
Taurine Ameliorates Streptozotocin-Induced Diabetes by Modulating Hepatic Glucose Metabolism and Oxidative Stress in Mice





Abstract
:
1. Introduction
2. Results
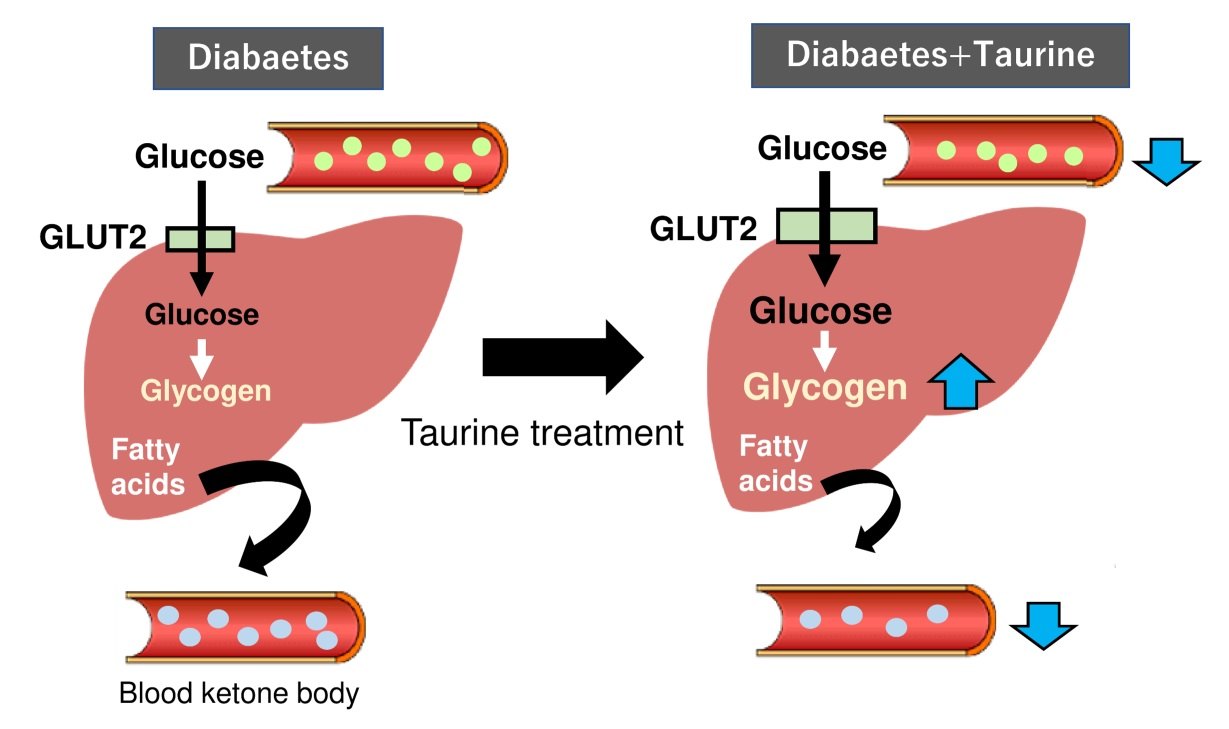
2.1. Body Weight and Blood Levels of Glucose, Insulin, and Ketone Body
2.2. Insulin Resistance
2.3. Glycogen Levels of the Liver and Kidney
2.4. MRNA Expression of Glucose Metabolism-Related Genes in the Liver
2.5. Microarray Analyses
2.6. Oxidative Stress in the Liver and Kidney
2.7. Liver Taurine Content
2.8. Pancreatic Insulin and Taurine
3. Discussion
4. Materials and Methods
4.1. Experimental Animals and Treatment
4.2. Glucose Tolerance Teat
4.3. Biochemical Measurements
4.4. Glucose and Glycogen Determination of Liver and Kidney
4.5. Malondialdehyde (MDA) Determination of Liver and Kidney
4.6. Liver Taurine Content Determination
4.7. RNA Isolation and Real-Time Polymerase Chain Reaction (PCR)
4.8. Histological and Immunohistochemical Studies
4.9. Microarray and Pathway Analyses
4.10. Statistics
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| GK | Glucokinase |
| GLUT-2 | Glucose transporter-2 |
| G6Pase | Glucose-6-phosphatase |
| MDA | Malondialdehyde |
| 8-OHdG | 8-Hydroxy-2′-deoxyguanosine |
| PEPCK | Phosphoenolpyruvate carboxykinase |
| ROS | Reactive oxygen species |
| SREBP-1c | Sterol regulatory element-binding protein 1c |
| STZ | Streptozotocin |
References
- Huxtable, R.J. Physiological actions of taurine. Physiol. Rev. 1992, 72, 101–163. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lambert, I.H.; Kristensen, D.M.; Holm, J.B.; Mortensen, O.H. Physiological role of taurine—From organism to organelle. Acta Physiol. 2015, 213, 191–212. [Google Scholar] [CrossRef] [PubMed]
- Sturman, J.A.; Rassin, D.K.; Gaull, G.E. Taurine in development. Life Sci. 1977, 21, 1–22. [Google Scholar] [CrossRef]
- Albrecht, J.; Schousboe, A. Taurine interaction with neurotransmitter receptors in the CNS: An update. Neurochem. Res. 2005, 30, 1615–1621. [Google Scholar] [CrossRef]
- Spriet, L.L.; Whitfield, J. Taurine and skeletal muscle function. Curr. Opin. Clin. Nutr. Metab. Care 2015, 18, 96–101. [Google Scholar] [CrossRef]
- Lombardini, J.B. Taurine: Retinal function. Brain Res. Rev. 1991, 16, 151–169. [Google Scholar] [CrossRef]
- Jong, C.J.; Sandal, P.; Schaffer, S.W. The role of taurine in mitochondria health: More than just an antioxidant. Molecules 2021, 26, 4913. [Google Scholar] [CrossRef]
- Hardison, W.G. Hepatic taurine concentration and dietary taurine as regulators of bile acid conjugation with taurine. Gastroenterology 1978, 75, 71–75. [Google Scholar] [CrossRef]
- Zulli, A. Taurine in cardiovascular disease. Curr. Opin. Clin. Nutr. Metab. Care 2011, 14, 57–60. [Google Scholar] [CrossRef]
- Menzie, J.; Pan, C.; Prentice, H.; Wu, J.Y. Taurine and central nervous system disorders. Amino Acids 2014, 46, 31–46. [Google Scholar] [CrossRef]
- Murakami, S. Role of taurine in the pathogenesis of obesity. Mol. Nutr. Food Res. 2015, 59, 1353–1363. [Google Scholar] [CrossRef]
- Miyazaki, T.; Matsuzaki, Y. Taurine and liver diseases: A focus on the heterogeneous protective properties of taurine. Amino Acids 2014, 46, 101–110. [Google Scholar] [CrossRef]
- Marcinkiewicz, J.; Kontny, E. Taurine and inflammatory diseases. Amino Acids 2014, 46, 7–20. [Google Scholar] [CrossRef] [Green Version]
- Warskulat, U.; Heller-Stilb, B.; Oermann, E.; Zilles, K.; Haas, H.; Lang, F.; Häussinger, D. Phenotype of the taurine transporter knockout mouse. Methods Enzymol. 2007, 428, 439–458. [Google Scholar]
- Warskulat, U.; Flögel, U.; Jacoby, C.; Hartwig, H.G.; Thewissen, M.; Merx, M.W.; Molojavyi, A.; Heller-Stilb, B.; Schrader, J.; Häussinger, D. Taurine transporter knockout depletes muscle taurine levels and results in severe skeletal muscle impairment but leaves cardiac function uncompromised. FASEB J. 2004, 18, 577–579. [Google Scholar] [CrossRef] [Green Version]
- Ito, T.; Kimura, Y.; Uozumi, Y.; Takai, M.; Muraoka, S.; Matsuda, T.; Ueki, K.; Yoshiyama, M.; Ikawa, M.; Okabe, M.; et al. Taurine depletion caused by knocking out the taurine transporter gene leads to cardiomyopathy with cardiac atrophy. J. Mol. Cell. Cardiol. 2008, 44, 927–937. [Google Scholar] [CrossRef]
- Ito, T.; Yoshikawa, N.; Inui, T.; Miyazaki, N.; Schaffer, S.W.; Azuma, J. Tissue depletion of taurine accelerates skeletal muscle senescence and leads to early death in mice. PLoS ONE 2014, 9, e107409. [Google Scholar] [CrossRef] [Green Version]
- Franconi, F.; Loizzo, A.; Ghirlanda, G.; Seghieri, G. Taurine supplementation and diabetes mellitus. Curr. Opin. Clin. Nutr. Metab. Care 2006, 9, 32–36. [Google Scholar] [CrossRef]
- Imae, M.; Asano, T.; Murakami, S. Potential role of taurine in the prevention of diabetes and metabolic syndrome. Amino Acids 2014, 46, 81–88. [Google Scholar] [CrossRef]
- Tenner, T.E., Jr.; Zhang, X.J.; Lombardini, J.B. Hypoglycemic effects of taurine in the alloxan-treated rabbit, a model for type 1 diabetes. Adv. Exp. Med. Biol. 2003, 526, 97–104. [Google Scholar]
- Trachtman, H.; Futterweit, S.; Maesaka, J.; Ma, C.; Valderrama, E.; Fuchs, A.; Tarectecan, A.A.; Rao, P.S.; Sturman, J.; Boles, T.H.; et al. Taurine ameliorates chronic streptozocin-induced diabetic nephropathy in rats. Am. J. Physiol. 1995, 269, F429–F438. [Google Scholar] [CrossRef]
- Das, J.; Sil, P.C. Taurine ameliorates alloxan-induced diabetic renal injury, oxidative stress-related signaling pathways and apoptosis in rats. Amino Acids 2012, 43, 1509–1523. [Google Scholar] [CrossRef]
- Nandhini, A.T.; Thirunavukkarasu, V.; Anuradha, C.V. Taurine modifies insulin signaling enzymes in the fructose-fed insulin resistant rats. Diabetes Metab. 2005, 31, 337–344. [Google Scholar] [CrossRef]
- Nakaya, Y.; Minami, A.; Harada, N.; Sakamoto, S.; Niwa, Y.; Ohnaka, M. Taurine improves insulin sensitivity in the Otsuka Long-Evans Tokushima Fatty rat, a model of spontaneous type 2 diabetes. Am. J. Clin. Nutr. 2000, 71, 54–58. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, E.J.; Bae, J.H.; Kim, S.Y.; Lim, J.G.; Baek, W.K.; Kwon, T.K.; Suh, S.I.; Park, J.W.; Lee, I.K.; Ashcroft, F.M.; et al. Inhibition of ATP-sensitive K+ channels by taurine through a benzamido-binding site on sulfonylurea receptor 1. Biochem. Pharmacol. 2004, 67, 1089–1096. [Google Scholar] [CrossRef] [PubMed]
- Ribeiro, R.A.; Vanzela, E.C.; Oliveira, C.A.; Bonfleur, M.L.; Boschero, A.C.; Carneiro, E.M. Taurine supplementation: Involvement of cholinergic/phospholipase C and protein kinase A pathways in potentiation of insulin secretion and Ca2+ handling in mouse pancreatic islets. Br. J. Nutr. 2010, 104, 1148–1155. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carneiro, E.M.; Latorraca, M.Q.; Araujo, E.; Beltrá, M.; Oliveras, M.J.; Navarro, M.; Berná, G.; Bedoya, F.J.; Velloso, L.A.; Soria, B.; et al. Taurine supplementation modulates glucose homeostasis and islet function. J. Nutr. Biochem. 2009, 20, 503–511. [Google Scholar] [CrossRef] [PubMed]
- Wu, N.; Lu, Y.; He, B.; Zhang, Y.; Lin, J.; Zhao, S.; Zhang, W.; Li, Y.; Han, P. Taurine prevents free fatty acid-induced hepatic insulin resistance in association with inhibiting JNK1 activation and improving insulin signaling in vivo. Diabetes Res. Clin. Pract. 2010, 90, 288–296. [Google Scholar] [CrossRef]
- Kulakowski, E.C.; Maturo, J. Hypoglycemic properties of taurine: Not mediated by enhanced insulin release. Biochem. Pharmacol. 1984, 33, 2835–2838. [Google Scholar] [CrossRef]
- Maturo, J.; Kulakowski, E.C. Taurine binding to the purified insulin receptor. Biochem. Pharmacol. 1988, 37, 3755–3760. [Google Scholar] [CrossRef]
- Borck, P.C.; Vettorazzi, J.F.; Branco, R.C.S.; Batista, T.M.; Santos-Silva, J.C.; Nakanishi, V.Y.; Boschero, A.C.; Ribeiro, R.A.; Carneiro, E.M. Taurine supplementation induces long-term beneficial effects on glucose homeostasis in ob/ob mice. Amino Acids 2018, 50, 765–774. [Google Scholar] [CrossRef]
- Di Leo, M.; Santini, S.A.; Silveri, N.G.; Giardina, B.; Franconi, F.; Ghirlanda, G. Long-term taurine supplementation reduces mortality rate in streptozotocin-induced diabetic rats. Amino Acids 2004, 27, 187–191. [Google Scholar] [CrossRef]
- Das, J.; Vasan, V.; Sil, P.C. Taurine exerts hypoglycemic effect in alloxan-induced diabetic rats, improves insulin-mediated glucose transport signaling pathway in heart and ameliorates cardiac oxidative stress and apoptosis. Toxicol. Appl. Pharmacol. 2012, 258, 296–308. [Google Scholar] [CrossRef]
- Barthel, A.; Schmoll, D. Novel concepts in insulin regulation of hepatic gluconeogenesis. Am. J. Physiol. Endocrinol. Metab. 2003, 285, E685–E692. [Google Scholar] [CrossRef] [Green Version]
- Thorens, B. GLUT2, glucose sensing and glucose homeostasis. Diabetologia 2015, 58, 221–232. [Google Scholar] [CrossRef] [Green Version]
- Rui, L. Energy metabolism in the liver. Compr. Physiol. 2014, 4, 177–197. [Google Scholar]
- Kanikarla-Marie, P.; Jain, S.K. Hyperketonemia and ketosis increase the risk of complications in type 1 diabetes. Free Radic. Biol. Med. 2016, 95, 268–277. [Google Scholar] [CrossRef] [Green Version]
- Tokunaga, H.; Yoneda, Y.; Kuriyama, K. Streptozotocin-induced elevation of pancreatic taurine content and suppressive effect of taurine on insulin secretion. Eur. J. Pharmacol. 1983, 87, 237–243. [Google Scholar] [CrossRef]
- Lin, S.; Wu, G.; Zhao, D.; Han, J.; Yang, Q.; Feng, Y.; Liu, M.; Yang, J.; Hu, J. Taurine increases insulin expression in STZ-treated rat islet cells in vitro. Adv. Exp. Med. Biol. 2017, 975, 319–328. [Google Scholar]
- Giacco, F.; Brownlee, M. Oxidative stress and diabetic complications. Circ. Res. 2010, 107, 1058–1070. [Google Scholar] [CrossRef] [Green Version]
- Jha, J.C.; Banal, C.; Chow, B.S.; Cooper, M.E.; Jandeleit-Dahm, K. Diabetes and kidney disease: Role of oxidative stress. Antioxid. Redox Signal. 2016, 25, 657–684. [Google Scholar] [CrossRef] [Green Version]
- Hansen, S.H. The role of taurine in diabetes and the development of diabetic complications. Diabetes Metab. Res. Rev. 2001, 17, 330–346. [Google Scholar] [CrossRef]
- Ito, T.; Schaffer, S.W.; Azuma, J. The potential usefulness of taurine on diabetes mellitus and its complications. Amino Acids 2012, 42, 1529–1539. [Google Scholar] [CrossRef] [Green Version]
- Pushpakiran, G.; Mahalakshmi, K.; Anuradha, C.V. Taurine restores ethanol-induced depletion of antioxidants and attenuates oxidative stress in rat tissues. Amino Acids 2004, 27, 91–96. [Google Scholar] [CrossRef]
- Das, J.; Ghosh, J.; Manna, P.; Sil, P.C. Taurine protects acetaminophen-induced oxidative damage in mice kidney through APAP urinary excretion and CYP2E1 inactivation. Toxicology 2010, 269, 24–34. [Google Scholar] [CrossRef]
- Li, S.; Wei, B.K.; Wang, J.; Dong, G.; Wang, X. Taurine supplementation ameliorates arsenic-induced hepatotoxicity and oxidative stress in mouse. Adv. Exp. Med. Biol. 2019, 1155, 463–470. [Google Scholar]
- Murakami, S.; Ono, A.; Kawasaki, A.; Takenaga, T.; Ito, T. Taurine attenuates the development of hepatic steatosis through the inhibition of oxidative stress in a model of nonalcoholic fatty liver disease in vivo and in vitro. Amino Acids 2018, 50, 1279–1288. [Google Scholar] [CrossRef]
- Sullivan, M.A.; Forbes, J.M. Glucose and glycogen in the diabetic kidney: Heroes or villains? EBioMedicine 2019, 47, 590–597. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nannipieri, M.; Lanfranchi, A.; Santerini, D.; Catalano, C.; Van de Werve, G.; Ferrannini, E. Influence of long-term diabetes on renal glycogen metabolism in the rat. Nephron 2001, 87, 50–57. [Google Scholar] [CrossRef] [PubMed]
- Abdulrazaq, N.B.; Cho, M.M.; Win, N.N.; Zaman, R.; Rahman, M.T. Beneficial effects of ginger (Zingiber officinale) on carbohydrate metabolism in streptozotocin-induced diabetic rats. Br. J. Nutr. 2012, 108, 1194–1201. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luc, K.; Schramm-Luc, A.; Guzik, T.J.; Mikolajczyk, T.P. Oxidative stress and inflammatory markers in prediabetes and diabetes. J. Physiol. Pharmacol. 2019, 70, 809–824. [Google Scholar]
- De Luca, G.; Calpona, P.R.; Caponetti, A.; Romano, G.; Di Benedetto, A.; Cucinotta, D.; Di Giorgio, R.M. Taurine and osmoregulation: Platelet taurine content, uptake, and release in type 2 diabetic patients. Metabolism 2001, 50, 60–64. [Google Scholar] [CrossRef]
- Reibel, D.K.; Shaffer, J.E.; Kocsis, J.J.; Neely, J.R. Changes in taurine content in heart and other organs of diabetic rats. J. Mol. Cell Cardiol. 1979, 11, 827–830. [Google Scholar] [CrossRef]
- Nguyen-Ngo, C.; Willcox, J.C.; Lappas, M. Anti-diabetic, anti-inflammatory, and anti-oxidant effects of naringenin in an in vitro human model and an in vivo murine model of gestational diabetes mellitus. Mol. Nutr. Food Res. 2019, 63, e1900224. [Google Scholar] [CrossRef]
- Shi, G.J.; Li, Y.; Cao, Q.H.; Wu, H.X.; Tang, X.Y.; Gao, X.H.; Yu, J.Q.; Chen, Z.; Yang, Y. In vitro and in vivo evidence that quercetin protects against diabetes and its complications: A systematic review of the literature. Biomed. Pharmacother. 2019, 109, 1085–1099. [Google Scholar] [CrossRef]
- Ito, T.; Yoshikawa, N.; Ito, H.; Schaffer, S.W. Impact of taurine depletion on glucose control and insulin secretion in mice. J. Pharmacol. Sci. 2015, 129, 59–64. [Google Scholar] [CrossRef] [Green Version]
- Ma, N.; Aoki, E.; Semba, R. An immunohistochemical study of aspartate, glutamate, and taurine in rat kidney. J. Histochem. Cytochem. 1994, 42, 621–626. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Symbol | Gene Description | Ratio | p-Value |
|---|---|---|---|
| Mt2 | metallothionein 2 | 19.662 | 0.0077 |
| Aldh1b1 | aldehyde dehydrogenase 1 family, member B1 | 2.893 | 0.0013 |
| Slc16a7 | solute carrier family 16, member 7 | 2.822 | 0.0099 |
| Sucnr1 | succinate receptor 1 | 2.767 | 0.0048 |
| Cyp3a59 | cytochrome P450, family 3, subfamily a, polypeptide 59 | 2.611 | 0.0097 |
| Ddah1 | dimethylarginine dimethylaminohydrolase 1 | 2.555 | 0.0055 |
| Hmgn2 | high mobility group nucleosomal binding domain 2 | 2.104 | 0.0072 |
| Hnmt | histamine N-methyltransferase | 2.073 | 0.0078 |
| Sult1a1 | sulfotransferase family 1A, phenol-preferring, member 1 | 2.025 | 0.0023 |
| Cyp2j6 | cytochrome P450, family 2, subfamily j, polypeptide 6 | 2.007 | 0.0015 |
| Siae | sialic acid acetylesterase | 1.986 | 0.0035 |
| Manea | mannosidase, endo-alpha | 1.942 | 0.0090 |
| Papss2 | 3-phosphoadenosine 5-phosphosulfate synthase 2 | 1.932 | 0.0034 |
| Slc6a12 | solute carrier family 6 | 1.926 | 0.0026 |
| Acss2 | acyl-CoA synthetase short-chain family member 2 | 1.925 | 0.0004 |
| Slc2a9 | solute carrier family 2, member 9 | 1.816 | 0.0073 |
| Ech1 | enoyl coenzyme A hydratase 1, peroxisomal | 0.657 | 0.0039 |
| Rela | v-rel reticuloendotheliosis viral oncogene homolog A | 0.644 | 0.0089 |
| Ube2m | ubiquitin-conjugating enzyme E2M | 0.627 | 0.0032 |
| H2afz | H2A histone family, member Z | 0.623 | 0.0073 |
| Ldah | lipid droplet associated hydrolase | 0.615 | 0.0055 |
| Mrpl38 | mitochondrial ribosomal protein L38 | 0.579 | 0.0004 |
| Selk | selenoprotein K | 0.578 | 0.0074 |
| Nfyb | nuclear transcription factor-Y beta | 0.576 | 0.0067 |
| Gucd1 | guanylyl cyclase domain containing 1 | 0.558 | 0.0066 |
| Mtus1 | mitochondrial tumor suppressor 1 | 0.551 | 0.0097 |
| Usp6nl | USP6 N-terminal like | 0.544 | 0.0002 |
| Cd9 | CD9 antigen | 0.543 | 0.0028 |
| Lurap1l | leucine rich adaptor protein 1-like | 0.535 | 0.0008 |
| Gpr182 | G protein-coupled receptor 182 | 0.533 | 0.0078 |
| Paqr9 | progestin and adipoQ receptor family member IX | 0.530 | 0.0023 |
| Arpc1b | actin related protein 2/3 complex, subunit 1B | 0.520 | 0.0001 |
| Inhbc | inhibin beta-C | 0.508 | 0.0087 |
| Crem | cAMP responsive element modulator | 0.482 | 0.0019 |
| Txndc5 | thioredoxin domain containing 5 | 0.453 | 0.0054 |
| Gm10181 | predicted gene 10181 | 0.442 | 0.0074 |
| Myc | myelocytomatosis oncogene | 0.247 | 0.0016 |
| Csad | cysteine sulfinic acid decarboxylase | 0.041 | 0.0048 |
| Gene | Forward Primer | Reverse Primer |
|---|---|---|
| GLUT-2 | TTCATGTCGGTGGGACTTGTG | TGGCAGTCATGCTCACGTAACT |
| G-6-P | AACGTCTGTCTGTCCCGGATCTAC | TTCCGGAGGCTGGCATTGTA |
| GK | GTACGACCGGATGGTGGATG | TCTACCAGCTTGAGCAGCAC |
| PEPCK | CGAATGTGTGGGCGATGAC | ACTGAGGTGCCAGGAGCAACT |
| SREBP1c | AATGACAAGATTGTGGAGCTCAAAG | ACACCAGGTCCTTCAGTGATT |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Murakami, S.; Funahashi, K.; Tamagawa, N.; Ning, M.; Ito, T. Taurine Ameliorates Streptozotocin-Induced Diabetes by Modulating Hepatic Glucose Metabolism and Oxidative Stress in Mice. Metabolites 2022, 12, 524. https://doi.org/10.3390/metabo12060524
Murakami S, Funahashi K, Tamagawa N, Ning M, Ito T. Taurine Ameliorates Streptozotocin-Induced Diabetes by Modulating Hepatic Glucose Metabolism and Oxidative Stress in Mice. Metabolites. 2022; 12(6):524. https://doi.org/10.3390/metabo12060524
Chicago/Turabian StyleMurakami, Shigeru, Kohei Funahashi, Natsuki Tamagawa, Ma Ning, and Takashi Ito. 2022. "Taurine Ameliorates Streptozotocin-Induced Diabetes by Modulating Hepatic Glucose Metabolism and Oxidative Stress in Mice" Metabolites 12, no. 6: 524. https://doi.org/10.3390/metabo12060524
APA StyleMurakami, S., Funahashi, K., Tamagawa, N., Ning, M., & Ito, T. (2022). Taurine Ameliorates Streptozotocin-Induced Diabetes by Modulating Hepatic Glucose Metabolism and Oxidative Stress in Mice. Metabolites, 12(6), 524. https://doi.org/10.3390/metabo12060524