

Training-Induced Increase in

Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Theoretical Methods
2.1. Ethical Approval
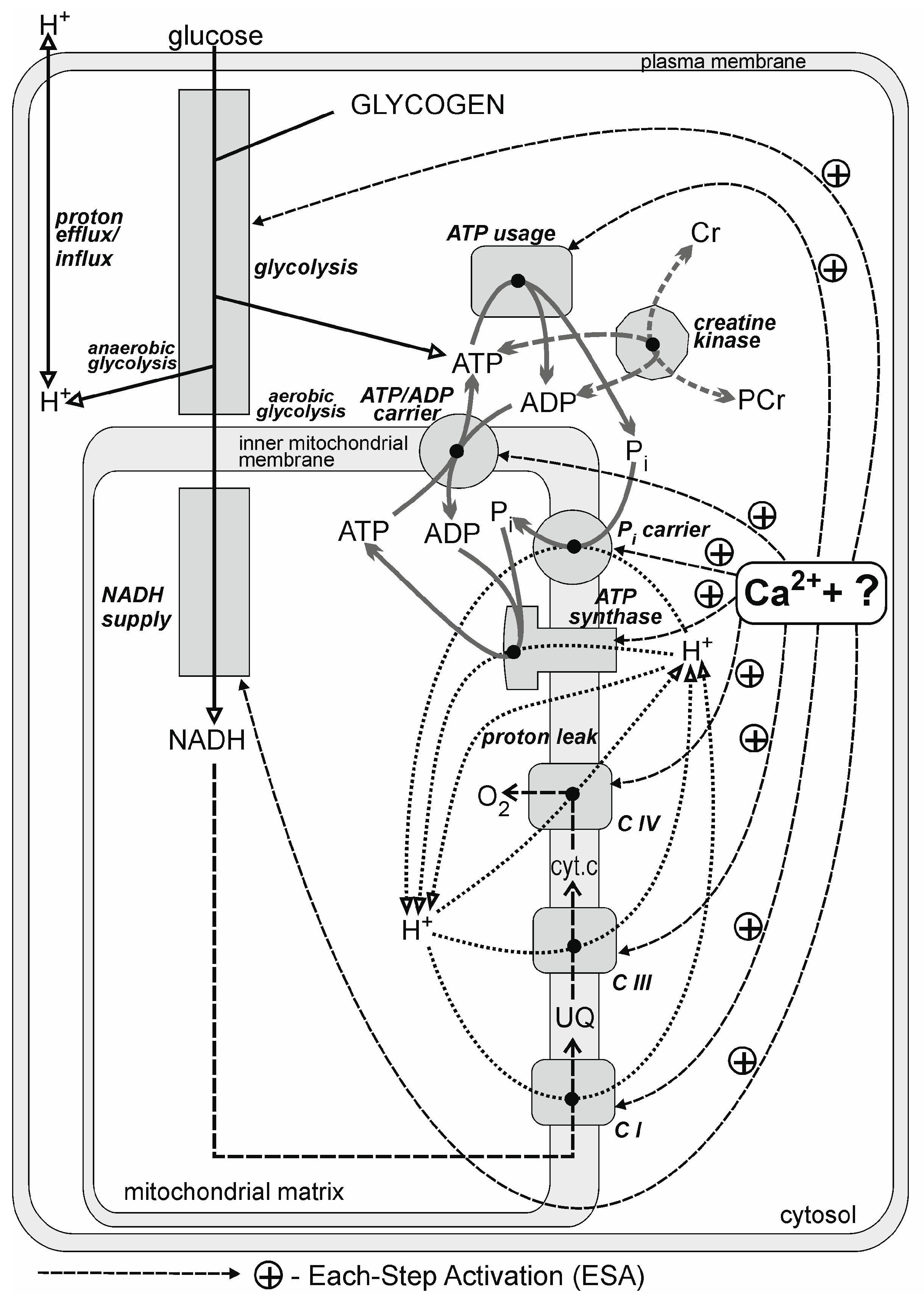
2.2. Computer Model
2.3. Bioenergetic Molecular Sequence of Events during Rest-to-Work Transition
2.4. Computer Simulations
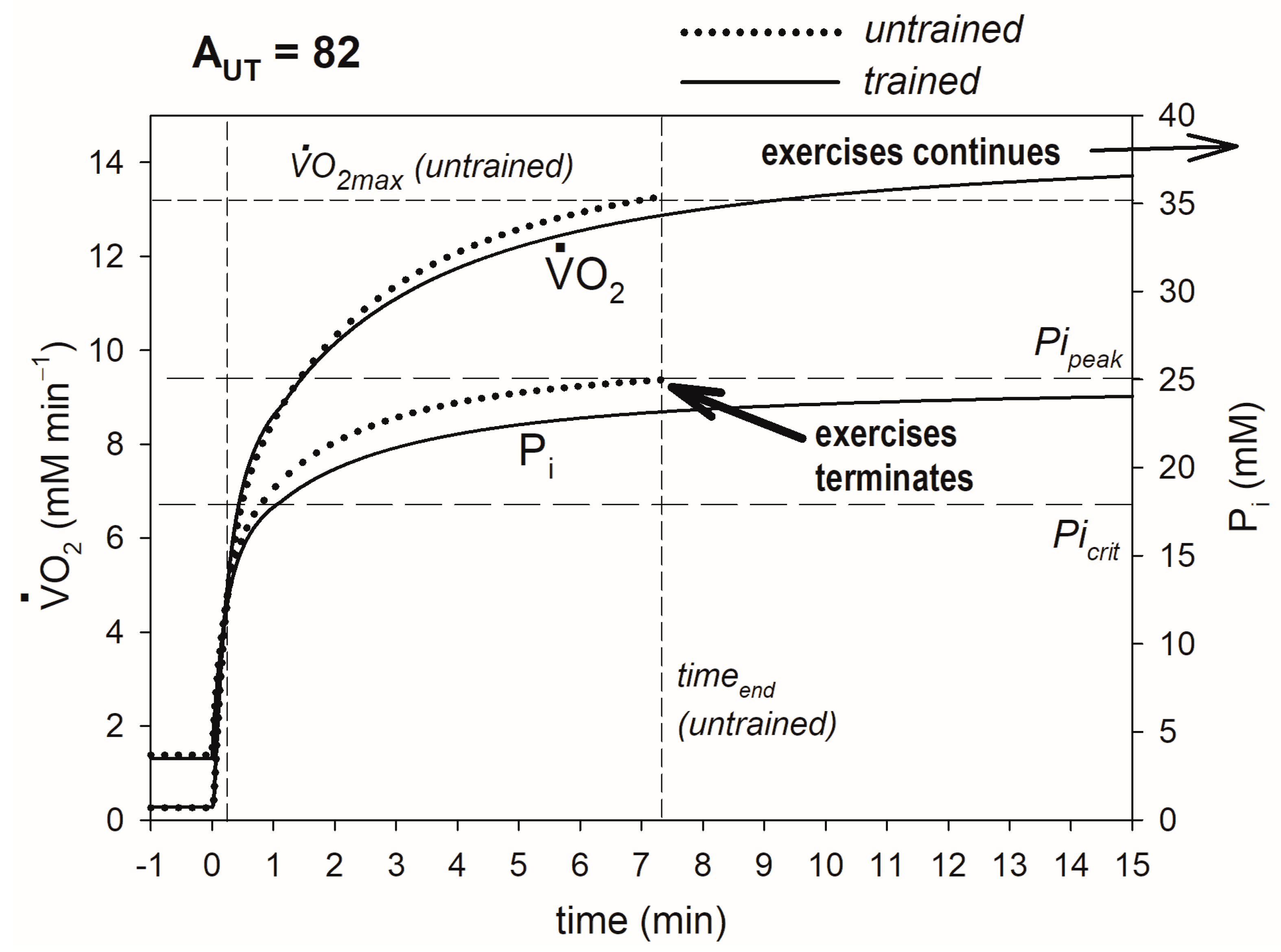
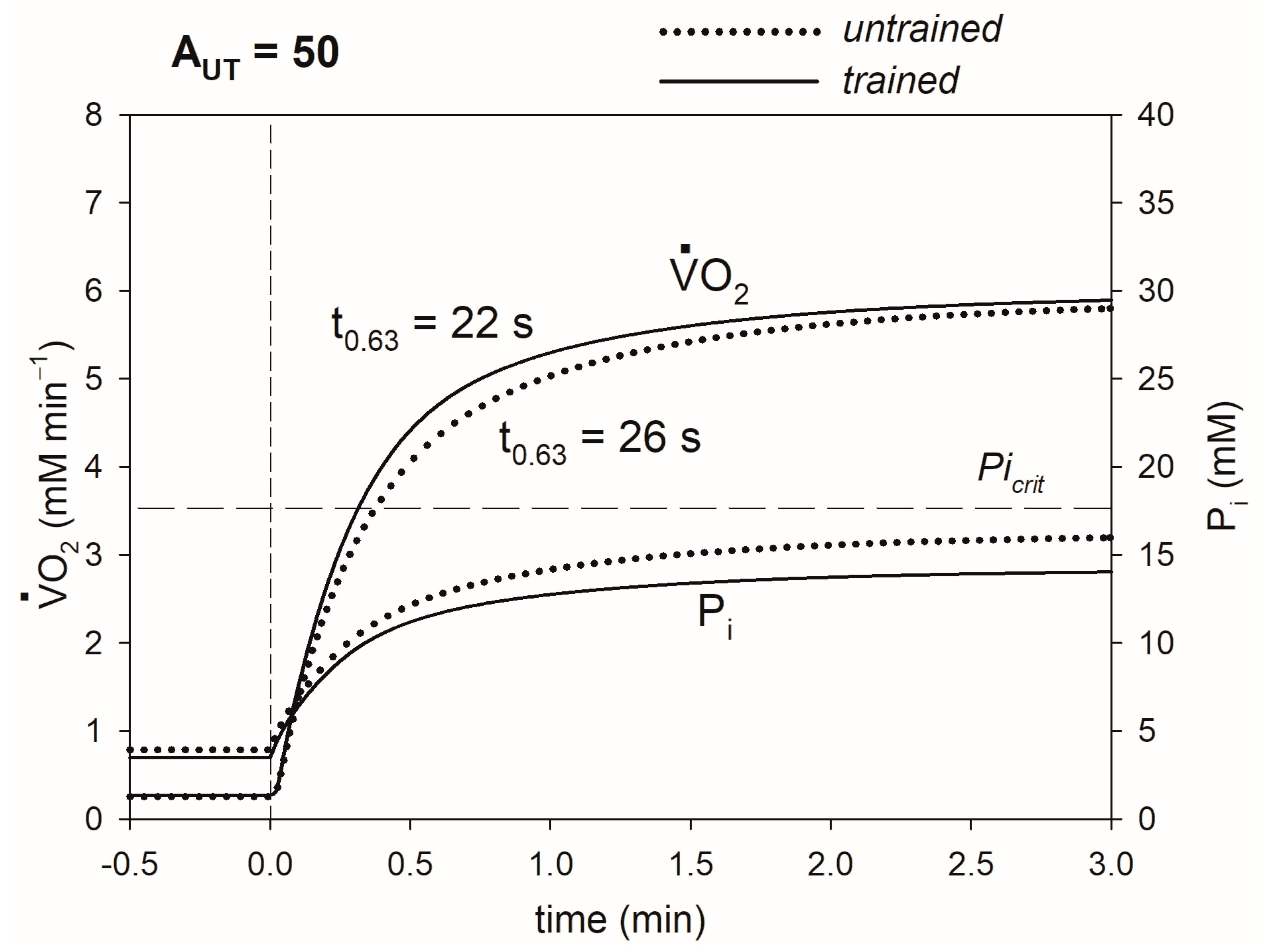
3. Results
4. Discussion
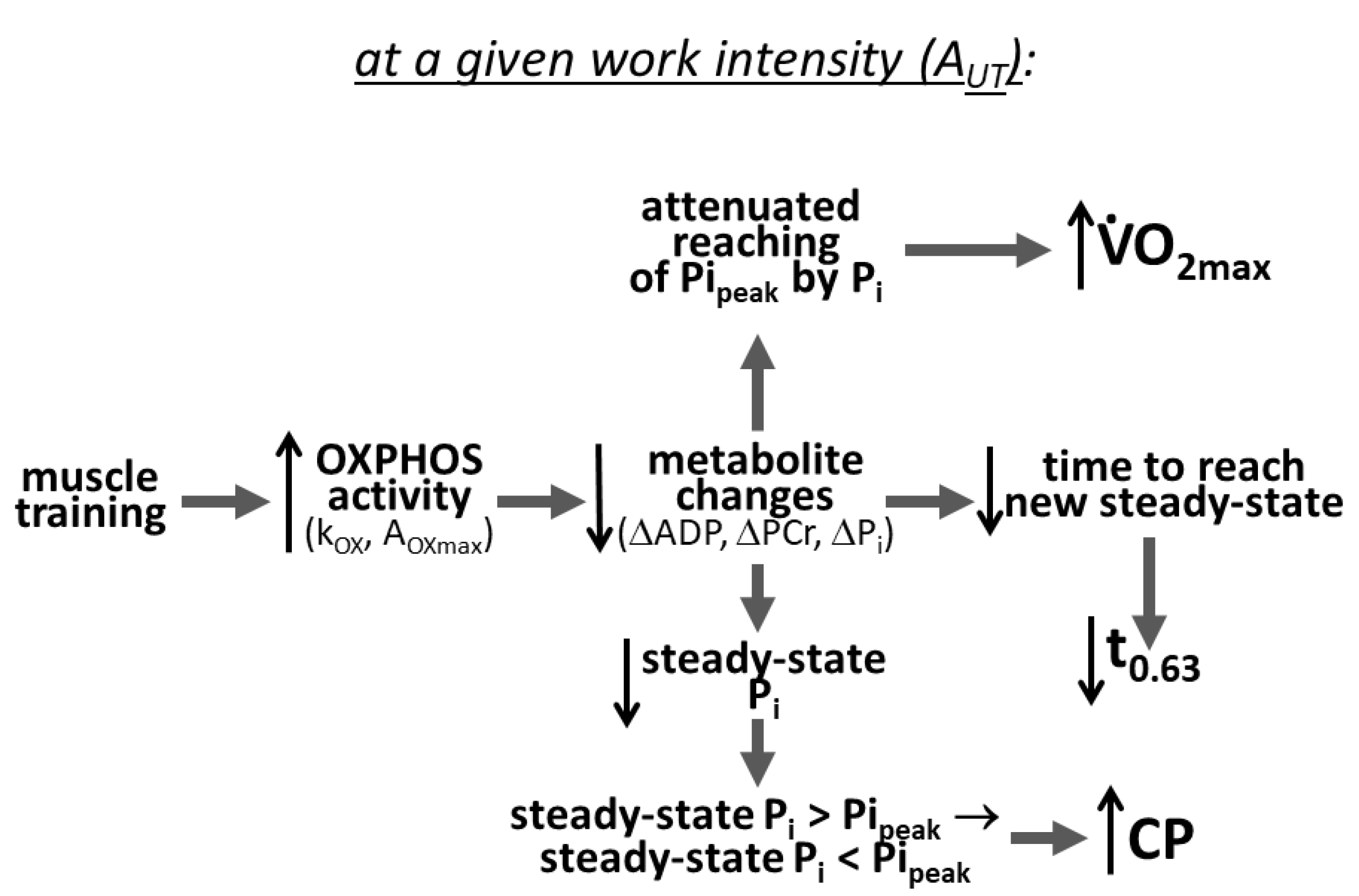
4.1. Mechanism of the Impact of Training-Induced Increase in OXPHOS Activity and Decrease in Pipeak on O2max, CP and t0.63
4.2. General Discussion
4.3. Study Limitations
5. Conclusions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hoppeler, H.; Howald, H.; Conley, K.; Lindsted, S.L.; Claassen, H.; Vock, P.; Weibel, E.R. Endurance training in humans: Aerobic capacity and structure of skeletal muscle. J. Appl. Physiol. 1985, 59, 320–327. [Google Scholar] [CrossRef]
- Casaburi, R.; Storer, T.W.; Ben-Dov, I.; Wasserman, K. Effect of endurance training on possible determinants of VO2 during heavy exercise. J. Appl. Physiol. 1987, 62, 199–207. [Google Scholar] [CrossRef]
- Gaesser, G.A.; Wilson, L.A. Effects of continuous and interval training on the parameters of the power-endurance time relationship for high-intensity exercise. Int. J. Sports Med. 1988, 9, 417–421. [Google Scholar] [CrossRef]
- Poole, D.C.; Ward, S.A.; Whipp, B.J. The effects of training on the metabolic and respiratory profile of high-intensity cycle ergometer exercise. Eur. J. Appl. Physiol. 1990, 59, 421–429. [Google Scholar] [CrossRef]
- Jenkins, D.G.; Quigley, B.M. Endurance training enhances critical power. Med. Sci. Sports Exerc. 1992, 2, 1283–1289. [Google Scholar] [CrossRef]
- Roca, J.; Agusti, A.G.N.; Alonso, A.; Poole, D.C.; Viegasl, C.; Barbera, J.A.; Rodrigez-Roisin, R.; Ferrer, A.; Wagner, P.D. Effects of training on muscle O2 transport at VO2max. J. Appl. Physiol. 1992, 73, 1067–1076. [Google Scholar] [CrossRef]
- Phillips, S.M.; Green, H.J.; MacDonald, M.J.; Hughson, R.L. Progressive effect of endurance training on VO2 kinetics at the onset of submaximal exercise. J. Appl. Physiol. 1995, 79, 1914–1920. [Google Scholar] [CrossRef]
- Suter, E.; Hoppeler, H.; Claassen, H. Ultrastructural modification of human skeletal-muscle tissue with 6-month moderate-intensity exercise training. Int. J. Sports Med. 1995, 16, 160–166. [Google Scholar] [CrossRef]
- Carter, H.; Jones, A.M.; Barstow, T.J.; Burnley, M.; Williams, C.; Doustl, J.H. Effect of endurance training: On oxygen uptake kinetics during treadmill running. J. Appl. Physiol. 2000, 89, 1744–1752. [Google Scholar] [CrossRef]
- Fernström, M.; Tonkonogil, M.; Sahlin, K. Effects of acute and chronic endurance exercise on mitochondrial uncoupling in human skeletal muscle. J. Physiol. 2004, 554, 755–763. [Google Scholar] [CrossRef]
- Berger, N.J.; Tolfrey, K.; Williams, A.G.; Jones, A.M. Influence of continuous and interval training on oxygen uptake on-kinetics. Med. Sci. Sports Exerc. 2006, 38, 504–512. [Google Scholar] [CrossRef]
- Hottenrott, K.; Ludyga, S.; Schulze, S. Effects of high intensity training and continuous endurance training on aerobic capacity and body composition in recreationally active runners. J. Sports Sci. Med. 2012, 11, 483–488. [Google Scholar]
- Zoladz, J.A.; Grassi, B.; Majerczak, J.; Szkutnik, Z.; Korostyński, M.; Karasiński, J.; Kilarski, W.; Korzeniewski, B. Training-induced acceleration of O2 uptake on-kinetics precedes mitochondria biogenesis in humans. Exp. Physiol. 2013, 98, 883–898. [Google Scholar] [CrossRef]
- Zoladz, J.A.; Grassi, B.; Majerczak, J.; Szkutnik, Z.; Korostyński, M.; Grandys, M.; Jarmuszkiewicz, W.; Korzeniewski, B. Mechanisms responsible for the acceleration of pulmonary VO2 on-kinetics in humans after prolonged endurance training. Am. J. Physiol. 2014, 307, R1101–R1114. [Google Scholar]
- Burnley, M.; Jones, A.M. Oxygen uptake kinetics as a determinant of sports performance. Eur. J. Sport Sci. 2007, 7, 63–79. [Google Scholar] [CrossRef]
- Womack, C.J.; Davis, S.E.; Blumer, J.L.; Barrett, E.; Weltman, A.L.; Gaesser, G.A. Slow component of O2 uptake during heavy exercise: Adaptation to endurance training. J. Appl. Physiol. 1995, 79, 838–845. [Google Scholar] [CrossRef]
- Holloszy, J.O. Biochemical adaptations in muscle. Effects of exercise on mitochondrial oxygen uptake and respiratory enzyme activity in skeletal muscle. J. Biol. Chem. 1967, 242, 2278–2282. [Google Scholar] [CrossRef]
- Scalzo, R.L.; Peltonen, G.L.; Binns, S.E.; Shankaran, M.; Giordano, G.R.; Hartley, D.A.; Klochak, A.L.; Lonac, M.C.; Paris, H.L.; Szallar, S.E.; et al. Greater muscle protein synthesis and mitochondrial biogenesis in males compared with females during sprint interval training. FASEB J. 2014, 28, 2705–2714. [Google Scholar] [CrossRef]
- Baldwin, K.M.; Klinkerfuss, G.H.; Terjung, R.L.; Molé, P.A.; Holloszy, J.O. Respiratory capacity of white, red, and intermediate muscle: Adaptative response to exercise. Am. J. Physiol. 1972, 222, 373–378. [Google Scholar] [CrossRef]
- Jacobs, R.A.; Flückl, D.; Bonne, T.C.; Bürgi, S.; Christensen, P.M.; Toigo, M.; Lundby, C. Improvements in exercise performance with high-intensity interval training coincide with an increase in skeletal muscle mitochondrial content and function. J. Appl. Physiol. 2013, 115, 785–793. [Google Scholar] [CrossRef]
- Pesta, D.; Hoppel, F.; Macek, C.; Messner, H.; Faulhaber, M.; Kobel, C.; Parson, W.; Burtscher, M.; Schocke, M.; Gnaiger, E. Similar qualitative and quantitative changes of mitochondrial respiration following strength and endurance training in normoxia and hypoxia in sedentary humans. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2011, 301, R1078–R1087. [Google Scholar] [CrossRef]
- Wibom, R.; Hultman, E.; Johansson, M.; Matherei, K.; Constantin-Teodosiu, D.; Schantz, P.G. Adaptation of mitochondrial ATP production in human skeletal muscle to endurance training and detraining. J. Appl. Physiol. 1992, 73, 2004–2010. [Google Scholar] [CrossRef]
- Tarnopolsky, M.A.; Rennie, C.D.; Robertshaw, H.A.; Fedak-Tarnopolsky, S.N.; Devries, M.C.; Hamadeh, M.J. Influence of endurance exercise training and sex on intramyocellular lipid and mitochondrial ultrastructure, substrate use, and mitochondrial enzyme activity. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2007, 292, R1271–R1278. [Google Scholar] [CrossRef]
- Zoladz, J.A.; Majerczak, J.; Glganski, L.; Grandys, M.; Zapart-Bukowska, J.; Kuczek, P.; Kołodziejski, L.; Walkowicz, L.; Szymoniak-Chochół, D.; Kilarski, W.; et al. Endurance training increases the running performance of untrained men without changing the mitochondrial volume density in the gastrocnemius muscle. Int. J. Mol. Sci. 2022, 23, 10843. [Google Scholar] [CrossRef]
- Krieger, D.A.; Tate, C.A.; McMillin-Wood, J.; Booth, F.W. Populations of rat skeletal muscle mitochondria after exercise and immobilization. J. Appl. Physiol. 1980, 48, 23–28. [Google Scholar] [CrossRef]
- McCully, K.K.; Boden, B.P.; Tuchler, M.; Fountain, M.R.; Chance, B. Wrist flexor muscles of elite rowers measured with magnetic resonance spectroscopy. J. Appl. Physiol. 1989, 67, 926–932. [Google Scholar] [CrossRef]
- Sundberg, C.W.; Prost, R.W.; Fitts, R.H.; Hunter, S.K. Bioenergetic basis for the increased fatigability with ageing. J. Physiol. 2019, 597, 4943–4957. [Google Scholar] [CrossRef]
- Korzeniewski, B.; Rossiter, H.B. Exceeding a “critical” Pi: Implications for VO2 and metabolite slow components, muscle fatigue and the power-duration relationship. Eur. J. Appl. Physiol. 2020, 120, 1609–1619. [Google Scholar] [CrossRef]
- Korzeniewski, B.; Rossiter, H.B. Factors determining training-induced changes in VO2max, critical power and VO2 on-kinetics in skeletal muscle. J. Appl. Physiol. 2021, 130, 498–507. [Google Scholar] [CrossRef]
- Korzeniewski, B. Pi-induced muscle fatigue leads to near-hyperbolic power–duration dependence. Eur. J. Appl. Physiol. 2019, 119, 2201–2213. [Google Scholar] [CrossRef]
- Korzeniewski, B.; Zoladz, J.A. Possible factors determining the non-linearity in the VO2-power output relationship in humans theoretical studies. Japan. J. Physiol. 2003, 53, 271–280. [Google Scholar] [CrossRef] [PubMed]
- Korzeniewski, B. Mechanisms of the effect of oxidative phosphorylation deficiencies on the skeletal muscle bioenergetic system. J. Appl. Physiol. 2021, 131, 768–777. [Google Scholar] [CrossRef]
- Korzeniewski, B. Effect of training on skeletal muscle bioenergetic system in patients with mitochondrial myopathies: A computational study. Resp. Physiol. Neurobiol. 2022, 296, 103799. [Google Scholar] [CrossRef] [PubMed]
- Korzeniewski, B.; Zoladz, J.A. A model of oxidative phosphorylation in mammalian skeletal muscle. Biophys. Chem. 2001, 92, 17–34. [Google Scholar] [CrossRef] [PubMed]
- Korzeniewski, B.; Liguzinski, P. Theoretical studies on the regulation of anaerobic glycolysis and its influence on oxidative phosphorylation in skeletal muscle. Biophys. Chem. 2004, 110, 147–169. [Google Scholar] [CrossRef]
- Korzeniewski, B.; Rossiter, H.B. Each-step activation of oxidative phosphorylation is necessary to explain muscle metabolite kinetic responses to exercise and recovery in humans. J. Physiol. 2015, 593, 5255–5268. [Google Scholar] [CrossRef]
- Korzeniewski, B. Regulation of oxidative phosphorylation is different in electrically- and cortically-stimulated skeletal muscle. PLoS ONE 2018, 13, e0195620. [Google Scholar] [CrossRef]
- Korzeniewski, B. Muscle VO2-power output nonlinearity in constant-power, step-incremental, and ramp-incremental exercise: Magnitude and underlying mechanisms. Physiol. Rep. 2018, 6, e13915. [Google Scholar] [CrossRef]
- Korzeniewski, B. Regulation of ATP supply during muscle contraction: Theoretical studies. Biochem. J. 1998, 330, 1189–1195. [Google Scholar] [CrossRef]
- Korzeniewski, B. Regulation of oxidative phosphorylation in different muscles and various experimental conditions. Biochem. J. 2003, 375, 799–804. [Google Scholar] [CrossRef]
- Korzeniewski, B. Regulation of oxidative phosphorylation through parallel activation. Biophys. Chem. 2007, 129, 93–110. [Google Scholar] [CrossRef]
- Korzeniewski, B. Regulation of oxidative phosphorylation through each-step activation (ESA): Evidences from computer modeling. Prog. Biophys. Mol. Biol. 2017, 125, 1–23. [Google Scholar] [CrossRef] [PubMed]
- Fell, D.A.; Thomas, S. Physiological control of metabolic flux: The requirement for multisite modulation. Biochem. J. 1995, 311, 35–39. [Google Scholar] [CrossRef] [PubMed]
- Thomas, S.; Fell, D.A. The role of multiple enzyme activation in metabolic flux control. Adv. Enzyme. Regul. 1998, 38, 65–85. [Google Scholar] [CrossRef] [PubMed]
- Korzeniewski, B.; Rossiter, H.B. Skeletal muscle biochemical origin of exercise intensity domains and their relation to whole-body VO2 kinetics. Biosci. Rep. 2022, 42, BSR20220798. [Google Scholar] [CrossRef] [PubMed]
- Korzeniewski, B. Sensitivity of VO2max, critical power and VO2 on-kinetics to O2 concentration/delivery and other factors in skeletal muscle. Resp. Physiol. Neurobiol. 2023, 307, 103977. [Google Scholar] [CrossRef]
- Whipp, H.B. Domains of aerobic function and their limiting parameters. In The Physiology and Pathophysiology of Exercise Tolerance; Steinacker, J.M., Ward, S.A., Eds.; Plenum: New York, NY, USA, 1996; pp. 83–89. [Google Scholar]
- Easterby, J.S. A generalized theory of the transition time for sequential enzyme reactions. Biochem. J. 1981, 199, 155–161. [Google Scholar] [CrossRef]
- Korzeniewski, B.; Zoladz, J.A. Factors determining the oxygen consumption rate (VO2) on-kinetics in skeletal muscle. Biochem. J. 2004, 379, 703–710. [Google Scholar] [CrossRef]
- Goulding, R.P.; Rossiter, H.B.; Marwood, S.; Ferguson, C. Bioenergetic mechanisms linking VO2 kinetics and exercise tolerance. Exerc. Sport Sci. Rev. 2021, 49, 274–283. [Google Scholar] [CrossRef]
- Allen, D.G.; Lamb, G.D.; Westerblad, H. Skeletal muscle fatigue: Cellular mechanisms. Physiol. Rev. 2008, 88, 287–332. [Google Scholar] [CrossRef]
- Wilson, J.R.; McCully, K.K.; Mancini, D.M.; Boden, B.; Chance, B. Relationship of muscular fatigue to pH and diprotonated Pi in humans: A 31P-NMR study. J. Appl. Physiol. 1988, 64, 2333–2339. [Google Scholar] [CrossRef] [PubMed]
- Allen, D.G.; Westerblad, H. Role of phosphate and calcium stores in muscle fatigue. J. Physiol. 2001, 536, 657–665. [Google Scholar] [CrossRef] [PubMed]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Korzeniewski, B.
Training-Induced Increase in
Korzeniewski B.
Training-Induced Increase in
Korzeniewski, Bernard.
2023. "Training-Induced Increase in
Korzeniewski, B.
(2023). Training-Induced Increase in