
Seasonal Variations of Low-Molecular-Weight Organic Acids in Three Evergreen Broadleaf Rhododendron Forests


Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Site
2.2. Sample Collection and Processing
2.3. Instruments and Reagents
2.4. LMWOA Determination Methods
2.5. Statistical Analysis
3. Results
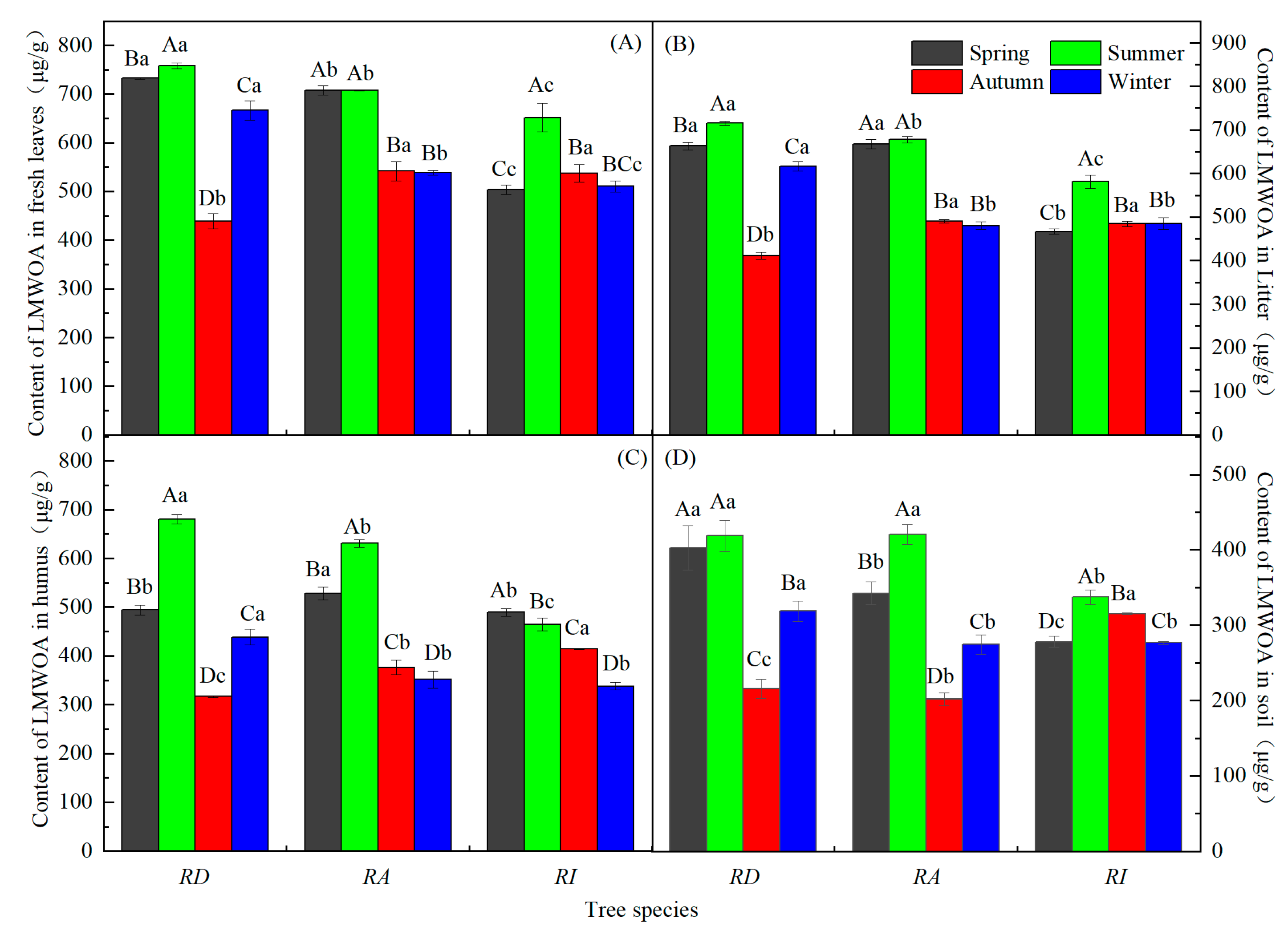
3.1. Seasonal Dynamics of the LMWOAs in Three Forests
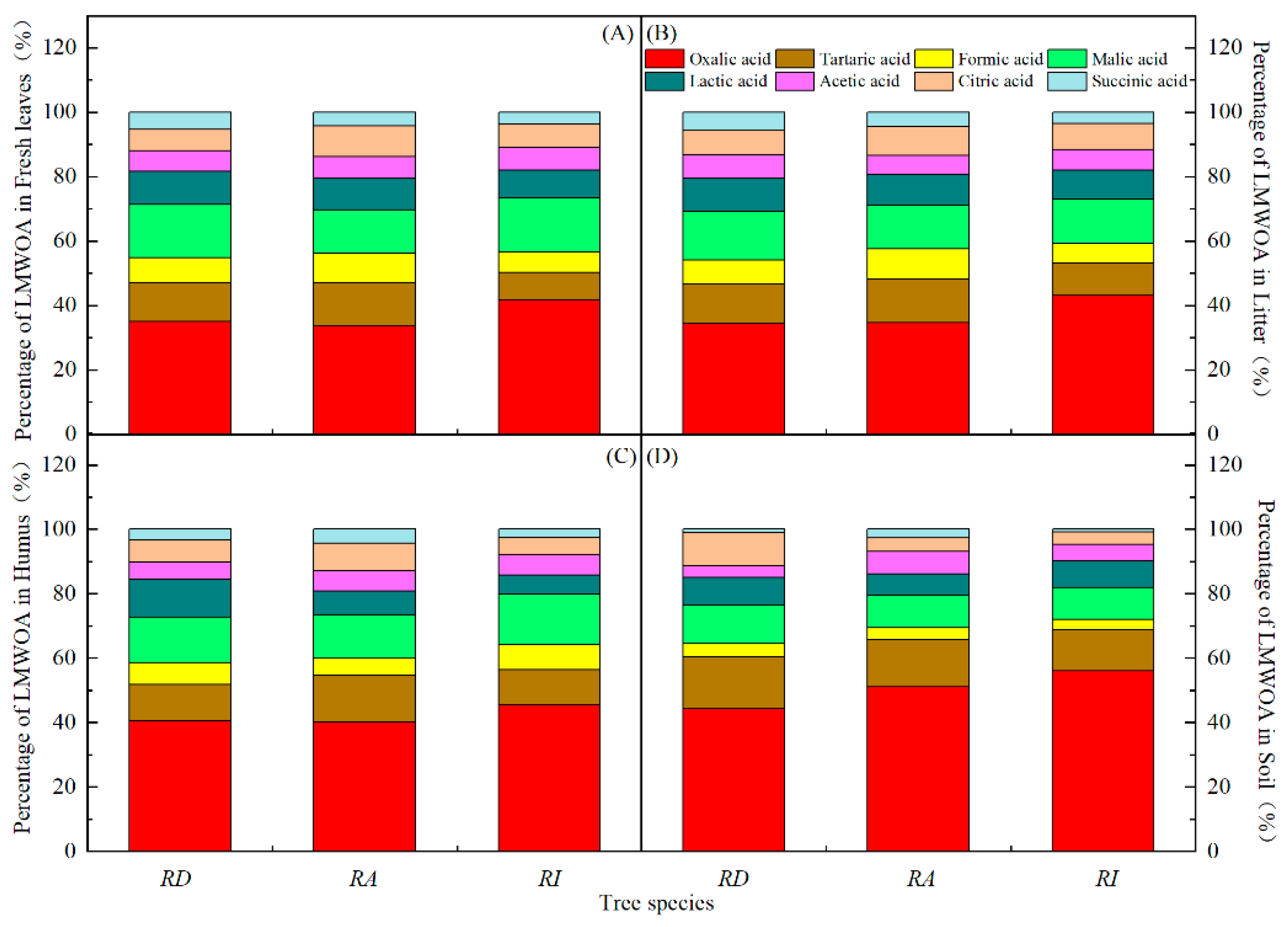
3.2. Seasonal Characteristics of the LMWOAs in Three Forests
3.3. Differences and Characteristics of the LMWOAs in Three Forests
4. Discussion
4.1. Composition and Source of LMWOAs in the Forest
4.2. Seasonal Characteristics of Forest LMWOAs
4.3. Potential Allelopathy of Forest LMWOAs
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hubova, P.; Tejnecky, V.; Ash, C.; Boruvka, L.; Drabek, O. Low-molecular-mass organic acids in the forest soil environment. Mini Rev. Org. Chem. 2017, 14, 75–84. [Google Scholar] [CrossRef] [Green Version]
- Kuzyakov, Y.; Razavi, B.S. Rhizosphere size and shape: Temporal dynamics and spatial stationarity. Soil Biol. Biochem. 2019, 135, 343–360. [Google Scholar] [CrossRef]
- Xiao, M.; Wu, F.C. A review of environmental characteristics and effects of low-molecular weight organic acids in the surface ecosystem. J. Environ. Sci. 2014, 26, 935–954. [Google Scholar] [CrossRef] [PubMed]
- Vondrackova, S.; Szakova, J.; Drabek, O.; Tejnecky, V.; Hejcman, M.; Mullerova, V.; Tlustoš, P. Aluminium uptake and translocation in Al hyperaccumulator Rumex obtusifolius is affected by low-molecular-weight organic acids content and soil pH. PLoS ONE 2015, 10, e0123351. [Google Scholar] [CrossRef] [PubMed]
- Zwetsloot, M.J.; Kessler, A.; Bauerle, T.L. Phenolic root exudate and tissue compounds vary widely among temperate forest tree species and have contrasting effects on soil microbial respiration. New Phytol. 2018, 218, 530–541. [Google Scholar] [CrossRef] [Green Version]
- Mochizuki, T.; Kawamura, K.; Miyazaki, Y.; Boreddy, S.K.R. Distributions and sources of low-molecular-weight monocarboxylic acids in gas and particles from a deciduous broadleaf forest in northern Japan. Atmos. Chem. Phys. 2019, 19, 2421–2432. [Google Scholar] [CrossRef] [Green Version]
- Adeleke, R.; Nwangburuka, C.; Oboirien, B. Origins, roles and fate of organic acids in soils: A review. S. Afr. J. Bot. 2017, 108, 393–406. [Google Scholar] [CrossRef]
- Chen, J.; Lu, T.; Wang, Y.; Li, J.; Fu, X.; Qi, Z.; Zhang, Q. Transport of graphene oxide nanoparticles in saturated kaolinite- and goethite-coated sand columns: Effects of low-molecular-weight organic acids. Environ. Sci. Pollut. Res. 2019, 26, 24922–24932. [Google Scholar] [CrossRef]
- Sokolova, T.A. Low-molecular-weight organic acids in soils: Sources, composition, concentrations, and functions: A review. Eurasian Soil Sci. 2020, 53, 580–594. [Google Scholar] [CrossRef]
- Chi, J.; Zhang, W.; Wang, L.; Putnis, C.V. Direct observations of the occlusion of soil organic matter within calcite. Environ. Sci. Technol. 2019, 53, 8097–8104. [Google Scholar] [CrossRef]
- Schack-Kirchner, H.; Loew, C.A.; Lang, F. The cumulative amount of exuded citrate controls its efficiency to mobilize soil phosphorus. Front. For. Glob. Chang. 2020, 3, 130–142. [Google Scholar] [CrossRef]
- Panchal, P.; Miller, A.J.; Giri, J. Organic acids: Versatile stress-response roles in plants. J. Exp. Bot. 2021, 72, 4038–4052. [Google Scholar] [CrossRef] [PubMed]
- Zou, X.; Wei, D.; Wu, P.; Zhang, Y.; Hu, Y.; Chen, S.; Ma, X. Strategies of organic acid production and exudation in response to low-phosphorus stress in Chinese fir genotypes differing in phosphorus-use efficiencies. Trees 2018, 32, 897–912. [Google Scholar]
- Sokolova, T.A.; Tolpeshta, I.I.; Danilin, I.V.; Izosimova, Y.G.; Chalova, T.S. Acid–base characteristics and clay mineralogy in the rhizospheres of norway maple and common spruce and in the bulk mass of podzolic soil. Eurasian Soil Sci. 2019, 52, 707–717. [Google Scholar] [CrossRef]
- Kong, C.H.; Xuan, T.D.; Khanh, T.D.; Tran, H.D.; Trung, N.T. Allelochemicals and signaling chemicals in plants. Molecules 2019, 24, 2737. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, C.; Quan, W.; Qian, C.; Wu, Y. Distribution of chemical compounds in different soil layers of Rhododendron forest. Allelopath. J. 2019, 48, 191–202. [Google Scholar] [CrossRef]
- Macias-Benitez, S.; Garcia-Martinez, A.M.; Jimenez, P.C.; Gonzalez, J.M.; Moral, M.T.; Rubio, J.P. Rhizospheric organic acids as biostimulants: Monitoring feedbacks on soil microorganisms and biochemical properties. Front. Plant Sci. 2020, 11, 633. [Google Scholar] [CrossRef]
- Kaiser, K.; Guggenberger, G.; Haumaier, L.; Zech, W. The composition of dissolved organic matter in forest soil solutions: Changes induced by seasons and passage through the mineral soil. Org. Geochem. 2002, 33, 307–318. [Google Scholar] [CrossRef]
- Tang, F.; Quan, W.; Li, C.; Huang, X.; Wu, X.; Yang, Q.; Pan, Y.; Xu, T.; Qian, C.; Gu, Y. Effects of small gaps on the relationship among soil properties, topography, and plant species in subtropical Rhododendron secondary forest, southwest China. Int. J. Environ. Res. Public Health 2019, 16, 1919. [Google Scholar] [CrossRef] [Green Version]
- Qian, C.; Quan, W.; Li, C.; Xiang, Z. Analysis of volatile terpenoid compounds in Rhododendron species by multidimensional gas chromatography with quadrupole time-of-flight mass spectrometry. Microchem. J. 2019, 149, 104064. [Google Scholar] [CrossRef]
- Song, G.; Wang, Q.; Jin, J. Leaf photosynthetic capacity of sunlit and shaded mature leaves in a deciduous forest. Forests 2020, 11, 318. [Google Scholar] [CrossRef]
- Olsen, S.R.; Dean, L. Phosphourus. In Methods of Soil Analysis; Black, C.A., Ed.; Academic Press: Cambridge, MA, USA, 1965; pp. 1044–1047. [Google Scholar]
- Bremner, J.M.; Mulvaney, C.S. Nitrogen–Total. In Methods of Soil Analysis, 2nd ed.; Page, A.L., Miller, R.H., Keeney, D.R., Eds.; Academic Press: Cambridge, MA, USA, 1982; pp. 595–622. [Google Scholar]
- Ali, T.; Bylund, D.; Essén, S.A.; Lundström, U.S. Liquid extraction of low molecular mass organic acids and hydroxamate siderophores from boreal forest soil. Soil Biol. Biochem. 2011, 43, 2417–2422. [Google Scholar] [CrossRef]
- Zhao, K.; Wu, Y. Rhizosphere calcareous soil P-extraction at the expense of organic carbon from root-exuded organic acids induced by phosphorus deficiency in several plant species. Soil Sci. Plant Nutr. 2014, 60, 640–650. [Google Scholar] [CrossRef]
- Ohta, T.; Hiura, T. Root exudation of low-molecular-mass-organic acids by six tree species alters the dynamics of calcium and magnesium in soil. Can. J. Soil Sci. 2016, 96, 199–206. [Google Scholar] [CrossRef]
- Dijkstra, F.A.; Geibe, C.; Holmström, S.; Lundström, U.S.; Van Breemen, N. The effect of organic acids on base cation leaching from the forest floor under six North American tree species. Eur. J. Soil Sci. 2001, 52, 205–214. [Google Scholar] [CrossRef] [Green Version]
- Xiang, L.; Chen, X.; Yu, P.; Li, X.; Li, Q. Oxalic acid in root exudates enhances accumulation of perfluorooctanoic acid in lettuce. Environ. Sci. Technol. 2020, 54, 13046–13055. [Google Scholar] [CrossRef]
- Barrow, N.; Abhijit, D.; Arup, S. Mechanisms by which citric acid increases phosphate availability. Plant Soil 2018, 423, 193–204. [Google Scholar] [CrossRef]
- Zhu, H.; Bing, H.; Wu, Y.; Sun, H.; Zhou, J. Low molecular weight organic acids regulate soil phosphorus availability in the soils of subalpine forests, eastern Tibetan Plateau. Catena 2021, 203, 105328. [Google Scholar] [CrossRef]
- Fox, T.R. The Influence of Low-Molecular-Weight Organic Acids on Properties and Processes in Forest Soils. In Carbon Forms and Functions in Forest Soils; William, W., McFee, J., Michael, K., Eds.; Academic Press: Cambridge, MA, USA, 1995; pp. 43–62. [Google Scholar]
- Waithaisong, K.; Robin, A.; Martin, A.; Clairotte, M.; Villeneuve, M.; Plassard, C. Quantification of organic P and low-molecular-weight organic acids in ferralsol soil extracts by ion chromatography. Geoderma 2015, 257, 94–101. [Google Scholar] [CrossRef]
- Oliveira, A.P.; Pereira, J.A.; Andrade, P.B.; Valentão, P.; Seabra, R.M.; Silva, B.M. Organic acids composition of Cydonia oblonga Miller leaf. Food Chem. 2008, 111, 393–399. [Google Scholar] [CrossRef]
- Falasca, A.; Melckl, D.; Paris, D.; Savianol, G.; Motta, A.; Iorizzi, M. Seasonal changes in the metabolic fingerprint of Juniperus communis L. berry extracts by 1H NMR-based metabolomics. Metabolomics 2014, 10, 165–174. [Google Scholar] [CrossRef]
- Kimura, F.; Sato, M.; Kato-Noguchi, H. Allelopathy of pine litter: Delivery of allelopathic substances into forest floor. J. Plant Biol. 2015, 58, 61–67. [Google Scholar] [CrossRef]
- Fu, Y.; Quan, W.; Li, C.; Qian, C.; Tang, F.; Chen, X. Allelopathic effects of phenolic acids on seedling growth and photosynthesis in Rhododendron delavayi Franch. Photosynthetica 2019, 57, 377–387. [Google Scholar] [CrossRef] [Green Version]
- Pan, Y.; Li, C.; Hao, J.; Gao, Y.; Quan, W. Physiological responses of Rhododendron delavayi Franch seedlings to glycolic acid. Allelopath. J. 2021, 54, 47–59. [Google Scholar] [CrossRef]
- Van Hees, P.A.W.; Godbold, D.L.; Jentschke, G.; Jones, D.L. Impact of ectomycorrhizas on the concentration and biodegradation of simple organic acids in a forest soil. Eur. J. Soil Sci. 2003, 54, 697–706. [Google Scholar] [CrossRef]
- Gatti, A.B.; Takao, L.K.; Pereira, V.C.; Ferreira, A.G.; Lima, M.I.S.; Gualtieri, S.C.J. Seasonality effect on the allelopathy of cerrado species. Braz. J. Biol. 2014, 74, 64–69. [Google Scholar] [CrossRef] [Green Version]
- Tomar, N.S.; Sharma, M.; Agarwal, R.M. Phytochemical analysis of Jatropha curcas L. during different seasons and developmental stages and seedling growth of wheat (Triticum aestivum L.) as affected by extracts/leachates of Jatropha curcas L. Physiol. Mol. Biol. Plants 2015, 21, 83–92. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| General Situation | RD | RA | RI |
|---|---|---|---|
| Altitude (m) | 1700 | 1658 | 1701 |
| Latitude and longitude | 105°51′49.76″ E 27°14′7.01″ N | 105°51′7.94″ E 27°14′5.11″ N | 105°51′39.54″ E 27°14′32.2″ N |
| Thickness of litter (cm) | 3~5 | 2~4 | 4~7 |
| Thickness of humus (cm) | 10.5 | 16.5 | 20.4 |
| Slope position | Mid-slope | Uphill | Uphill |
| Aspect | Southeast 115° | Southeast 128° | East 102° |
| Slope | 22° | 30° | 30° |
| Soil Properties | RD | RA | RI |
|---|---|---|---|
| pH | 4.31 ± 0.06 | 4.87 ± 0.08 | 4.80 ± 0.08 |
| Humidity (%) | 68.40 ± 2.66 | 62.50 ± 5.00 | 62.20 ± 1.96 |
| SOC (g/kg) | 81.27 ± 4.74 | 101.01 ± 6.09 | 97.94 ± 9.85 |
| TN (g/kg) | 2.15 ± 0.11 | 2.24 ± 0.16 | 4.00 ± 0.32 |
| HN (mg/kg) | 40.13 ± 1.39 | 43.54 ± 1.56 | 59.89 ± 9.68 |
| TP (g/kg) | 0.63 ± 0.04 | 0.67 ± 0.03 | 0.77 ± 0.05 |
| AP (mg/kg) | 0.42 ± 0.03 | 0.28 ± 0.02 | 1.02 ± 0.06 |
| TK (g/kg) | 3.38 ± 0.20 | 3.71 ± 0.12 | 2.84 ± 0.19 |
| AK (mg/kg) | 48.82 ± 2.29 | 44.93 ± 2.08 | 50.40 ± 4.73 |
| Factors | df | Oxalic Acid | Tartaric Acid | Formic Acid | Malic Acid | Lactic Acid | Acetic Acid | Citric Acid | Succinic Acid |
|---|---|---|---|---|---|---|---|---|---|
| Species | 2 | 0.639 ns | 8.15 ns | 1.504 ns | 1.645 ns | 2.052 ns | 0.225 ns | 1.184 ns | 2.668 ns |
| Samples | 3 | 49.379 *** | 6.104 * | 11.683 ** | 75.79 *** | 16.133 ** | 18.968 ** | 6.914 * | 15.083 ** |
| Season | 3 | 5.497 * | 7.774 * | 5.13 * | 4.73 * | 0.563 ns | 2.871 ns | 2.095 ns | 10.033 ** |
| Species × Samples | 6 | 0.432 ns | 0.704 ns | 3.315 * | 0.581 ns | 1.337 ns | 2.139 ns | 3.112 * | 3.83 * |
| Species × Season | 6 | 1.391 ns | 0.338 ns | 1.55 ns | 0.909 ns | 0.777 ns | 1.631 ns | 2.075 ns | 2.854 * |
| Samples × Season | 9 | 3.226 * | 1.246 ns | 1.202 ns | 3.087 * | 4.82 ** | 5.036 ** | 4.002 ** | 3.759 * |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lv, X.; Hao, J.; Zhao, Y.; Li, C.; Quan, W. Seasonal Variations of Low-Molecular-Weight Organic Acids in Three Evergreen Broadleaf Rhododendron Forests. Metabolites 2023, 13, 119. https://doi.org/10.3390/metabo13010119
Lv X, Hao J, Zhao Y, Li C, Quan W. Seasonal Variations of Low-Molecular-Weight Organic Acids in Three Evergreen Broadleaf Rhododendron Forests. Metabolites. 2023; 13(1):119. https://doi.org/10.3390/metabo13010119
Chicago/Turabian StyleLv, Xiaofei, Jiangtao Hao, Yumeng Zhao, Chaochan Li, and Wenxuan Quan. 2023. "Seasonal Variations of Low-Molecular-Weight Organic Acids in Three Evergreen Broadleaf Rhododendron Forests" Metabolites 13, no. 1: 119. https://doi.org/10.3390/metabo13010119
APA StyleLv, X., Hao, J., Zhao, Y., Li, C., & Quan, W. (2023). Seasonal Variations of Low-Molecular-Weight Organic Acids in Three Evergreen Broadleaf Rhododendron Forests. Metabolites, 13(1), 119. https://doi.org/10.3390/metabo13010119