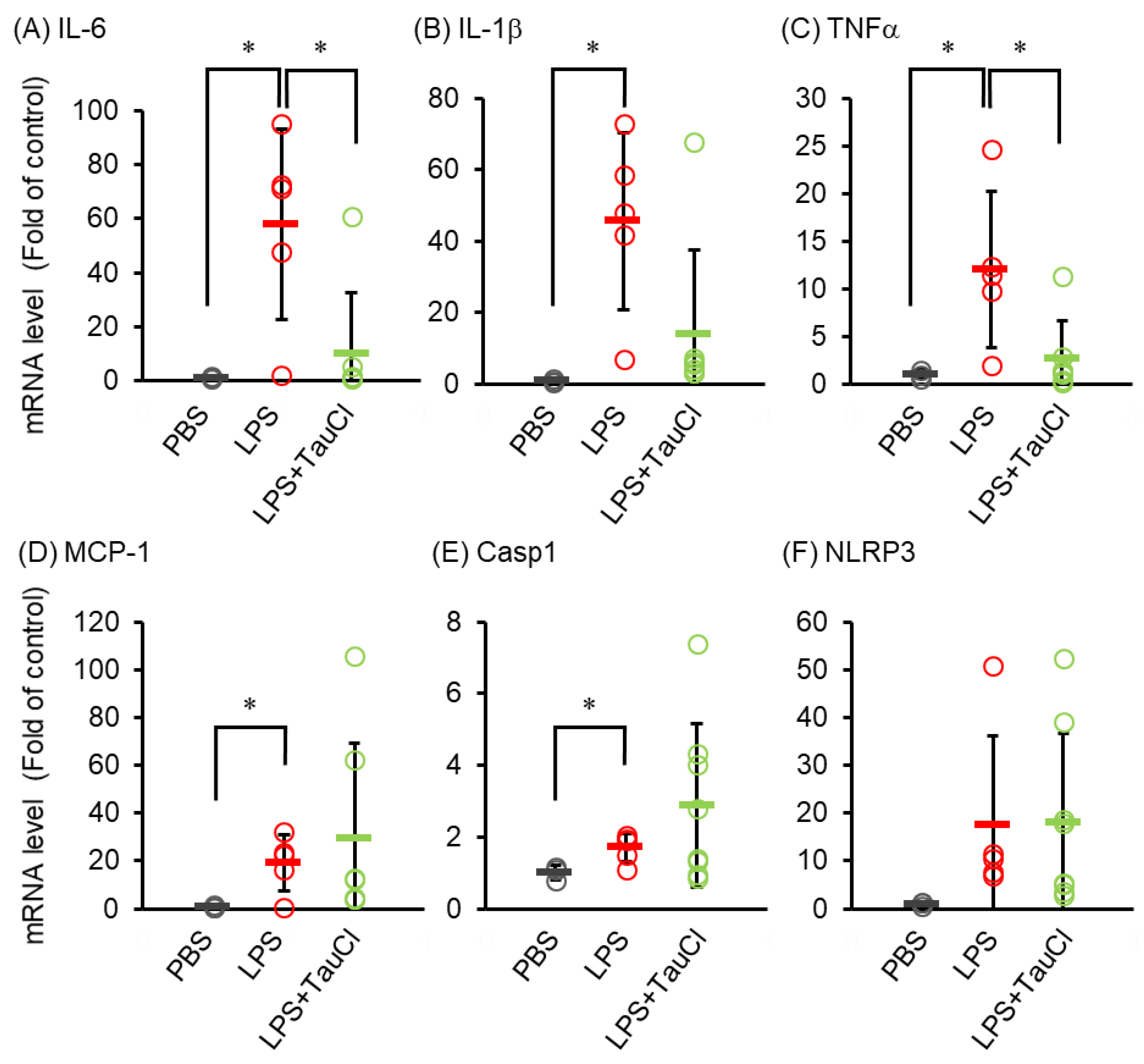
2.2. Effect of TauCl on the LPS-Induced Cytokine mRNA Expression in Lung
2.3. Effects of TauCl on Systemic Cytokine Level in LPS-Treated Mice
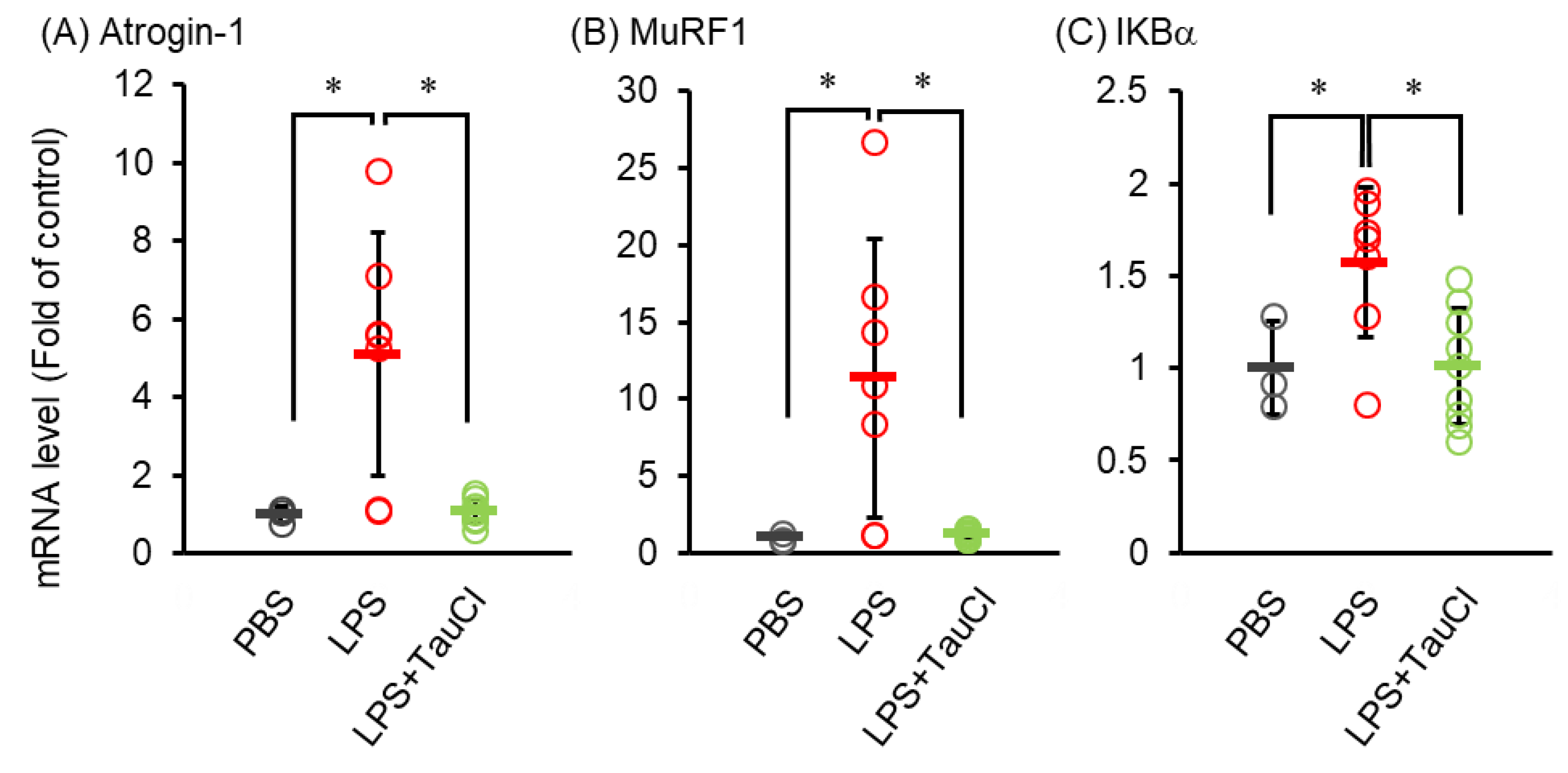
2.4. Effects of TauCl against Muscle Wasting
3. Discussion
4. Materials and Methods
4.1. TauCl Synthesis
4.2. Animal Care
4.3. LPS-Induced Pneumonia Model
4.4. mRNA Measurement
4.5. Plasma Cytokine Assay
4.6. Tissue Section and Immunostaining
4.7. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Fajgenbaum, D.C.; June, C.H. Cytokine Storm. N. Engl. J. Med. 2020, 383, 2255–2273. [Google Scholar] [CrossRef] [PubMed]
- Chesney, R.W. Taurine: Its Biological Role and Clinical Implications. Adv. Pediatr. 1985, 32, 1–42. [Google Scholar] [PubMed]
- Ito, T.; Kimura, Y.; Uozumi, Y.; Takai, M.; Muraoka, S.; Matsuda, T.; Ueki, K.; Yoshiyama, M.; Ikawa, M.; Okabe, M.; et al. Taurine depletion caused by knocking out the taurine transporter gene leads to cardiomyopathy with cardiac atrophy. J. Mol. Cell. Cardiol. 2008, 44, 927–937. [Google Scholar] [CrossRef] [PubMed]
- Tsubotani, K.; Maeyama, S.; Murakami, S.; Schaffer, S.W.; Ito, T. Taurine suppresses liquid–liquid phase separation of lysozyme protein. Amino Acids 2021, 53, 745–751. [Google Scholar] [CrossRef]
- Bhat, M.A.; Ahmad, K.; Khan, M.S.A.; Bhat, M.A.; Almatroudi, A.; Rahman, S.; Jan, A.T. Expedition into Taurine Biology: Structural Insights and Therapeutic Perspective of Taurine in Neurodegenerative Diseases. Biomolecules 2020, 10, 863. [Google Scholar] [CrossRef]
- Ito, T.; Schaffer, S.; Azuma, J. The effect of taurine on chronic heart failure: Actions of taurine against catecholamine and angiotensin II. Amino Acids 2014, 46, 111–119. [Google Scholar] [CrossRef]
- Ito, T.; Schaffer, S.W.; Azuma, J. The potential usefulness of taurine on diabetes mellitus and its complications. Amino Acids 2011, 42, 1529–1539. [Google Scholar] [CrossRef] [Green Version]
- Schuller-Levis, G.B.; Gordon, R.E.; Wang, C.; Park, E. Taurine Reduces Lung Inflammation and Fibrosis Caused by Bleomycin. Adv. Exp. Med. Biol. 2003, 526, 395–402. [Google Scholar] [CrossRef]
- Marcinkiewicz, J.; Kontny, E. Taurine and inflammatory diseases. Amino Acids 2014, 46, 7–20. [Google Scholar] [CrossRef] [Green Version]
- Dudani, A.; Martyres, A.; Fliss, H. Short Communication:Rapid Preparation of Preventive and Therapeutic Whole-Killed Retroviral Vaccines Using the Microbicide Taurine Chloramine. AIDS Res. Hum. Retrovir. 2008, 24, 635–642. [Google Scholar] [CrossRef]
- Nagl, M.; Larcher, C.; Gottardi, W. Activity of N-chlorotaurine against herpes simplex- and adenoviruses. Antivir. Res. 1998, 38, 25–30. [Google Scholar] [CrossRef]
- Nagl, M.; Arnitz, R.; Lackner, M. N-Chlorotaurine, a Promising Future Candidate for Topical Therapy of Fungal Infections. Mycopathologia 2018, 183, 161–170. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gottardi, W.; Nagl, M. N-chlorotaurine, a natural antiseptic with outstanding tolerability. J. Antimicrob. Chemother. 2010, 65, 399–409. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, C.; Cha, Y.-N. Taurine chloramine produced from taurine under inflammation provides anti-inflammatory and cytoprotective effects. Amino Acids 2013, 46, 89–100. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.S.; Ji, H.-I.; Chung, H.; Kim, C.; Lee, S.H.; Lee, Y.-A.; Yang, H.-I.; Yoo, M.C.; Hong, S.J. Taurine chloramine modulates the expression of adipokines through inhibition of the STAT-3 signaling pathway in differentiated human adipocytes. Amino Acids 2013, 45, 1415–1422. [Google Scholar] [CrossRef]
- Kontny, E.; Grabowska, A.; Kowalczewski, J.; Kurowska, M.; Ku-Rowska, M.; Janicka, I. Taurine Chloramine Inhibi-tion of Cell Proliferation and Cytokine Production by Rheumatoid Arthritis Fibroblast-Like Syno-Viocytes. Arthritis Rheum. Off. J. Am. Coll. Rheumatol. 1999, 42, 2552–2560. [Google Scholar] [CrossRef]
- Kanayama, A.; Inoue, J.-I.; Sugita-Konishi, Y.; Shimizu, M.; Miyamoto, Y. Oxidation of IκBα at Methionine 45 Is One Cause of Taurine Chloramine-induced Inhibition of NF-κB Activation. J. Biol. Chem. 2002, 277, 24049–24056. [Google Scholar] [CrossRef] [Green Version]
- Domscheit, H.; Hegeman, M.A.; Carvalho, N.; Spieth, P.M. Molecular Dynamics of Lipopolysaccharide-Induced Lung Injury in Rodents. Front. Physiol. 2020, 11, 36. [Google Scholar] [CrossRef]
- Webster, J.M.; Kempen, L.J.A.P.; Hardy, R.S.; Langen, R.C.J. Inflammation and Skeletal Muscle Wasting During Cachexia. Front. Physiol. 2020, 11, 1449. [Google Scholar] [CrossRef]
- McKinnell, I.W.; Rudnicki, M.A. Molecular Mechanisms of Muscle Atrophy. Cell 2004, 119, 907–910. [Google Scholar] [CrossRef] [Green Version]
- Bivona, J.J.; Crymble, H.M.; Guigni, B.A.; Stapleton, R.D.; Files, D.C.; Toth, M.J.; Poynter, M.E.; Suratt, B.T. Macrophages augment the skeletal muscle proinflammatory response through TNFα following LPS-induced acute lung injury. FASEB J. 2021, 35, e21462. [Google Scholar] [CrossRef] [PubMed]
- Ruland, J. Return to homeostasis: Downregulation of NF-κB responses. Nat. Immunol. 2011, 12, 709–714. [Google Scholar] [CrossRef] [PubMed]
- Cheung, W.W.; Hao, S.; Mak, R.H. Emerging roles of nuclear phosphatase SCP4 in CKD-associated muscle wasting. Kidney Int. 2017, 92, 281–283. [Google Scholar] [CrossRef] [PubMed]
- Lin, S.; Hirai, S.; Yamaguchi, Y.; Goto, T.; Takahashi, N.; Tani, F.; Mutoh, C.; Sakurai, T.; Murakami, S.; Yu, R.; et al. Taurine improves obesity-induced inflammatory responses and modulates the unbalanced phenotype of adipose tissue macrophages. Mol. Nutr. Food Res. 2013, 57, 2155–2165. [Google Scholar] [CrossRef] [PubMed]
- Peskin, A.V.; Winterbourn, C.C. Kinetics of the reactions of hypochlorous acid and amino acid chloramines with thiols, methionine, and ascorbate. Free Radic. Biol. Med. 2001, 30, 572–579. [Google Scholar] [CrossRef]
- Peskina, A.V.; Midwinter, R.G.; Harwood, D.T.; Winterbourn, C.C. Chlorine transfer between glycine, taurine, and histamine: Reaction rates and impact on cellular reactivity. Free Radic. Biol. Med. 2005, 38, 397–405. [Google Scholar] [CrossRef]
- Nakajima, Y.; Osuka, K.; Seki, Y.; Gupta, R.C.; Hara, M.; Takayasu, M.; Wakabayashi, T. Taurine Reduces Inflammatory Responses after Spinal Cord Injury. J. Neurotrauma 2010, 27, 403–410. [Google Scholar] [CrossRef]
- Liu, Y.; Li, F.; Zhang, L.; Wu, J.; Wang, Y.; Yu, H. Taurine alleviates lipopolysaccharide-induced liver injury by anti-inflammation and antioxidants in rats. Mol. Med. Rep. 2017, 16, 6512–6517. [Google Scholar] [CrossRef] [Green Version]
- Kwaśny-Krochin, B.; Bobek, M.; Kontny, E.; Gluszko, P.; Biedroń, R.; Chain, B.; Maśliński, W.; Marcinkiewicz, J. Effect of taurine chloramine, the product of activated neutrophils, on the development of collagen-induced arthritis in DBA 1/J mice. Amino Acids 2002, 23, 419–426. [Google Scholar] [CrossRef]
- Wang, Y.; Cha, Y.-N.; Kim, K.S.; Kim, C. Taurine chloramine inhibits osteoclastogenesis and splenic lymphocyte proliferation in mice with collagen-induced arthritis. Eur. J. Pharmacol. 2011, 668, 325–330. [Google Scholar] [CrossRef]
- Cao, X. COVID-19: Immunopathology and its implications for therapy. Nat. Rev. Immunol. 2020, 20, 269–270. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lackner, M.; Rössler, A.; Volland, A.; Stadtmüller, M.; Müllauer, B.; Banki, Z.; Ströhle, J.; Luttick, A.; Fenner, J.; Stoiber, H.; et al. N-Chlorotaurine, a Novel Inhaled Virucidal Antiseptic Is Highly Active against Respiratory Viruses Including SARS-CoV-2 (COVID-19). 2020. Available online: https://assets.researchsquare.com/files/rs-118665/v1_covered.pdf?c=1631849287 (accessed on 12 April 2022). [CrossRef]
- Ito, T.; Yoshikawa, N.; Ito, H.; Schaffer, S.W. Impact of taurine depletion on glucose control and insulin secretion in mice. J. Pharmacol. Sci. 2015, 129, 59–64. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Langen, R.C.J.; Haegens, A.; Vernooy, J.H.J.; Wouters, E.F.M.; de Winther, M.; Carlsen, H.; Steele, C.; Shoelson, S.E.; Schols, A.M.W.J. NF-κB Activation Is Required for the Transition of Pulmonary Inflammation to Muscle Atrophy. Am. J. Respir. Cell Mol. Biol. 2012, 47, 288–297. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nguyen, K.H.; Ito, S.; Maeyama, S.; Schaffer, S.W.; Murakami, S.; Ito, T. In Vivo and In Vitro Study of N-Methyltaurine on Pharmacokinetics and Antimuscle Atrophic Effects in Mice. ACS Omega 2020, 5, 11241–11246. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Forward Primer (5′→3′) | Reverse Primer (5′→3′) |
|---|---|---|
| IL-1β | CCTTG GGCCT CAAAG GAAAG A | TTGCT TGGGA TCCAC ACTCT CC |
| IL-6 | CACTT CACAA GTCGG AGGCT T | GAATT GCCAT TGCAC AACTC TTTTC |
| TNF-α | CAAAA TTCGA GTGAC AAGCC TGTA | CACCA CTAGT TGGTT GTCTT TGAGA |
| MCP-1 | CTGTC ATGCT TCTGG GCCTG | GGCGT TAACT GCATC TGGCT GA |
| Casp1 | GCATG CCGTG GAGAG AAACA | ATGGG CCTTC TTAAT GCCAT CAT |
| NLRP3 | GCAGA GCCTA CAGTT GGGTG | CTTCC ACGCC TACCA GGAAA T |
| Atrogin-1 | TTCAG CAGCC TGAAC TACGA | AGTAT CCATG GCGCT CCTTC |
| MuRF1 | GCGTG ACCAC AGAGG GTAAA | CTCTG CGTCC AGAGC GTG |
| IκBα | TAGCA GTCTT GACGC AGACC | GACAC GTGTG GCCAT TGTAG |
| GAPDH | GCCGG TGCTG AGTAT GTCGT | CCCTT TTGGC TCCAC CCTT |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Khanh Hoang, N.; Maegawa, E.; Murakami, S.; Schaffer, S.W.; Ito, T. N-Chlorotaurine Reduces the Lung and Systemic Inflammation in LPS-Induced Pneumonia in High Fat Diet-Induced Obese Mice. Metabolites 2022, 12, 349. https://doi.org/10.3390/metabo12040349
Khanh Hoang N, Maegawa E, Murakami S, Schaffer SW, Ito T. N-Chlorotaurine Reduces the Lung and Systemic Inflammation in LPS-Induced Pneumonia in High Fat Diet-Induced Obese Mice. Metabolites. 2022; 12(4):349. https://doi.org/10.3390/metabo12040349
Chicago/Turabian StyleKhanh Hoang, Nguyen, Eiji Maegawa, Shigeru Murakami, Stephen W. Schaffer, and Takashi Ito. 2022. "N-Chlorotaurine Reduces the Lung and Systemic Inflammation in LPS-Induced Pneumonia in High Fat Diet-Induced Obese Mice" Metabolites 12, no. 4: 349. https://doi.org/10.3390/metabo12040349
APA StyleKhanh Hoang, N., Maegawa, E., Murakami, S., Schaffer, S. W., & Ito, T. (2022). N-Chlorotaurine Reduces the Lung and Systemic Inflammation in LPS-Induced Pneumonia in High Fat Diet-Induced Obese Mice. Metabolites, 12(4), 349. https://doi.org/10.3390/metabo12040349