
The Cerebellum’s Role in Affective Disorders: The Onset of Its Social Dimension



Abstract
:
{kind=link}
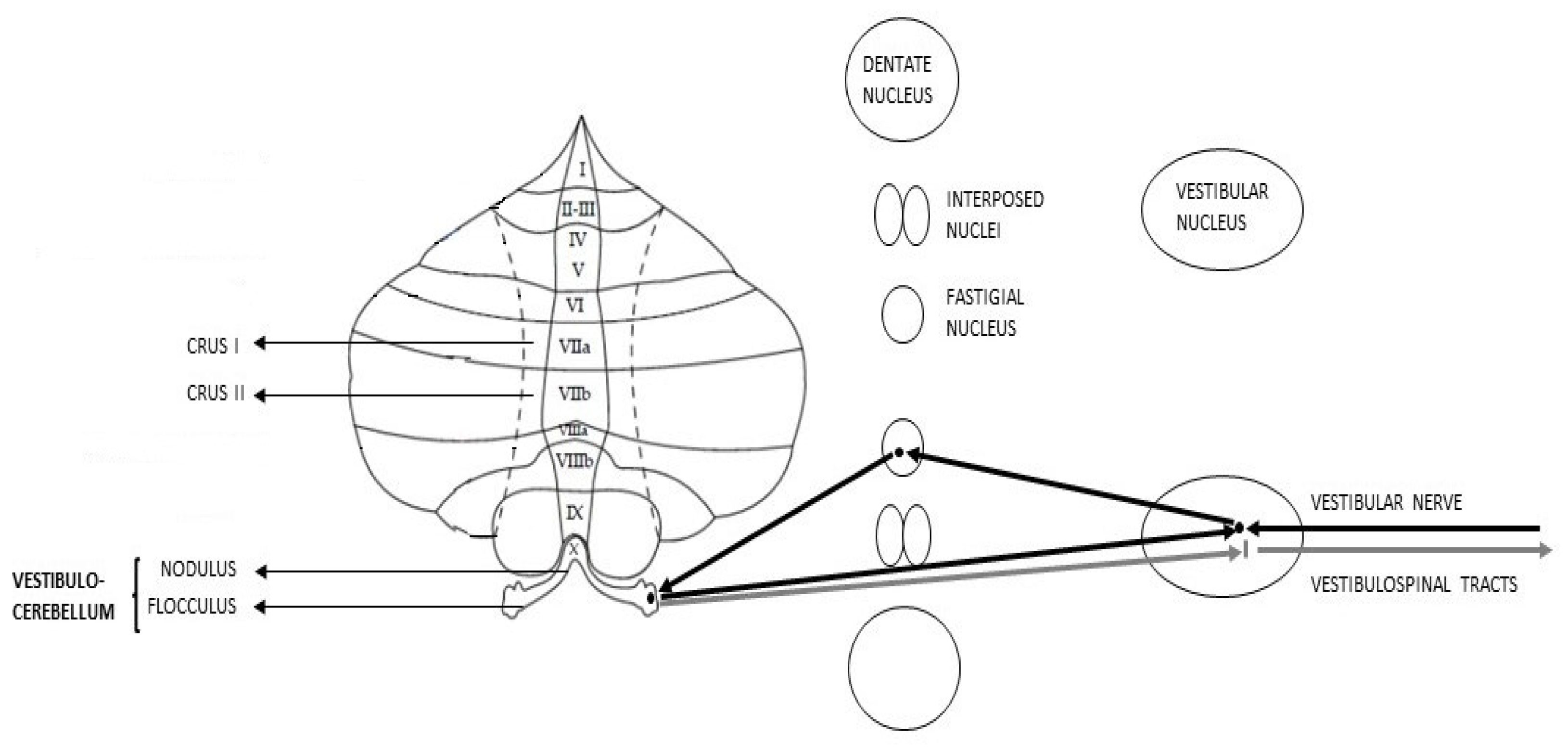{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Methods
3. Affective Disorders and Cerebellum
4. Vestibulocerebellum
5. Pontocerebellum
6. Spinocerebellum
7. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| Abbreviation | Definition | Abbreviation | Definition |
| MDD | Major Depressive Disorder | Thl | Thalamus |
| BD | Bipolar Disorder | Hyp | Hypothalamus |
| Scz | Schizophrenia | DN | Dentate Nucleus |
| Cbm | Cerebellum | FN | Fastigial Nucleus |
| vCbm | Vestibulocerebellum | LS | Limbic System |
| pCbm | Pontocerebellum | DA | Dopamine |
| sCbm | Spinocerebellum | Glu | Glutamate |
| Str | Striatum | 5-HT | Serotonin |
| NA | Nucleus Accumbens | NE | Norepinephrine |
| Amg | Amygdala | GABA | Gamma-AminoButyric Acid |
| Hip | Hippocampus | fMRI | Functional MRI |
| PFC | Prefrontal Cortex | rsfMRI | Resting State fMRI |
References
- Wen, M.; Dong, Z.; Zhang, L.; Li, B.; Zhang, Y.; Li, K. Depression and Cognitive Impairment: Current Understanding of Its Neurobiology and Diagnosis. Neuropsychiatr. Dis. Treat. 2022, 18, 2783–2794. [Google Scholar] [CrossRef] [PubMed]
- American Psychiatric Association. Diagnostic and Statistical Manual of Mental Disorders, 5th ed.; American Psychiatric Association: Arlington, VA, USA, 2013. [Google Scholar] [CrossRef]
- Vieta, E.; Berk, M.; Schulze, T.G.; Carvalho, A.F.; Suppes, T.; Calabrese, J.R.; Gao, K.; Miskowiak, K.W.; Grande, I. Bipolar disorders. Nat. Rev. Dis. Prim. 2018, 4, 18008. [Google Scholar] [CrossRef]
- Lee, M.T.; Peng, W.-H.; Kan, H.-W.; Wu, C.-C.; Wang, D.-W.; Ho, Y.-C. Neurobiology of Depression: Chronic Stress Alters the Glutamatergic System in the Brain—Focusing on AMPA Receptor. Biomedicines 2022, 10, 1005. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Y.; Zou, D.; Li, Y.; Gu, S.; Dong, J.; Ma, X.; Xu, S.; Wang, F.; Huang, J.H. Monoamine Neurotransmitters Control Basic Emotions and Affect Major Depressive Disorders. Pharmaceuticals 2022, 15, 1203. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Cui, M.; Cao, J.-L.; Han, M.-H. The Role of Beta-Adrenergic Receptors in Depression and Resilience. Biomedicines 2022, 10, 2378. [Google Scholar] [CrossRef] [PubMed]
- Sarawagi, A.; Soni, N.D.; Patel, A.B. Glutamate and GABA Homeostasis and Neurometabolism in Major Depressive Disorder. Front. Psychiatry 2021, 12, 637863. [Google Scholar] [CrossRef]
- Mandal, P.K.; Gaur, S.; Roy, R.G.; Samkaria, A.; Ingole, R.; Goel, A. Schizophrenia, Bipolar and Major Depressive Disorders: Overview of Clinical Features, Neurotransmitter Alterations, Pharmacological Interventions, and Impact of Oxidative Stress in the Disease Process. ACS Chem. Neurosci. 2022, 13, 2784–2802. [Google Scholar] [CrossRef]
- Adamaszek, M.; D’agata, F.; Ferrucci, R.; Habas, C.; Keulen, S.; Kirkby, K.C.; Leggio, M.; Mariën, P.; Molinari, M.; Moulton, E.; et al. Consensus Paper: Cerebellum and Emotion. Cerebellum 2017, 16, 552–576. [Google Scholar] [CrossRef]
- Wang, S.; Leri, F.; Rizvi, S.J. Anhedonia as a central factor in depression: Neural mechanisms revealed from preclinical to clinical evidence. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2021, 110, 110289. [Google Scholar] [CrossRef]
- Li, Y.; Zhang, B.; Pan, X.; Wang, Y.; Xu, X.; Wang, R.; Liu, Z. Dopamine-Mediated Major Depressive Disorder in the Neural Circuit of Ventral Tegmental Area-Nucleus Accumbens-Medial Prefrontal Cortex: From Biological Evidence to Computational Models. Front. Cell. Neurosci. 2022, 16, 923039. [Google Scholar] [CrossRef]
- Nakamura, Y.; Ishida, T.; Tanaka, S.C.; Mitsuyama, Y.; Yokoyama, S.; Shinzato, H.; Itai, E.; Okada, G.; Kobayashi, Y.; Kawashima, T.; et al. Distinctive alterations in the mesocorticolimbic circuits in various psychiatric disorders. Psychiatry Clin. Neurosci. 2023, 77, 345–354. [Google Scholar] [CrossRef] [PubMed]
- Delva, N.C.; Stanwood, G.D. Dysregulation of brain dopamine systems in major depressive disorder. Exp. Biol. Med. 2021, 246, 1084–1093. [Google Scholar] [CrossRef] [PubMed]
- Bhatt, S.; Devadoss, T.; Manjula, S.N.; Rajangam, J. 5-HT3 Receptor Antagonism: A Potential Therapeutic Approach for the Treatment of Depression and other Disorders. Curr. Neuropharmacol. 2021, 19, 1545–1559. [Google Scholar] [CrossRef] [PubMed]
- Lan, M.J.; Zanderigo, F.; Pantazatos, S.P.; Sublette, M.E.; Miller, J.; Ogden, R.T.; Mann, J.J. Serotonin 1A Receptor Binding of [11C]CUMI-101 in Bipolar Depression Quantified Using Positron Emission Tomography: Relationship to Psychopathology and Antidepressant Response. Int. J. Neuropsychopharmacol. 2022, 25, 534–544. [Google Scholar] [CrossRef] [PubMed]
- Helton, S.G.; Lohoff, F.W. Serotonin pathway polymorphisms and the treatment of major depressive disorder and anxiety disorders. Pharmacogenomics 2015, 16, 541–553. [Google Scholar] [CrossRef]
- Bartlett, E.A.; Zanderigo, F.; Shieh, D.; Miller, J.; Hurley, P.; Rubin-Falcone, H.; Oquendo, M.A.; Sublette, M.E.; Ogden, R.T.; Mann, J.J. Serotonin transporter binding in major depressive disorder: Impact of serotonin system anatomy. Mol. Psychiatry 2022, 27, 3417–3424. [Google Scholar] [CrossRef]
- Kantrowitz, J.T.; Dong, Z.; Milak, M.S.; Rashid, R.; Kegeles, L.S.; Javitt, D.C.; Lieberman, J.A.; Mann, J.J. Ventromedial prefrontal cortex/anterior cingulate cortex Glx, glutamate, and GABA levels in medication-free major depressive disorder. Transl. Psychiatry 2021, 11, 419. [Google Scholar] [CrossRef]
- Kong, L.; Li, H.; Lin, F.; Zheng, W.; Zhang, H.; Wu, R. Neurochemical and microstructural alterations in bipolar and depressive disorders: A multimodal magnetic resonance imaging study. Front. Neurol. 2023, 14, 1089067. [Google Scholar] [CrossRef]
- Maloku, E.; Covelo, I.R.; Hanbauer, I.; Guidotti, A.; Kadriu, B.; Hu, Q.; Davis, J.M.; Costa, E. Lower number of cerebellar Purkinje neurons in psychosis is associated with reduced reelin expression. Proc. Natl. Acad. Sci. USA 2010, 107, 4407–4411. [Google Scholar] [CrossRef]
- Fatemi, S.H.; Stary, J.M.; Earle, J.A.; Araghi-Niknam, M.; Eagan, E. GABAergic dysfunction in schizophrenia and mood disorders as reflected by decreased levels of glutamic acid decarboxylase 65 and 67 kDa and Reelin proteins in cerebellum. Schizophr. Res. 2005, 72, 109–122. [Google Scholar] [CrossRef]
- Xu, S.; Liu, Y.; Pu, J.; Gui, S.; Zhong, X.; Tian, L.; Song, X.; Qi, X.; Wang, H.; Xie, P. Chronic Stress in a Rat Model of Depression Disturbs the Glutamine–Glutamate–GABA Cycle in the Striatum, Hippocampus, and Cerebellum. Neuropsychiatr. Dis. Treat. 2020, 16, 557–570. [Google Scholar] [CrossRef]
- Blatt, G.L.; Oblak, A.L.; Schmahmann, J.D. Cerebellar connections with limbic circuits: Anatomy and functional impli-cations. In Handbook of the Cerebellum and Cerebellar Disorders; Springer: Dordrecht, The Netherlands, 2013; pp. 479–496. [Google Scholar] [CrossRef]
- Roy, A.K.; Shehzad, Z.; Margulies, D.S.; Kelly, A.C.; Uddin, L.Q.; Gotimer, K.; Biswal, B.B.; Castellanos, F.X.; Milham, M.P. Functional connectivity of the human amygdala using resting state fMRI. NeuroImage 2009, 45, 614–626. [Google Scholar] [CrossRef]
- Jung, J.-Y.; Cho, S.-E.; Kim, N.; Kang, C.-K.; Kang, S.-G. Decreased resting-state functional connectivity of the habenula-cerebellar in a major depressive disorder. Front. Psychiatry 2022, 13, 925823. [Google Scholar] [CrossRef]
- Heath, R.G. Modulation of Emotion with A Brain Pacemamer. Treatment for Intractable Psychiatric Illness. J. Nerv. Ment. Dis. 1977, 165, 300–317. Available online: https://pubmed.ncbi.nlm.nih.gov/303280/ (accessed on 1 April 2023). [CrossRef] [PubMed]
- Heath, R.G.; Llewellyn, R.C.; Rouchell, A.M. The Cerebellar Pacemaker for Intractable Behavioral Disorders and Epilepsy: Follow-Up Report. Biol. Psychiatry 1980, 15, 243–256. Available online: http://www.ncbi.nlm.nih.gov/pubmed/7417614 (accessed on 15 December 2022).
- Heath, R.G.; Rouchell, A.M.; Llewellyn, R.C.; Walker, C.F. Cerebellar Pacemaker Patients: An Update. Biol. Psychiatry 1981, 16, 953–962. Available online: http://www.ncbi.nlm.nih.gov/pubmed/7306618 (accessed on 15 December 2022). [PubMed]
- Turner, R.N.; Hewstone, M.; Voci, A. Reducing explicit and implicit outgroup prejudice via direct and extended contact: The mediating role of self-disclosure and intergroup anxiety. J. Pers. Soc. Psychol. 2007, 93, 369–388. [Google Scholar] [CrossRef] [PubMed]
- Schutter, D.J.L.G.; Van Honk, J. The cerebellum on the rise in human emotion. Cerebellum 2005, 4, 290–294. [Google Scholar] [CrossRef]
- Habas, C.; Kamdar, N.; Nguyen, D.; Prater, K.; Beckmann, C.F.; Menon, V.; Greicius, M.D. Distinct Cerebellar Contributions to Intrinsic Connectivity Networks. J. Neurosci. 2009, 29, 8586–8594. [Google Scholar] [CrossRef]
- D’Angelo, E.; Casali, S. Seeking a unified framework for cerebellar function and dysfunction: From circuit operations to cognition. Front. Neural Circuits 2012, 6, 116. [Google Scholar] [CrossRef]
- Çavdar, S.; Güneş, Y.C.; Algın, O. Connections of the Dentate Nucleus with the Amygdala: Experimental Rat and Human 3-Tesla Tractography Study. Brain Connect. 2022, 12, 905–913. [Google Scholar] [CrossRef] [PubMed]
- Allen, G.; McColl, R.; Barnard, H.; Ringe, W.K.; Fleckenstein, J.; Cullum, C.M. Magnetic resonance imaging of cerebellar–prefrontal and cerebellar–parietal functional connectivity. NeuroImage 2005, 28, 39–48. [Google Scholar] [CrossRef]
- Yu, M.; Wang, S.-M. Neuroanatomy, Nucleus Fastigial. 2023. Available online: https://pubmed.ncbi.nlm.nih.gov/31613518/ (accessed on 1 April 2023).
- Hallett, M. Parkinson’s disease tremor: Pathophysiology. Park. Relat. Disord. 2012, 18, S85–S86. [Google Scholar] [CrossRef] [PubMed]
- Wu, T.; Hallett, M. The cerebellum in Parkinson’s disease. Brain 2013, 136, 696–709. [Google Scholar] [CrossRef] [PubMed]
- Lewis, J.D.; Theilmann, R.J.; Townsend, J.; Evans, A.C. Network efficiency in autism spectrum disorder and its relation to brain overgrowth. Front. Hum. Neurosci. 2013, 7, 845. [Google Scholar] [CrossRef]
- Lefaivre, S.C.; Brown, M.J.N.; Almeida, Q.J. Cerebellar involvement in Parkinson’s disease resting tremor. Cerebellum Ataxias 2016, 3, 13. [Google Scholar] [CrossRef]
- Schmahmann, J.D.; Sherman, J.C. The cerebellar cognitive affective syndrome. Brain 1998, 121, 561–579. [Google Scholar] [CrossRef]
- Benagiano, V.; Rizzi, A.; Lorusso, L.; Flace, P.; Saccia, M.; Cagiano, R.; Ribatti, D.; Roncali, L.; Ambrosi, G. The functional anatomy of the cerebrocerebellar circuit: A review and new concepts. J. Comp. Neurol. 2018, 526, 769–789. [Google Scholar] [CrossRef]
- Alalade, E.; Denny, K.; Potter, G.; Steffens, D.; Wang, L. Altered Cerebellar-Cerebral Functional Connectivity in Geriatric Depression. PLoS ONE 2011, 6, e20035. [Google Scholar] [CrossRef] [PubMed]
- Minichino, A.; Bersani, F.S.; Trabucchi, G.; Albano, G.; Primavera, M.; Chiaie, R.D.; Biondi, M. The role of cerebellum in unipolar and bipolar depression: A review of the main neurobiological findings. Riv. Psichiatr. 2014, 49, 124–131. [Google Scholar] [CrossRef]
- Tanaka, H.; Ishikawa, T.; Lee, J.; Kakei, S. The Cerebro-Cerebellum as a Locus of Forward Model: A Review. Front. Syst. Neurosci. 2020, 14, 19. [Google Scholar] [CrossRef]
- Stoodley, C.J.; Schmahmann, J.D. Evidence for topographic organization in the cerebellum of motor control versus cognitive and affective processing. Cortex 2010, 46, 831–844. [Google Scholar] [CrossRef] [PubMed]
- Sang, L.; Qin, W.; Liu, Y.; Han, W.; Zhang, Y.; Jiang, T.; Yu, C. Resting-state functional connectivity of the vermal and hemispheric subregions of the cerebellum with both the cerebral cortical networks and subcortical structures. NeuroImage 2012, 61, 1213–1225. [Google Scholar] [CrossRef] [PubMed]
- Beyer, J.L.; Krishnan, K.R.R. Volumetric brain imaging findings in mood disorders. Bipolar Disord. 2002, 4, 89–104. [Google Scholar] [CrossRef]
- Liotti, M.; Mayberg, H.S.; Brannan, S.K.; McGinnis, S.; Jerabek, P.; Fox, P.T. Differential limbic–cortical correlates of sadness and anxiety in healthy subjects: Implications for affective disorders. Biol. Psychiatry 2000, 48, 30–42. [Google Scholar] [CrossRef] [PubMed]
- Strata, P.; Provini, L.; Redman, S. On the concept of spinocerebellum. Proc. Natl. Acad. Sci. USA 2012, 109, E622. [Google Scholar] [CrossRef]
- Baldaçara, L.; Borgio, J.G.F.; de Lacerda, A.L.T.; Jackowski, A.P. Cerebellum and psychiatric disorders. Braz. J. Psychiatry 2008, 30, 281–289. [Google Scholar] [CrossRef]
- Hilber, P. The Role of the Cerebellar and Vestibular Networks in Anxiety Disorders and Depression: The Internal Model Hypothesis. Cerebellum 2022, 21, 791–800. [Google Scholar] [CrossRef]
- Lopez, C. A neuroscientific account of how vestibular disorders impair bodily self-consciousness. Front. Integr. Neurosci. 2013, 7, 91. [Google Scholar] [CrossRef]
- Cullen, K.E. The vestibular system: Multimodal integration and encoding of self-motion for motor control. Trends Neurosci. 2012, 35, 185–196. [Google Scholar] [CrossRef]
- Indovina, I.; Riccelli, R.; Staab, J.P.; Lacquaniti, F.; Passamonti, L. Personality traits modulate subcortical and cortical vestibular and anxiety responses to sound-evoked otolithic receptor stimulation. J. Psychosom. Res. 2014, 77, 391–400. [Google Scholar] [CrossRef]
- Coelho, C.M.; Balaban, C.D. Visuo-vestibular contributions to anxiety and fear. Neurosci. Biobehav. Rev. 2015, 48, 148–159. [Google Scholar] [CrossRef]
- Balaban, C.D. Projections from the parabrachial nucleus to the vestibular nuclei: Potential substrates for autonomic and limbic influences on vestibular responses. Brain Res. 2004, 996, 126–137. [Google Scholar] [CrossRef]
- Tovote, P.; Fadok, J.P.; Lüthi, A. Neuronal circuits for fear and anxiety. Nat. Rev. Neurosci. 2015, 16, 317–331. [Google Scholar] [CrossRef] [PubMed]
- Strata, P. The Emotional Cerebellum. Cerebellum 2015, 14, 570–577. [Google Scholar] [CrossRef] [PubMed]
- Sacchetti, B.; Sacco, T.; Strata, P. Reversible inactivation of amygdala and cerebellum but not perirhinal cortex impairs reactivated fear memories. Eur. J. Neurosci. 2007, 25, 2875–2884. [Google Scholar] [CrossRef]
- Sacchetti, B.; Baldi, E.; Lorenzini, C.A.; Bucherelli, C. Cerebellar role in fear-conditioning consolidation. Proc. Natl. Acad. Sci. USA 2002, 99, 8406–8411. [Google Scholar] [CrossRef]
- Chrobak, A.; Bohaterewicz, B.; Tereszko, A.; Krupa, A.; Sobczak, A.; Ceglarek, A.; Wielgus, M.; Fafrowicz, M.; Siwek, M.; Bryll, A.; et al. Altered functional connectivity among frontal eye fields, thalamus and cerebellum in bipolar disorder. Psychiatr. Pol. 2020, 54, 487–497. [Google Scholar] [CrossRef] [PubMed]
- Katayama, K.-I.; Morimura, N.; Kobayashi, K.; Corbett, D.; Okamoto, T.; Ornthanalai, V.G.; Matsunaga, H.; Fujita, W.; Matsumoto, Y.; Akagi, T.; et al. Slitrk2 deficiency causes hyperactivity with altered vestibular function and serotonergic dysregulation. iScience 2022, 25, 104604. [Google Scholar] [CrossRef]
- Rabellino, D.; Thome, J.; Densmore, M.; Théberge, J.; McKinnon, M.C.; Lanius, R.A. The Vestibulocerebellum and the Shattered Self: A Resting-State Functional Connectivity Study in Posttraumatic Stress Disorder and Its Dissociative Subtype. Cerebellum 2022, 1–15. [Google Scholar] [CrossRef]
- Koziol, L.F.; Budding, D.; Andreasen, N.; D’Arrigo, S.; Bulgheroni, S.; Imamizu, H.; Ito, M.; Manto, M.; Marvel, C.; Parker, K.; et al. Consensus Paper: The Cerebellum’s Role in Movement and Cognition. Cerebellum 2014, 13, 151–177. [Google Scholar] [CrossRef]
- Schmahmann, J.D. From movement to thought: Anatomic substrates of the cerebellar contribution to cognitive processing. Hum. Brain Mapp. 1996, 4, 174–198. [Google Scholar] [CrossRef]
- Buckner, R.L. The Cerebellum and Cognitive Function: 25 Years of Insight from Anatomy and Neuroimaging. Neuron 2013, 80, 807–815. [Google Scholar] [CrossRef]
- Bostan, A.C.; Dum, R.P.; Strick, P.L. Cerebellar networks with the cerebral cortex and basal ganglia. Trends Cogn. Sci. 2013, 17, 241–254. [Google Scholar] [CrossRef]
- Middleton, F.A.; Strick, P.L. Anatomical Evidence for Cerebellar and Basal Ganglia Involvement in Higher Cognitive Function. Science 1994, 266, 458–461. [Google Scholar] [CrossRef]
- Habas, C. Research note: A resting-state, cerebello-amygdaloid intrinsically connected network. Cerebellum Ataxias 2018, 5, 4. [Google Scholar] [CrossRef] [PubMed]
- Habas, C. Topography of Emotions in Cerebellum as Appraised by Functional Imaging. In The Emotional Cerebellum—Advances in Experimental Medicine and Biology; Adamaszek, M., Manto, M., Schutter, D.J.L.G., Eds.; Springer: Cham, Switzerland, 2022; pp. 77–86. [Google Scholar] [CrossRef]
- Habas, C. Functional Imaging of the Deep Cerebellar Nuclei: A Review. Cerebellum 2010, 9, 22–28. [Google Scholar] [CrossRef] [PubMed]
- Shinn, A.K.; Roh, Y.S.; Ravichandran, C.T.; Baker, J.T.; Öngür, D.; Cohen, B.M. Aberrant cerebellar connectivity in bipolar disorder with psychosis. Biol. Psychiatry Cogn. Neurosci. Neuroimaging 2017, 2, 438–448. [Google Scholar] [CrossRef]
- Schmahmann, J.D. The Role of the Cerebellum in Cognition and Emotion: Personal Reflections Since 1982 on the Dysmetria of Thought Hypothesis, and Its Historical Evolution from Theory to Therapy. Neuropsychol. Rev. 2010, 20, 236–260. [Google Scholar] [CrossRef]
- Jung, S.J.; Vlasov, K.; D’ambra, A.F.; Parigi, A.; Baya, M.; Frez, E.P.; Villalobos, J.; Fernandez-Frentzel, M.; Anguiano, M.; Ideguchi, Y.; et al. Novel Cerebello-Amygdala Connections Provide Missing Link between Cerebellum and Limbic System. Front. Syst. Neurosci. 2022, 16, 879634. [Google Scholar] [CrossRef]
- Guo, W.; Liu, F.; Zhang, Z.; Liu, J.; Yu, M.; Zhang, J.; Xiao, C.; Zhao, J. Unidirectionally affected causal connectivity of cortico-limbic-cerebellar circuit by structural deficits in drug-naive major depressive disorder. J. Affect. Disord. 2015, 172, 410–416. [Google Scholar] [CrossRef]
- Feng, L.; Wu, D.; Ma, S.; Dong, L.; Yue, Y.; Li, T.; Tang, Y.; Ye, Z.; Mao, G. Resting-state functional connectivity of the cerebellum-cerebrum in older women with depressive symptoms. BMC Psychiatry 2023, 23, 732. [Google Scholar] [CrossRef]
- Liu, L.-K.; Chou, K.-H.; Hsu, C.-C.H.; Peng, L.-N.; Lee, W.-J.; Chen, W.-T.; Lin, C.-P.; Chung, C.-P.; Wang, P.-N.; Chen, L.-K. Cerebellar-limbic neurocircuit is the novel biosignature of physio-cognitive decline syndrome. Aging 2020, 12, 25319–25336. [Google Scholar] [CrossRef]
- Hou, L.; Lam, B.Y.-H.; Wong, N.M.L.; Lu, W.; Zhang, R.; Ning, Y.; Lin, K. Integrity of cerebellar tracts associated with the risk of bipolar disorder. Transl. Psychiatry 2022, 12, 335. [Google Scholar] [CrossRef]
- Soontornniyomkij, B.; Everall, I.P.; Chana, G.; Tsuang, M.T.; Achim, C.L.; Soontornniyomkij, V. Tyrosine kinase B protein expression is reduced in the cerebellum of patients with bipolar disorder. J. Affect. Disord. 2011, 133, 646–654. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Luo, Z.; Qiu, S.; Jia, Y.; Zhong, S.; Chen, G.; Lai, S.; Qi, Z.; Luo, X.; Huang, G.; et al. Abnormalities of aquaporin-4 in the cerebellum in bipolar II disorder: An ultra-high b-values diffusion weighted imaging study. J. Affect. Disord. 2020, 274, 136–143. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Liu, H.; Tang, Y.; Yan, R.; Jiang, X.; Fan, G.; Sun, W. Decreased Functional Connectivity of Vermis-Ventral Prefrontal Cortex in Bipolar Disorder. Front. Hum. Neurosci. 2021, 15, 711688. [Google Scholar] [CrossRef] [PubMed]
- Saleem, A.; Harmata, G.; Jain, S.; Voss, M.W.; Fiedorowicz, J.G.; Williams, A.J.; Shaffer, J.J.; Richards, J.G.; Barsotti, E.J.; Sathyaputri, L.; et al. Functional Connectivity of the Cerebellar Vermis in Bipolar Disorder and Associations with Mood. Front. Psychiatry 2023, 14, 1147540. [Google Scholar] [CrossRef]
- Schmahmann, J.D.; Weilburg, J.B.; Sherman, J.C. The neuropsychiatry of the cerebellum—Insights from the clinic. Cerebellum 2007, 6, 254–267. [Google Scholar] [CrossRef]
- Depping, M.S.; Schmitgen, M.M.; Kubera, K.M.; Wolf, R.C. Cerebellar Contributions to Major Depression. Front. Psychiatry 2018, 9, 634. [Google Scholar] [CrossRef]
- Depping, M.S.; Schmitgen, M.M.; Bach, C.; Listunova, L.; Kienzle, J.; Kubera, K.M.; Roesch-Ely, D.; Wolf, R.C. Abnormal Cerebellar Volume in Patients with Remitted Major Depression with Persistent Cognitive Deficits. Cerebellum 2020, 19, 762–770. [Google Scholar] [CrossRef]
- Bench, C.J.; Friston, K.J.; Brown, R.G.; Scott, L.C.; Frackowiak, R.S.J.; Dolan, R.J.; Bench, C.J.; Friston, K.J.; Brown, R.G.; Scott, L.C.; et al. The anatomy of melancholia—Focal abnormalities of cerebral blood flow in major depression. Psychol. Med. 1992, 22, 607–615. [Google Scholar] [CrossRef]
- Amen, D.G.; Taylor, D.V.; Meysami, S.; Raji, C.A. Deficits in Regional Cerebral Blood Flow on Brain SPECT Predict Treatment Resistant Depression. J. Alzheimer’s Dis. 2018, 63, 529–538. [Google Scholar] [CrossRef] [PubMed]
- Dolan, R.J.; Bench, C.J.; Brown, R.G.; Scott, L.C.; Friston, K.J.; Frackowiak, R.S.; Dolan, R.J.; Bench, C.J.; Brown, R.G.; Scott, L.C.; et al. Regional cerebral blood flow abnormalities in depressed patients with cognitive impairment. J. Neurol. Neurosurg. Psychiatry 1992, 55, 768–773. [Google Scholar] [CrossRef] [PubMed]
- Su, L.; Cai, Y.; Xu, Y.; Dutt, A.; Shi, S.; Bramon, E. Cerebral metabolism in major depressive disorder: A voxel-based meta-analysis of positron emission tomography studies. BMC Psychiatry 2014, 14, 321. [Google Scholar] [CrossRef]
- Bogoian, H.R.; King, T.Z.; Turner, J.A.; Semmel, E.S.; Dotson, V.M. Linking depressive symptom dimensions to cerebellar subregion volumes in later life. Transl. Psychiatry 2020, 10, 201. [Google Scholar] [CrossRef] [PubMed]
- Strakowski, S.M.; DelBello, M.P.; Adler, C.M. The functional neuroanatomy of bipolar disorder: A review of neuroimaging findings. Mol. Psychiatry 2005, 10, 105–116. [Google Scholar] [CrossRef]
- Yucel, K.; Nazarov, A.; Taylor, V.H.; Macdonald, K.; Hall, G.B.; MacQueen, G.M. Cerebellar vermis volume in major depressive disorder. Brain Struct. Funct. 2013, 218, 851–858. [Google Scholar] [CrossRef]
- Lupo, M.; Olivito, G.; Siciliano, L.; Masciullo, M.; Molinari, M.; Cercignani, M.; Bozzali, M.; Leggio, M. Evidence of Cerebellar Involvement in the Onset of a Manic State. Front. Neurol. 2018, 9, 774. [Google Scholar] [CrossRef]
- Todeva-Radneva, A.; Kandilarova, S.; Paunova, R.; Stoyanov, D.; Zdravkova, T.; Sladky, R. Functional Connectivity of the Anterior Cingulate Cortex and the Right Anterior Insula Differentiates between Major Depressive Disorder, Bipolar Disorder and Healthy Controls. Biomedicines 2023, 11, 1608. [Google Scholar] [CrossRef] [PubMed]
- Fateh, A.A.; Cui, Q.; Duan, X.; Yang, Y.; Chen, Y.; Li, D.; He, Z.; Chen, H. Disrupted dynamic functional connectivity in right amygdalar subregions differentiates bipolar disorder from major depressive disorder. Psychiatry Res. Neuroimaging 2020, 304, 111149. [Google Scholar] [CrossRef]
- Singh, M.K.; Spielman, D.; Libby, A.; Adams, E.; Acquaye, T.; Howe, M.; Kelley, R.; Reiss, A.; Chang, K.D. Neurochemical deficits in the cerebellar vermis in child offspring of parents with bipolar disorder. Bipolar Disord. 2011, 13, 189–197. [Google Scholar] [CrossRef] [PubMed]
- Uwisengeyimana, J.d.D.; Nguchu, B.A.; Wang, Y.; Zhang, D.; Liu, Y.; Qiu, B.; Wang, X. Cognitive function and cerebellar morphometric changes relate to abnormal intra-cerebellar and cerebro-cerebellum functional connectivity in old adults. Exp. Gerontol. 2020, 140, 111060. [Google Scholar] [CrossRef] [PubMed]
- Yildiz-Yesiloglu, A.; Ankerst, D.P. Neurochemical alterations of the brain in bipolar disorder and their implications for pathophysiology: A systematic review of the in vivo proton magnetic resonance spectroscopy findings. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2006, 30, 969–995. [Google Scholar] [CrossRef] [PubMed]
- Akshoomoff, N.A.; Courchesne, E.; Townsend, J. Attention coordination and anticipatory control. Int. Rev. Neurobiol. 1997, 41, 575–584. [Google Scholar] [CrossRef]
- Vandervert, L. The prominent role of the cerebellum in the learning, origin and advancement of culture. Cerebellum Ataxias 2016, 3, 10. [Google Scholar] [CrossRef]
- Metoki, A.; Wang, Y.; Olson, I.R. The Social Cerebellum: A Large-Scale Investigation of Functional and Structural Specificity and Connectivity. Cereb. Cortex 2022, 32, 987–1003. [Google Scholar] [CrossRef]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Stanca, S.; Rossetti, M.; Bongioanni, P. The Cerebellum’s Role in Affective Disorders: The Onset of Its Social Dimension. Metabolites 2023, 13, 1113. https://doi.org/10.3390/metabo13111113
Stanca S, Rossetti M, Bongioanni P. The Cerebellum’s Role in Affective Disorders: The Onset of Its Social Dimension. Metabolites. 2023; 13(11):1113. https://doi.org/10.3390/metabo13111113
Chicago/Turabian StyleStanca, Stefano, Martina Rossetti, and Paolo Bongioanni. 2023. "The Cerebellum’s Role in Affective Disorders: The Onset of Its Social Dimension" Metabolites 13, no. 11: 1113. https://doi.org/10.3390/metabo13111113
APA StyleStanca, S., Rossetti, M., & Bongioanni, P. (2023). The Cerebellum’s Role in Affective Disorders: The Onset of Its Social Dimension. Metabolites, 13(11), 1113. https://doi.org/10.3390/metabo13111113