
The Impact of the Metabolic Syndrome and Its Components on Resting Energy Expenditure

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Participant Selection & Ethical Standards
2.2. Statistical Analysis
3. Results
4. Discussion
5. Strengths & Weaknesses
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Donahoo, W.T.; Levine, J.A.; Melanson, E.L. Variability in energy expenditure and its components. Curr. Opin. Clin. Nutr. Metab. Care 2004, 7, 599–605. [Google Scholar] [CrossRef] [PubMed]
- Goran, M.I.; Nagy, T.R.; Gower, B.A.; Mazariegos, M.; Solomons, N.; Hood, V.; Johnson, R. Influence of sex, seasonality, ethnicity and geographic location on the components of total energy expenditure in young children: Implications for energy requirements. Am. J. Clin. Nutr. 1998, 68, 675–682. [Google Scholar] [CrossRef] [PubMed]
- Westerterp, K.R. Control of energy expenditure in humans. Eur. J. Clin. Nutr. 2017, 71, 340–344. [Google Scholar] [CrossRef] [PubMed]
- Soares, M.J.; Müller, M.J. Resting energy expenditure and body composition: Critical aspects for clinical nutrition. Eur. J. Clin. Nutr. 2018, 72, 1208–1214. [Google Scholar] [CrossRef] [PubMed]
- Garby, L.; Lammert, O. Between-subject variation in energy expenditure: Estimation of the effect of variation in organ size. Eur. J. Clin. Nutr. 1994, 48, 376–378. [Google Scholar] [PubMed]
- Piers, L.S.; Soares, M.J.; McCormack, L.M.; O’Deaa, K. Is there evidence for an age-related reduction in metabolic rate? J. Appl. Physiol. 1998, 85, 2196–2204. [Google Scholar] [CrossRef]
- Illner, K.; Brinkmann, G.; Heller, M.; Bosy-Westphal, A.; Müller, M.J. Metabolically active components of fat-free mass and resting energy expenditure in non-obese adults. Am. J. Physiol. Endocrinol. Metab. 2000, 278, E308–E315. [Google Scholar] [CrossRef]
- Löffler, M.C.; Betz, M.J.; Blondin, D.P.; Augustin, R.; Sharma, A.K.; Tseng, Y.H.; Scheele, C.; Zimdahl, H.; Mark, M.; Hennige, A.M.; et al. Challenges in tackling energy expenditure as obesity therapy: From preclinical models to clinical application. Mol. Metab. 2021, 51, 101237. [Google Scholar] [CrossRef]
- Al Adsani, H.; Hoffer, L.J.; Silva, J.E. Resting energy expenditure is sensitive to small dose changes in patients on chronic thyroid hormone replacement. J. Clin. Endocrinol. Metab. 1997, 82, 1118–1125. [Google Scholar] [CrossRef]
- Soares, M.J.; Piers, L.S.; O’Deaa, K.; Collier, G.R. Plasma leptin concentrations, basal metabolic rates and respiratory quotients in young and older adults. Int. J. Obes. Relat. Metab. Disord. 2000, 24, 1592–1599. [Google Scholar] [CrossRef] [Green Version]
- Sepandar, F.; Rashidbeygi, E.; Maghbooli, Z.; Khorrami-Nezhad, L.; Hajizadehoghaz, M.; Mirzaei, K. The association between resting metabolic rate and metabolic syndrome May Be mediated by adipokines in overweight and obese women. Diabetes Metab. Syndr. Clin. Res. Rev. 2019, 13, 530–534. [Google Scholar] [CrossRef] [PubMed]
- Calton, E.K.; Pathak, K.; Soares, M.J.; Alfonso, H.; Keane, K.N.; Newsholme, P.; Cummings, N.K.; Chan She Ping-Delfos, W.; Hamidi, A. Vitamin D status and insulin sensitivity are novel predictors of resting metabolic rate: A cross-sectional analysis in Australian adults. Eur. J. Nutr. 2016, 55, 2075–2080. [Google Scholar] [CrossRef] [PubMed]
- Soares, M.J.; Calton, E.K.; Pathak, K.; Zhao, Y. Hypothesized Pathways for the Association of Vitamin D Status and Insulin Sensitivity with Resting Energy Expenditure: A Cross Sectional Mediation Analysis in Australian Adults of European Ancestry. Eur. J. Clin. Nutr. 2022, Advance online publication. [CrossRef]
- Keane, K.N.; Calton, E.K.; Cruzat, V.F.; Soares, M.J.; Newsholme, P. The impact of cryopreservation on human peripheral blood leucocyte bioenergetics. Clin. Sci. Lond. 2015, 128, 723–733. [Google Scholar] [CrossRef]
- von Hurst, P.R.; Stonehouse, W.; Coad, J. Vitamin D supplementation reduces insulin resistance in South Asian women living in New Zealand who are insulin resistant and vitamin D deficient-a randomised, placebo-controlled trial. Br. J. Nutr. 2010, 103, 549–555. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Belenchia, A.M.; Aneesh, K.T.; Laura, S.H.; Peterson, C.A. Correcting vitamin D insufficiency improves insulin sensitivity in obese adolescents: A randomized controlled trial. Am. J. Clin. Nutr. 2013, 97, 774–781. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Theodoratou, E.; Tzoulaki, L.; Zgaga, L.; Ioannidis, J.P.A. Vitamin D and multiple health outcomes: Umbrella review of systematic reviews and meta-analyses of observational studies and randomised trials. BMJ 2014, 348, g2035. [Google Scholar] [CrossRef] [Green Version]
- Pathak, K.; Soares, M.J.; Zhao, Y.; James, A.P.; Sherriff, J.L.; Newsholme, P. Postprandial changes in glucose oxidation and insulin sensitivity in metabolic syndrome: Influence of fibroblast growth factor 21 and vitamin D status. Nutrition 2017, 37, 37–42. [Google Scholar] [CrossRef]
- Pannu, P.K.; Soares, M.J.; Piers, L.S.; Zhao, Y.; Ansari, Z. The association of vitamin D status and dietary calcium intake with individual components of the metabolic syndrome: A population based study in Victoria, Australia. Cardiovasc. Endocrinol. 2017, 6, 136–144. [Google Scholar] [CrossRef]
- Soares, M.J.; Pannu, P.K.; Calton, E.K.; Hills, A.P.; Read, C.J. Vitamin D status and Calcium intake in systemic inflammation, insulin resistance and the metabolic syndrome: An update on current evidence. Trends Food Sci. Technol. 2017, 62, 79–90. [Google Scholar] [CrossRef]
- Nsatimba, A.; Pathak, K.; Soares, M.J. Ethnic differences in resting metabolic rate, respiratory quotient and body temperature: A comparison of Africans and European Australians. Eur. J. Nutr. 2016, 55, 1831–1838. [Google Scholar] [CrossRef] [PubMed]
- Cooper, J.A.; Watras, A.C.; O’Brienn, M.J.; Luke, A.; Dobratz, J.R.; Earthman, C.P.; Schoeller, D.A. Assessing validity and reliability of resting metabolic rate in six gas analysis systems. J. Am. Diet. Assoc. 2009, 109, 128–132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaviani, S.; Schoeller, D.A.; Ravussin, E.; Melanson, E.L.; Henes, S.T.; Dugas, L.R.; Dechert, R.E.; Mitri, G.; Schoffelen, P.F.; Gubbels, P.; et al. Determining the accuracy and reliability of indirect calorimeters utilizing the methanol combustion technique. Nutr. Clin. Pract. 2018, 33, 206–216. [Google Scholar] [CrossRef] [PubMed]
- Weir, J.B. New methods for calculating metabolic rate with special reference to protein metabolism. Nutrition 1990, 6, 213–221. [Google Scholar] [CrossRef] [PubMed]
- Compher, C.; Frankenfield, D.; Keim, N.; Roth-Yousey, L. Best practice methods to apply to measurement of resting metabolic rate in adults: A systematic review. J. Am. Diet. Assoc. 2006, 106, 881–903. [Google Scholar] [CrossRef]
- Hull, H.; He, Q.; Thornton, J.; Javed, F.; Allen, L.; Wang, J.; Pierson, R.N., Jr.; Gallagher, D. iDXA, Prodigy, and DPXL dual-energy X-ray absorptiometry whole-body scans: A cross-calibration study. J. Clin. Densitom. 2009, 12, 95–102. [Google Scholar] [CrossRef] [Green Version]
- Alberti, K.G. Harmonizing the metabolic syndrome: A joint interim statement of the International Diabetes Federation Task Force on Epidemiology and Prevention; National Heart, Lung, and Blood Institute; American Heart Association; World Heart Federation; International Atherosclerosis Society; and International Association for the Study of Obesity. Circulation 2009, 120, 1640–1645. [Google Scholar]
- Lorenzo, C.; Haffner, S.M.; Stancakova, A.; Laakso, M. Relation of direct and surrogate measures of insulin resistance to cardiovascular risk factors in nondiabetic finnish offspring of type 2 diabetic individuals. J. Clin. Endocrinol. Metab. 2010, 95, 5082–5090. [Google Scholar] [CrossRef]
- Faul, F.; Erdfelder, E.; Buchner, A.; Lang, A.G. Statistical power analyses using G*Power 3.1: Tests for correlation and regression analyses. Behav. Res. Methods 2009, 41, 1149–1160. [Google Scholar] [CrossRef] [Green Version]
- Green, S. How many subjects does it take to do a regression analysis. Multivar. Behav. Res. 1991, 26, 499–510. [Google Scholar] [CrossRef]
- Straub, R.H. The brain and immune system prompt energy shortage in chronic inflammation and ageing. Nat. Rev. Rheumatol. 2017, 13, 743–751. [Google Scholar] [CrossRef] [PubMed]
- Piaggi, P.; Thearle, M.S.; Bogardus, C.; Krakoff, J. Fasting hyperglycemia predicts lower rates of weight gain by increased energy expenditure and fat oxidation rate. J. Clin. Endocrinol. Metabol. 2015, 100, 1078–1087. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wahlqvist, M.L.; Chang, H.Y.; Chen, C.C.; Hsu, C.C.; Chang, W.C.; Wang, W.S.; Hsiung, C.A. Is impaired energy regulation the core of the metabolic syndrome in various ethnic groups of the USA and Taiwan? BMC Endocr. Disord. 2010, 10, 11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rui, L. Energy metabolism in the liver. Compr. Physiol. 2014, 4, 177–197. [Google Scholar] [CrossRef] [Green Version]
- Luke, A.; Adeyemo, A.; Kramer, H.; Forrester, T.; Cooper, R.S. Association between blood pressure and resting energy expenditure independent of body size. Hypertension 2004, 43, 555–560. [Google Scholar] [CrossRef] [Green Version]
- Creber, C.; Cooper, R.S.; Plange-Rhule, J.; Bovet, P.; Lambert, E.V.; Forrester, T.E.; Schoeller, D.; Riesen, W.; Korte, W.; Cao, G.; et al. Independent association of resting energy expenditure with blood pressure: Confirmation in populations of the African diaspora. BMC Cardiovasc. Disord. 2018, 18, 4. [Google Scholar] [CrossRef] [Green Version]
- Ali, N.; Mahmood, S.; Manirujjaman, M.; Perveen, R.; Al Nahid, A.; Ahmed, S.; Khanum, F.A.; Rahman, M. Hypertension prevalence and influence of basal metabolic rate on blood pressure among adult students in Bangladesh. BMC Public Health 2017, 18, 58. [Google Scholar] [CrossRef] [Green Version]
- Monroe, M.B.; Seals, D.R.; Shapiro, L.F.; Bell, C.; Johnson, D.; Jones, P.P. Direct evidence for tonic sympathetic support of resting metabolic rate in health adult humans. Am. J. Physiol. Endocrinol. Metab. 2001, 280, E740–E744. [Google Scholar] [CrossRef]
- Welle, S.; Schwartz, R.G.; Statt, M. Reduced metabolic rate during beta-adrenergic blockade in humans. Metabolism 1991, 40, 619–622. [Google Scholar] [CrossRef]
- Valensi, P. Autonomic nervous system activity changes in patients with hypertension and overweight: Role and therapeutic implications. Cardiovasc. Diabetol. 2021, 20, 170. [Google Scholar] [CrossRef]
- Seravalle, G.; Grassi, G. Sympathetic nervous system, hypertension, obesity and metabolic syndrome. High Blood Press. Cardiovasc. Prev. 2016, 23, 175–179. [Google Scholar] [CrossRef] [PubMed]
- Sharma, A.M.; Pischon, T.; Hardt, S.; Kunz, I.; Luft, F.C. Hypothesis: β-adrenergic receptor blockers and weight gain: A systematic analysis. Hypertension 2001, 37, 250–254. [Google Scholar] [CrossRef] [Green Version]
- Bélanger, M.; Boulay, P. Effect of an aerobic exercise training program on resting metabolic rate in chronically beta-adrenergic blocked hypertensive patients. J. Cardiopulm. Rehabil. Prev. 2005, 25, 354–360. [Google Scholar] [CrossRef] [PubMed]
- Soares, M.J.; Cummings, N.K.; Ping-Delfos, W.L. Energy metabolism and the metabolic syndrome: Does a lower basal metabolic rate signal recovery following weight loss? Diabetes Metab. Syndr. Clin. Res. Rev. 2011, 5, 98–101. [Google Scholar] [CrossRef] [PubMed]
- Larsen, F.J.; Schiffer, T.A.; Sahlin, K.; Ekblom, B.; Weitzberg, W.; Lundberg, J.O. Mitochondrial oxygen affinity predicts basal metabolic rate in humans. FASEB 2011, 25, 2843–2852. [Google Scholar] [CrossRef] [PubMed]
- Bhopal, R.S.; Rafnsson, S.B. Could mitochondrial efficiency explain the susceptibility to adiposity, metabolic syndrome, diabetes and cardiovascular diseases in South Asian populations? Int. J. Epidemiol. 2009, 38, 1072–1081. [Google Scholar] [CrossRef] [PubMed] [Green Version]


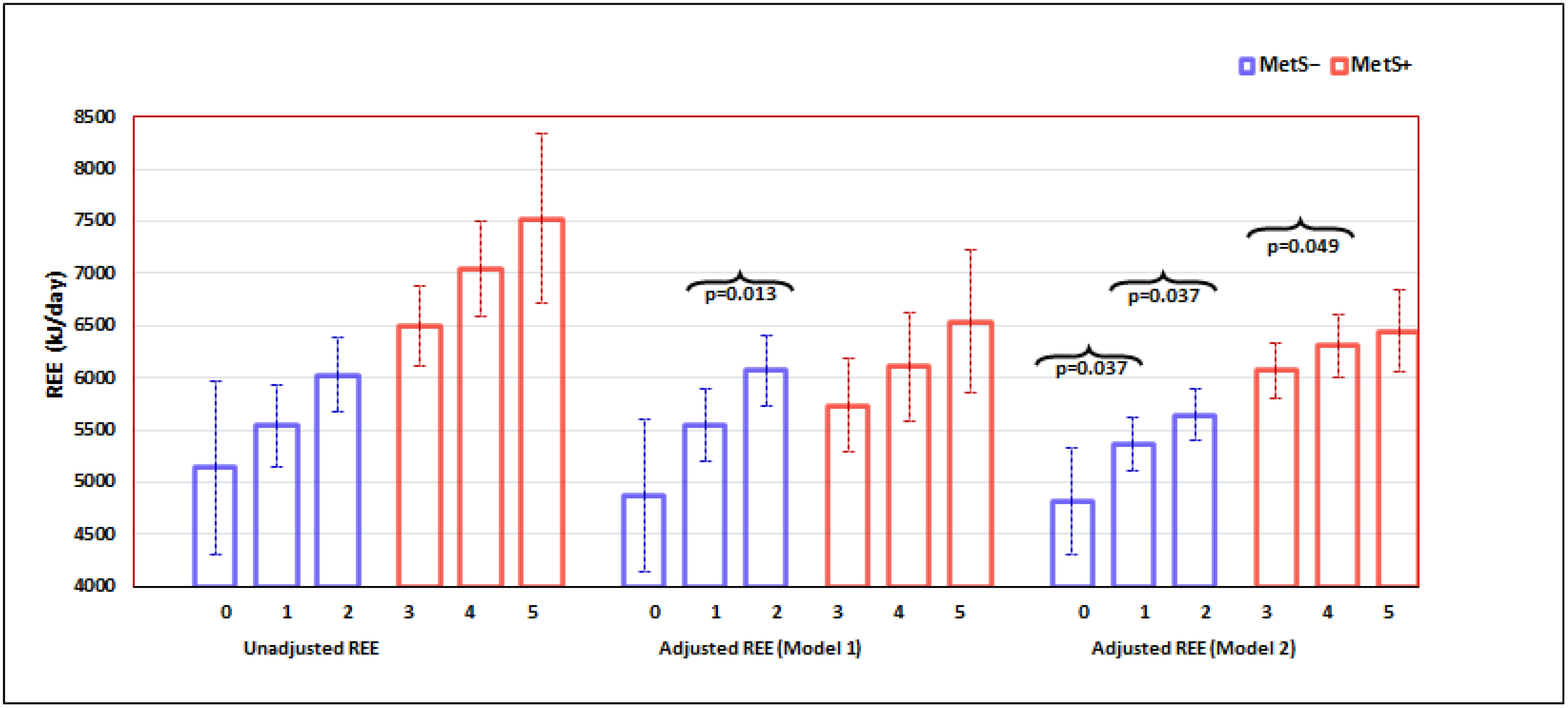{kind=link}
| Variable | MetS− n = 95 | MetS+ n = 85 | p Value * |
|---|---|---|---|
| Age, years | 41.4 ± 14.7 | 55.3 ± 10.5 | 0.001 |
| Gender (n, %) Female Male | 64 (67.4) 31 (32.6) | 44 (51.8) 41 (48.2) | 0.033 |
| Ethnicity (n, %) Sub- Saharan African European | 21 (22.1) 74 (77.9) | 5 (5.9) 80 (94.1) | 0.002 |
| Season (n, %) Winter/spring Summer/autumn | 63 (66.3) 32 (32.7) | 39 (45.9) 46 (54.1) | 0.006 |
| Time of data collection (n, %) 2004–2008 2013–2017 | 20 (21.1) 75 (78.9) | 49 (57.6) 36 (42.4) | 0.001 |
| BMI, kg/m2 Fat mass, kg | 27.2 ± 5.15 27.7 ± 11.2 | 32.9 ± 4.89 37.2 ± 10.3 | 0.001 0.001 |
| Fat-free mass, kg | 50.5 ± 10.9 | 57.4 ± 12.4 | 0.001 |
| Total MetS components (n, %) 0 1 2 3 4 5 | 9 (9.5) 38 (40.0) 48 (50.5) n.a n.a n.a | n.a n.a n.a 44 (51.8) 31 (36.5) 10 (11.8) | 0.001 |
| WC, cm | 91.1 ± 13.6 | 106.0 ± 11.9 | 0.001 |
| FPG, mmol/L | 5.2 ± 0.49 | 6.2 ± 0.88 | 0.001 |
| TG, mmol/L | 1.07 (0.51) | 2.04 (1.1127) | 0.001 |
| HDL-C, mmol/L | 1.85 (0.786) | 1.31 (0.499) | 0.001 |
| SBP, mmHg | 120 ± 13.4 | 133 ± 14.4 | 0.001 |
| DBP, mmHg | 71 ± 8.7 | 79.0 ± 8.8 | 0.001 |
| Inv_IN | 0.606 (0.187) | 0.496 (0.162) | 0.001 |
| 25OHD nmol/L | 60.6 ± 24.08 | 57.2 ± 18.57 | 0.293 |
| MetS− | MetS+ | * p Value | |
|---|---|---|---|
| Unadjusted REE, kJ/d | 5781.4 ± 132.9 | 6814.7 ± 140.5 | <0.001 |
| Adjusted REE, kJ/d (Model 1) | 5408.9 ± 135.6 | 6283.8 ± 138.9 | <0.001 |
| Adjusted REE, kJ/d (Model 2) | 5760.2 ± 86.3 | 5994.1 ± 87.3 | 0.025 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Soares, M.; Zhao, Y.; Calton, E.; Pathak, K.; Chan She Ping-Delfos, W.; Cummings, N.; Nsatimba, P. The Impact of the Metabolic Syndrome and Its Components on Resting Energy Expenditure. Metabolites 2022, 12, 722. https://doi.org/10.3390/metabo12080722
Soares M, Zhao Y, Calton E, Pathak K, Chan She Ping-Delfos W, Cummings N, Nsatimba P. The Impact of the Metabolic Syndrome and Its Components on Resting Energy Expenditure. Metabolites. 2022; 12(8):722. https://doi.org/10.3390/metabo12080722
Chicago/Turabian StyleSoares, Mario, Yun Zhao, Emily Calton, Kaveri Pathak, Wendy Chan She Ping-Delfos, Nicola Cummings, and Patience Nsatimba. 2022. "The Impact of the Metabolic Syndrome and Its Components on Resting Energy Expenditure" Metabolites 12, no. 8: 722. https://doi.org/10.3390/metabo12080722
APA StyleSoares, M., Zhao, Y., Calton, E., Pathak, K., Chan She Ping-Delfos, W., Cummings, N., & Nsatimba, P. (2022). The Impact of the Metabolic Syndrome and Its Components on Resting Energy Expenditure. Metabolites, 12(8), 722. https://doi.org/10.3390/metabo12080722