
Technical Report: A Comprehensive Comparison between Different Quantification Versions of Nightingale Health’s 1H-NMR Metabolomics Platform

 ,
,  , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Dataset Descriptions
2.1.1. BBMRI.NL
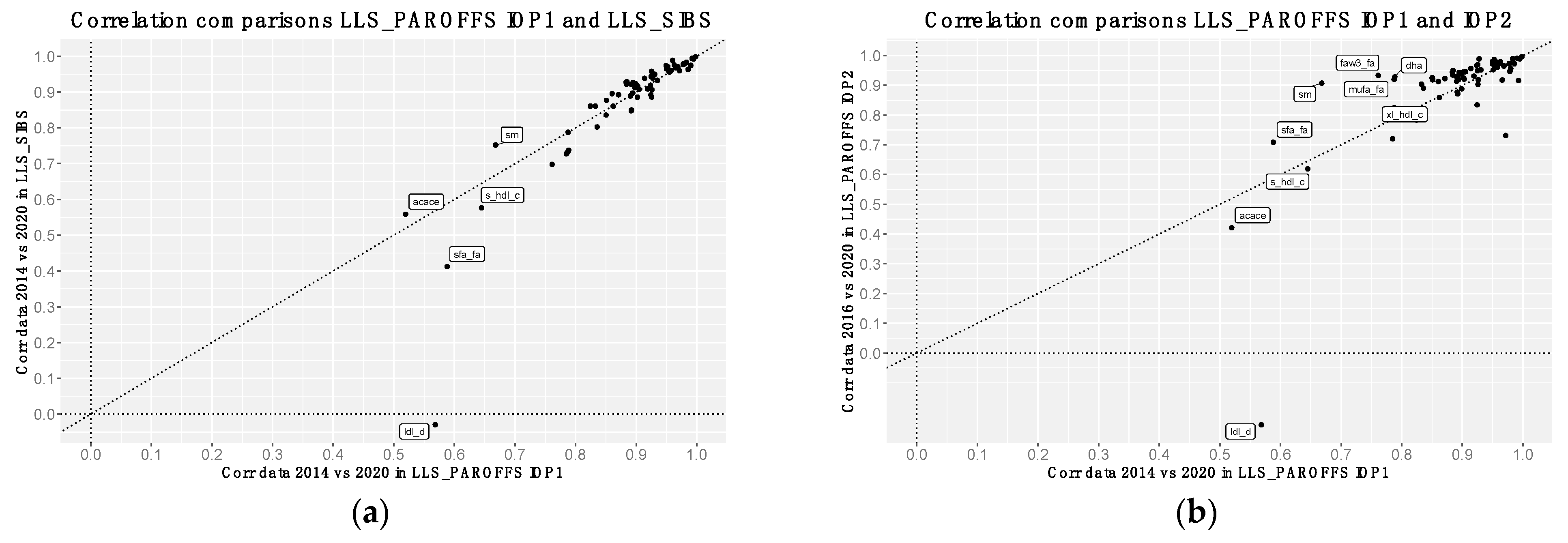
2.1.2. The Leiden Longevity Study
2.2. Comparison of the Metabolomic Analytes
2.3. MetaboHealth Score
2.4. MetaboAge
3. Results
3.1. An Overview of Changes in Measured Metabolic Features
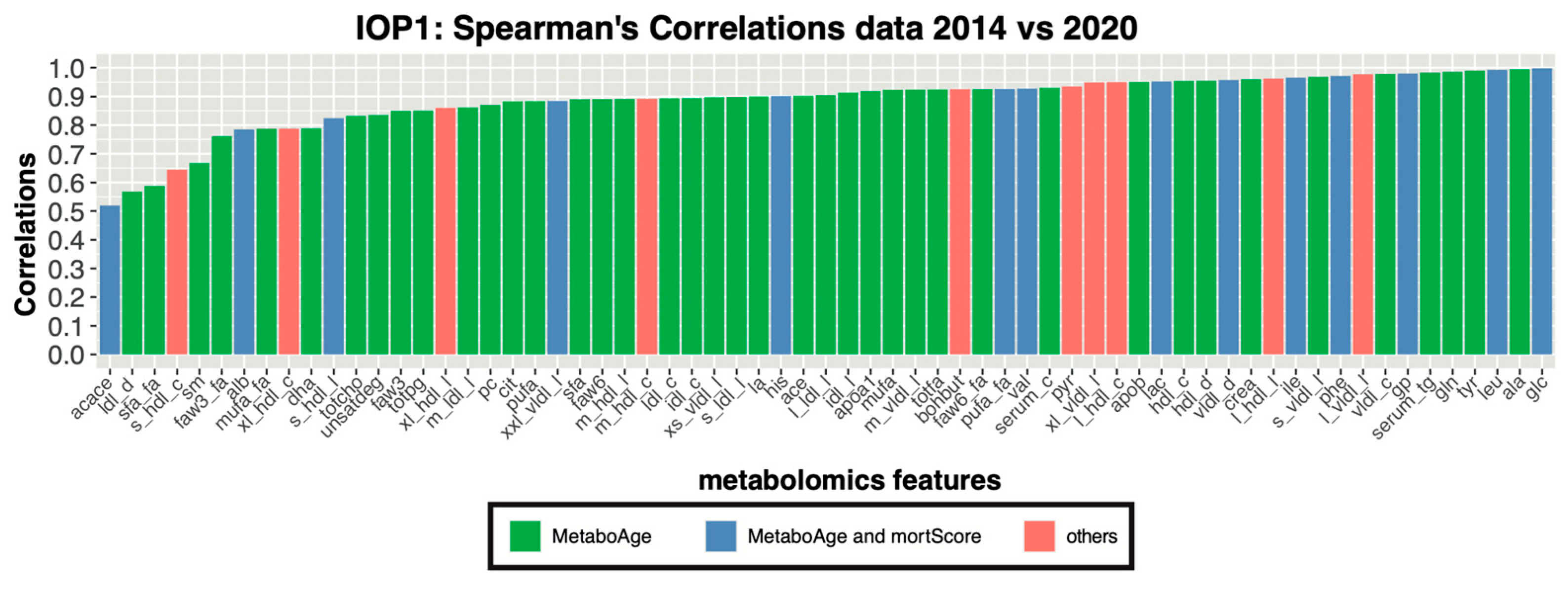
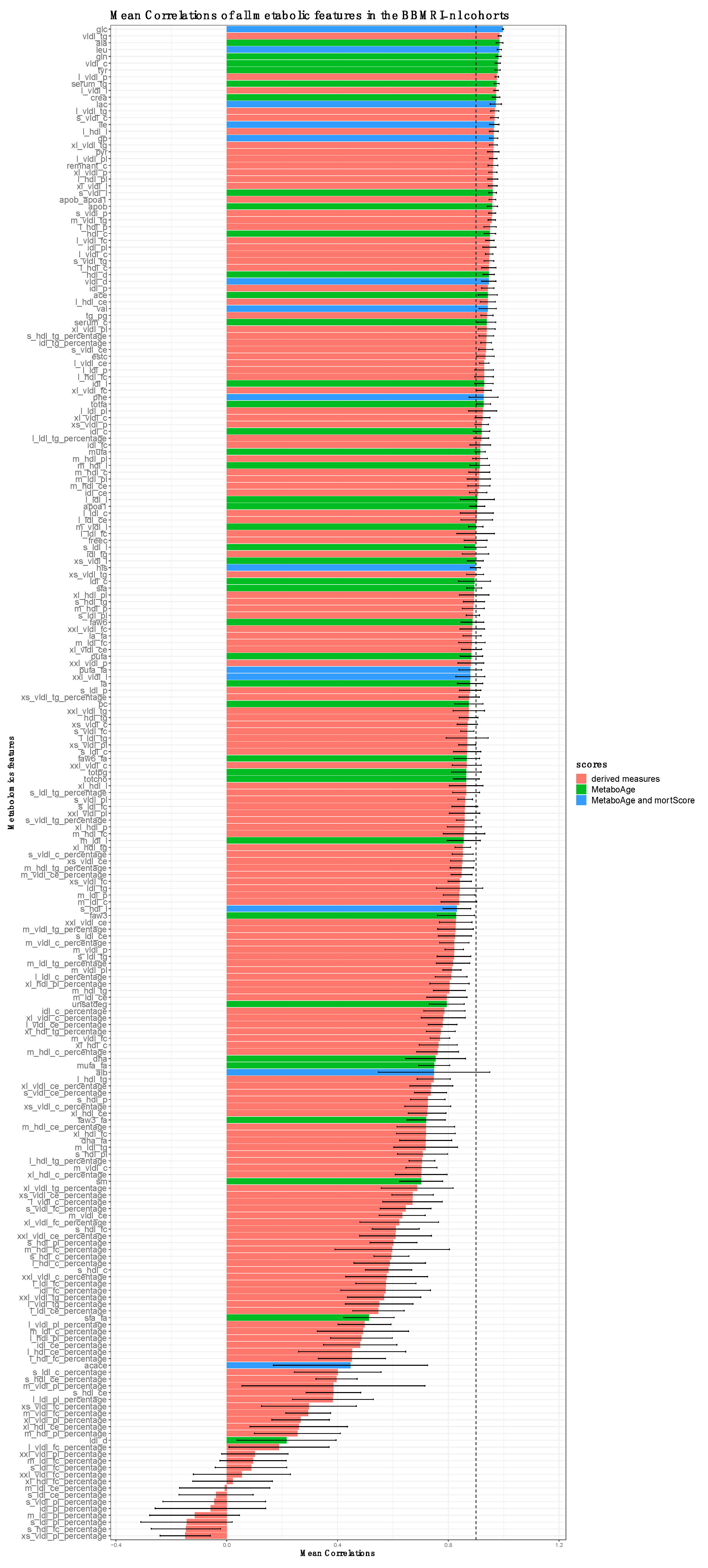
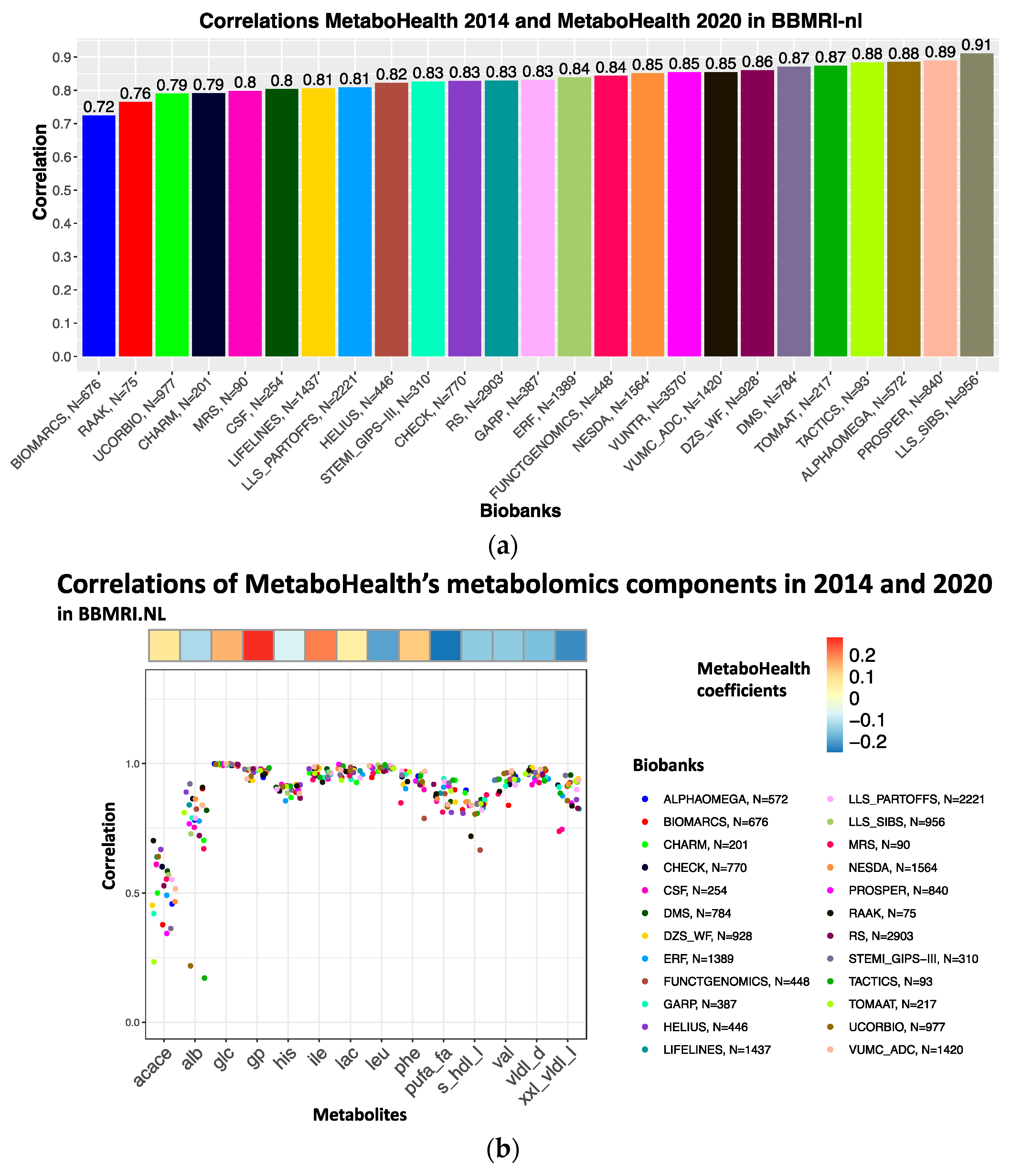
3.2. Correlation Analyses of Metabolomics Measurements between Platform Versions
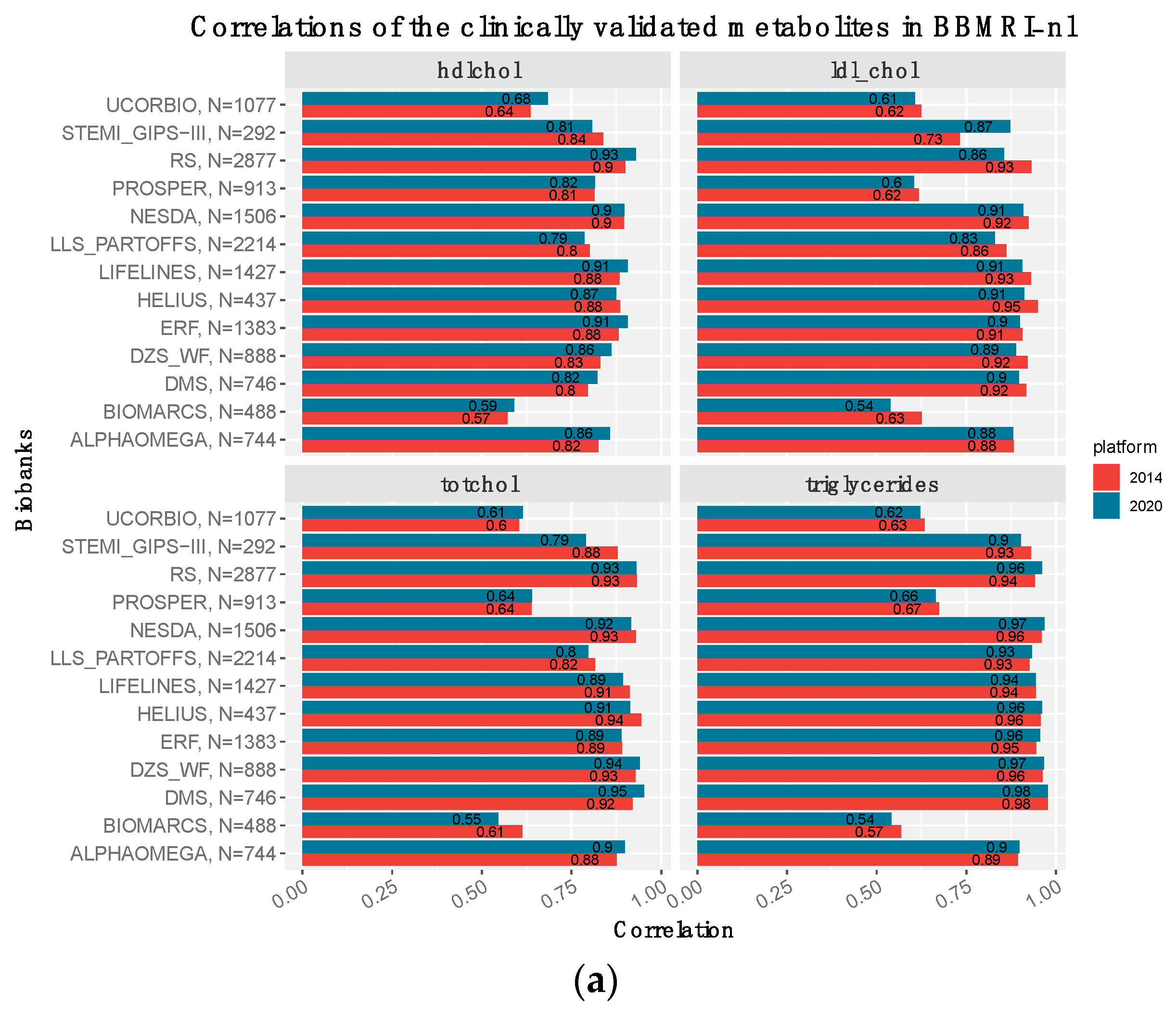
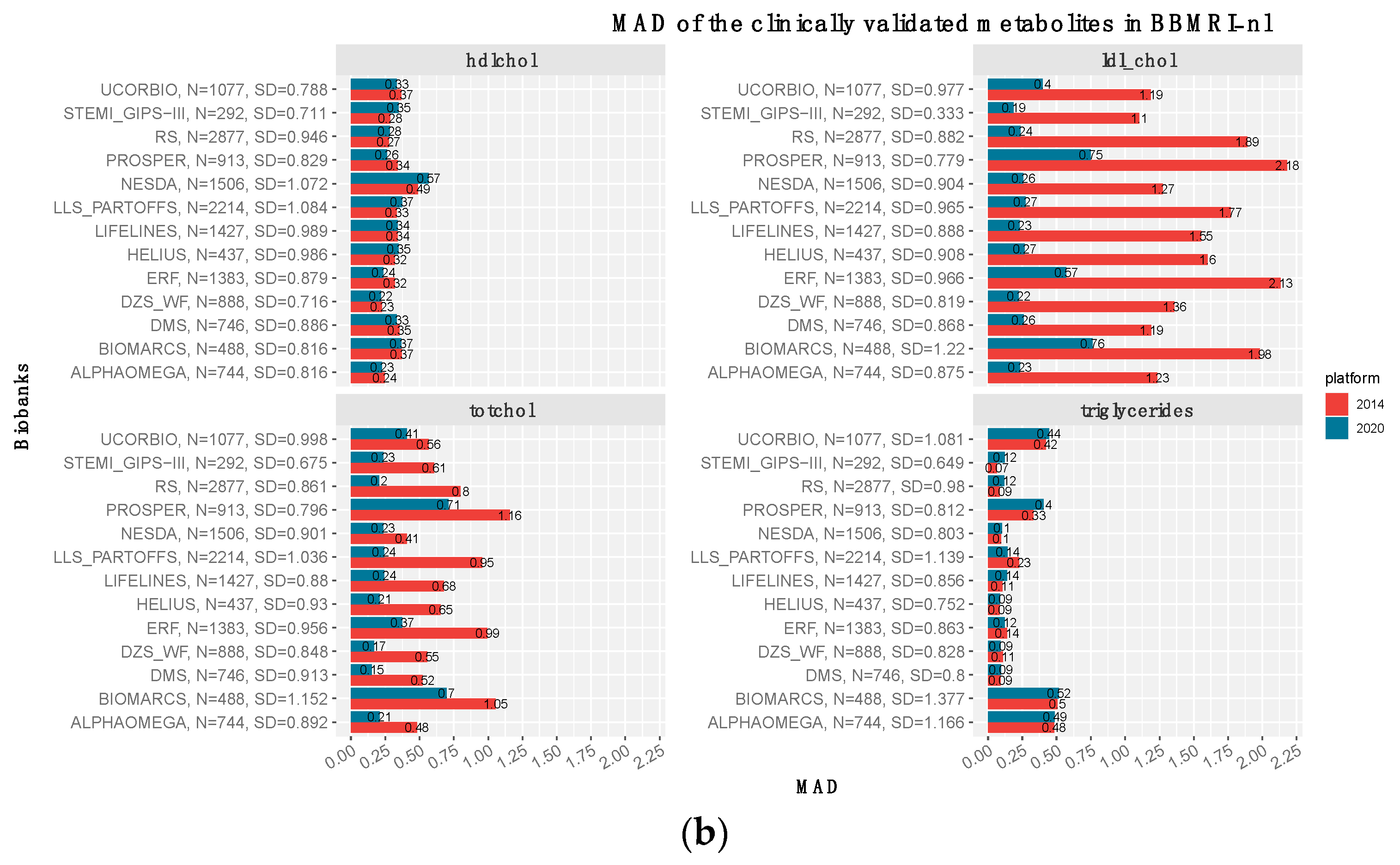
3.3. The Clinically Validated Biomarkers Show Similar Correlation, but Improved Calibration with Respect to Previous Quantification
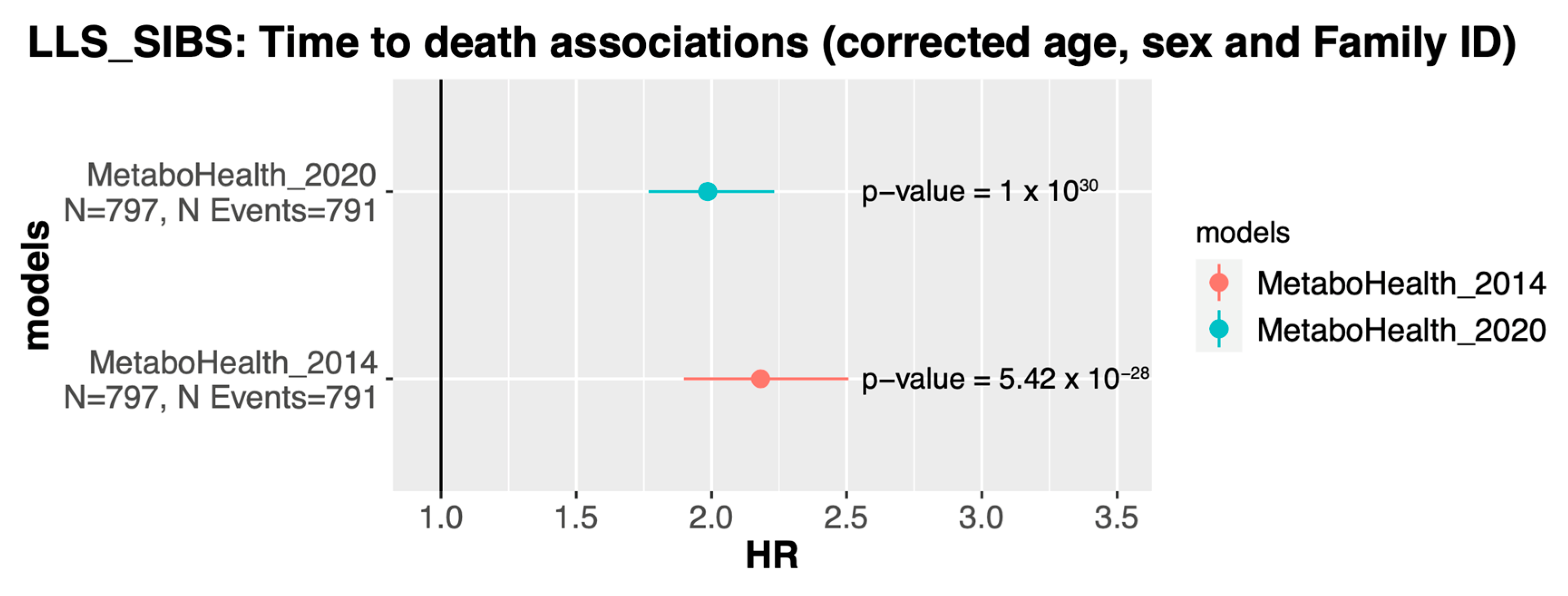
3.4. The MetaboHealth Score Shows a Comparable Association with Mortality Using Re-Quantified Data
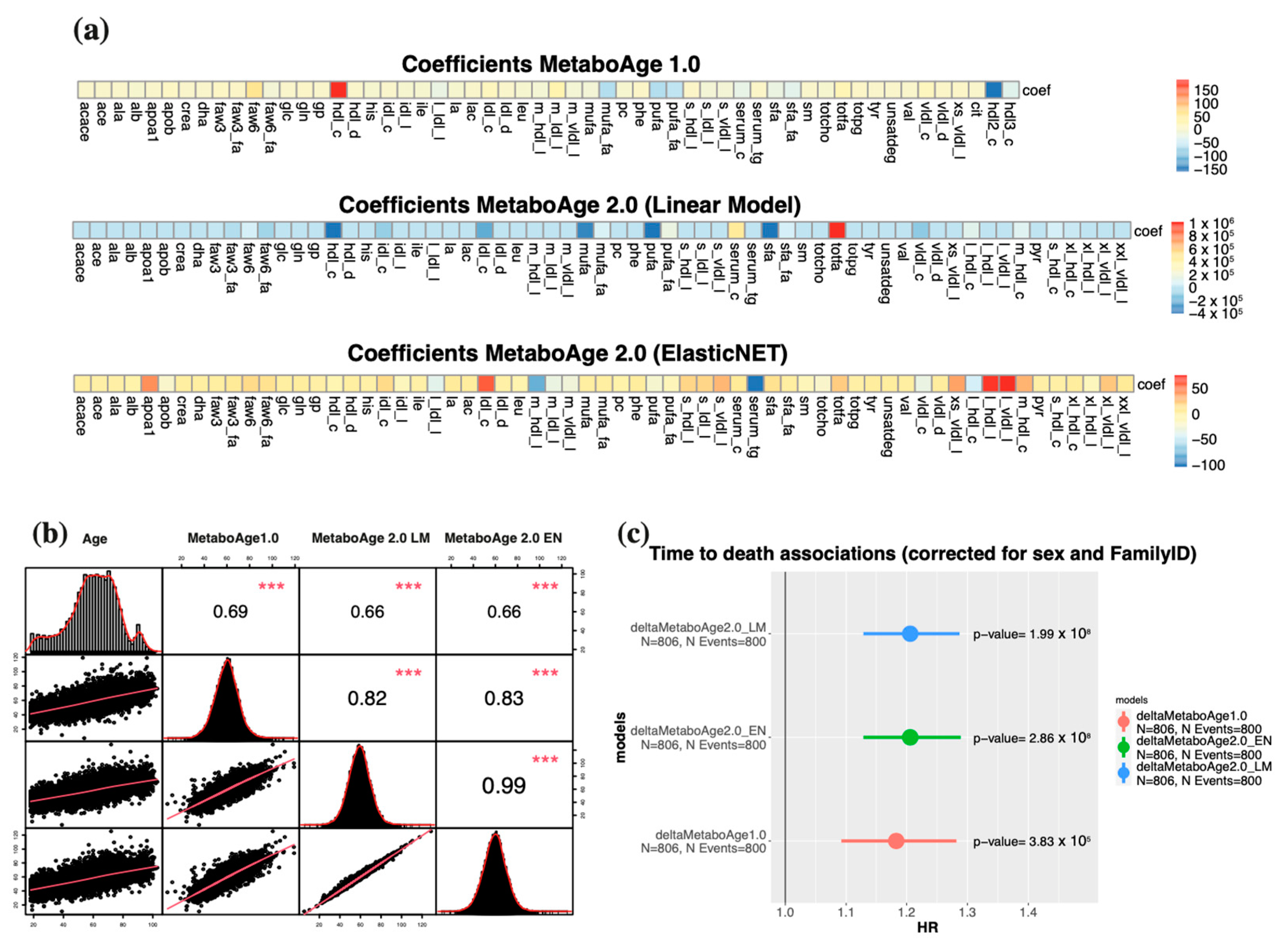
3.5. A Retrained MetaboAge on Re-Quantified Data Shows Similar Associations with Mortality Compared to the Previous Version of MetaboAge
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Van Den Akker, E.B.; Trompet, S.; Barkey Wolf, J.J.; Beekman, M.; Suchiman, H.E.; Deelen, J.; Asselbergs, F.W.; Boersma, E.; Cats, D.; Elders, P.M.; et al. Metabolic Age Based on the BBMRI-NL 1H-NMR Metabolomics Repository as Biomarker of Age-Related Disease. Circ. Genom. Precis. Med. 2020, 13, 541–547. [Google Scholar] [CrossRef]
- Deelen, J.; Kettunen, J.; Fischer, K.; van der Spek, A.; Trompet, S.; Kastenmüller, G.; Boyd, A.; Zierer, J.; van den Akker, E.B.; Ala-Korpela, M.; et al. A Metabolic Profile of All-Cause Mortality Risk Identified in an Observational Study of 44,168 Individuals. Nat. Commun. 2019, 10, 3346. [Google Scholar] [CrossRef] [PubMed]
- Nightingale Health UK Biobank Initiative; Julkunen, H.; Cichońska, A.; Slagboom, P.E.; Würtz, P. Metabolic Biomarker Profiling for Identification of Susceptibility to Severe Pneumonia and COVID-19 in the General Population. eLife 2021, 10, e63033. [Google Scholar] [CrossRef] [PubMed]
- Buergel, T.; Steinfeldt, J.; Ruyoga, G.; Pietzner, M.; Bizzarri, D.; Vojinovic, D.; Upmeier zu Belzen, J.; Loock, L.; Kittner, P.; Christmann, L.; et al. Metabolomic Profiles Predict Individual Multidisease Outcomes. Nat. Med. 2022, 28, 2309–2320. [Google Scholar] [CrossRef] [PubMed]
- Group, N.H.B.C.; Barrett, J.C.; Esko, T.; Fischer, K.; Jostins-Dean, L.; Jousilahti, P.; Julkunen, H.; Jääskeläinen, T.; Kerimov, N.; Kerminen, S.; et al. Metabolomic and Genomic Prediction of Common Diseases in 477,706 Participants in Three National Biobanks. medRxiv 2023, 2023, 2023-06. [Google Scholar]
- Xu, Y.; Ritchie, S.C.; Liang, Y.; Timmers, P.R.H.J.; Pietzner, M.; Lannelongue, L.; Lambert, S.A.; Tahir, U.A.; May-Wilson, S.; Foguet, C.; et al. An Atlas of Genetic Scores to Predict Multi-Omic Traits. Nature 2023, 616, 123–131. [Google Scholar] [CrossRef] [PubMed]
- Bizzarri, D.; Reinders, M.J.T.; Beekman, M.; Slagboom, P.E.; Bbmri-Nl; van den Akker, E.B. 1H-NMR Metabolomics-Based Surrogates to Impute Common Clinical Risk Factors and Endpoints. EBioMedicine 2022, 75, 103764. [Google Scholar] [CrossRef]
- Bharti, S.K.; Roy, R. Quantitative 1H NMR Spectroscopy. TrAC Trends Anal. Chem. 2012, 35, 5–26. [Google Scholar] [CrossRef]
- Dias, D.A.; Koal, T. Progress in Metabolomics Standardisation and Its Significance in Future Clinical Laboratory Medicine. EJIFCC 2016, 27, 331–343. [Google Scholar]
- Fearnley, L.G.; Inouye, M. Metabolomics in Epidemiology: From Metabolite Concentrations to Integrative Reaction Networks. Int. J. Epidemiol. 2016, 45, 1319–1328. [Google Scholar] [CrossRef]
- Yu, B.; Zanetti, K.A.; Temprosa, M.; Albanes, D.; Appel, N.; Barrera, C.B.; Ben-Shlomo, Y.; Boerwinkle, E.; Casas, J.P.; Clish, C.; et al. The Consortium of Metabolomics Studies (COMETS): Metabolomics in 47 Prospective Cohort Studies. Am. J. Epidemiol. 2019, 188, 991–1012. [Google Scholar] [CrossRef]
- Logue, M.W.; Smith, A.K.; Wolf, E.J.; Maniates, H.; Stone, A.; Schichman, S.A.; McGlinchey, R.E.; Milberg, W.; Miller, M.W. The Correlation of Methylation Levels Measured Using Illumina 450K and EPIC BeadChips in Blood Samples. Epigenomics 2017, 9, 1363–1371. [Google Scholar] [CrossRef]
- Würtz, P.; Kangas, A.J.; Soininen, P.; Lawlor, D.A.; Davey Smith, G.; Ala-Korpela, M. Quantitative Serum Nuclear Magnetic Resonance Metabolomics in Large-Scale Epidemiology: A Primer on -Omic Technologies. Am. J. Epidemiol. 2017, 186, 1084–1096. [Google Scholar] [CrossRef]
- Ala-Korpela, M.; Lehtimäki, T.; Kähönen, M.; Viikari, J.; Perola, M.; Salomaa, V.; Kettunen, J.; Raitakari, O.T.; Mäkinen, V.-P. Cross-Sectionally Calculated Metabolic Ageing Does Not Relate to Longitudinal Metabolic Changes—Support for Stratified Ageing Models. J. Clin. Endocrinol. Metab. 2023, 108, dgad032. [Google Scholar] [CrossRef]
- Mäkinen, V.-P.; Karsikas, M.; Kettunen, J.; Lehtimäki, T.; Kähönen, M.; Viikari, J.; Perola, M.; Salomaa, V.; Järvelin, M.-R.; Raitakari, O.T.; et al. Longitudinal Profiling of Metabolic Ageing Trends in Two Population Cohorts of Young Adults. Int. J. Epidemiol. 2022, 51, 1970–1983. [Google Scholar] [CrossRef]
- Geleijnse, J.M.; Giltay, E.J.; Schouten, E.G.; de Goede, J.; Oude Griep, L.M.; Teitsma-Jansen, A.M.; Katan, M.B.; Kromhout, D. Alpha Omega Trial Group Effect of Low Doses of N-3 Fatty Acids on Cardiovascular Diseases in 4,837 Post-Myocardial Infarction Patients: Design and Baseline Characteristics of the Alpha Omega Trial. Am. Heart J. 2010, 159, 539–546.e2. [Google Scholar] [CrossRef]
- Oemrawsingh, R.M.; Akkerhuis, K.M.; Umans, V.A.; Kietselaer, B.; Schotborgh, C.; Ronner, E.; Lenderink, T.; Liem, A.; Haitsma, D.; van der Harst, P.; et al. Cohort Profile of BIOMArCS: The BIOMarker Study to Identify the Acute Risk of a Coronary Syndrome-a Prospective Multicentre Biomarker Study Conducted in the Netherlands. BMJ Open 2016, 6, e012929. [Google Scholar] [CrossRef]
- van Oosterhout, W.P.J.; Weller, C.M.; Stam, A.H.; Bakels, F.; Stijnen, T.; Ferrari, M.D.; Terwindt, G.M. Validation of the Web-Based LUMINA Questionnaire for Recruiting Large Cohorts of Migraineurs. Cephalalgia 2011, 31, 1359–1367. [Google Scholar] [CrossRef]
- Wesseling, J.; Dekker, J.; van den Berg, W.B.; Bierma-Zeinstra, S.M.A.; Boers, M.; Cats, H.A.; Deckers, P.; Gorter, K.J.; Heuts, P.H.T.G.; Hilberdink, W.K.H.A.; et al. CHECK (Cohort Hip and Cohort Knee): Similarities and Differences with the Osteoarthritis Initiative. Ann. Rheum. Dis. 2009, 68, 1413–1419. [Google Scholar] [CrossRef]
- van Greevenbroek, M.M.J.; Jacobs, M.; van der Kallen, C.J.H.; Vermeulen, V.M.M.-J.; Jansen, E.H.J.M.; Schalkwijk, C.G.; Ferreira, I.; Feskens, E.J.M.; Stehouwer, C.D.A. The Cross-Sectional Association between Insulin Resistance and Circulating Complement C3 Is Partly Explained by Plasma Alanine Aminotransferase, Independent of Central Obesity and General Inflammation (the CODAM Study). Eur. J. Clin. Investig. 2011, 41, 372–379. [Google Scholar] [CrossRef]
- Schram, M.T.; Sep, S.J.S.; van der Kallen, C.J.; Dagnelie, P.C.; Koster, A.; Schaper, N.; Henry, R.M.A.; Stehouwer, C.D.A. The Maastricht Study: An Extensive Phenotyping Study on Determinants of Type 2 Diabetes, Its Complications and Its Comorbidities. Eur. J. Epidemiol. 2014, 29, 439–451. [Google Scholar] [CrossRef]
- van der Heijden, A.A.; Rauh, S.P.; Dekker, J.M.; Beulens, J.W.; Elders, P.; ’t Hart, L.M.; Rutters, F.; van Leeuwen, N.; Nijpels, G. The Hoorn Diabetes Care System (DCS) Cohort. A Prospective Cohort of Persons with Type 2 Diabetes Treated in Primary Care in the Netherlands. BMJ Open 2017, 7, e015599. [Google Scholar] [CrossRef]
- Pardo, L.M.; MacKay, I.; Oostra, B.; van Duijn, C.M.; Aulchenko, Y.S. The Effect of Genetic Drift in a Young Genetically Isolated Population. Ann. Hum. Genet. 2005, 69, 288–295. [Google Scholar] [CrossRef]
- Schirmer, M.; Smeekens, S.P.; Vlamakis, H.; Jaeger, M.; Oosting, M.; Franzosa, E.A.; Ter Horst, R.; Jansen, T.; Jacobs, L.; Bonder, M.J.; et al. Linking the Human Gut Microbiome to Inflammatory Cytokine Production Capacity. Cell 2016, 167, 1125–1136.e8. [Google Scholar] [CrossRef]
- Riyazi, N.; Meulenbelt, I.; Kroon, H.M.; Ronday, K.H.; Hellio le Graverand, M.-P.; Rosendaal, F.R.; Breedveld, F.C.; Slagboom, P.E.; Kloppenburg, M. Evidence for Familial Aggregation of Hand, Hip, and Spine but Not Knee Osteoarthritis in Siblings with Multiple Joint Involvement: The GARP Study. Ann. Rheum. Dis. 2005, 64, 438–443. [Google Scholar] [CrossRef]
- Stronks, K.; Snijder, M.B.; Peters, R.J.G.; Prins, M.; Schene, A.H.; Zwinderman, A.H. Unravelling the Impact of Ethnicity on Health in Europe: The HELIUS Study. BMC Public Health 2013, 13, 402. [Google Scholar] [CrossRef]
- Lumey, L.H.; Stein, A.D.; Kahn, H.S.; van der Pal-de Bruin, K.M.; Blauw, G.J.; Zybert, P.A.; Susser, E.S. Cohort Profile: The Dutch Hunger Winter Families Study. Int. J. Epidemiol. 2007, 36, 1196–1204. [Google Scholar] [CrossRef]
- Scholtens, S.; Smidt, N.; Swertz, M.A.; Bakker, S.J.L.; Dotinga, A.; Vonk, J.M.; van Dijk, F.; van Zon, S.K.R.; Wijmenga, C.; Wolffenbuttel, B.H.R.; et al. Cohort Profile: LifeLines, a Three-Generation Cohort Study and Biobank. Int. J. Epidemiol. 2015, 44, 1172–1180. [Google Scholar] [CrossRef]
- Schoenmaker, M.; de Craen, A.J.M.; de Meijer, P.H.E.M.; Beekman, M.; Blauw, G.J.; Slagboom, P.E.; Westendorp, R.G.J. Evidence of Genetic Enrichment for Exceptional Survival Using a Family Approach: The Leiden Longevity Study. Eur. J. Hum. Genet. 2006, 14, 79–84. [Google Scholar] [CrossRef]
- Penninx, B.W.J.H.; Beekman, A.T.F.; Smit, J.H.; Zitman, F.G.; Nolen, W.A.; Spinhoven, P.; Cuijpers, P.; De Jong, P.J.; Van Marwijk, H.W.J.; Assendelft, W.J.J.; et al. The Netherlands Study of Depression and Anxiety (NESDA): Rationale, Objectives and Methods. Int. J. Methods Psychiatr. Res. 2008, 17, 121–140. [Google Scholar] [CrossRef]
- Shepherd, J.; Blauw, G.J.; Murphy, M.B.; Cobbe, S.M.; Bollen, E.L.; Buckley, B.M.; Ford, I.; Jukema, J.W.; Hyland, M.; Gaw, A.; et al. The Design of a Prospective Study of Pravastatin in the Elderly at Risk (PROSPER). PROSPER Study Group. PROspective Study of Pravastatin in the Elderly at Risk. Am. J. Cardiol. 1999, 84, 1192–1197. [Google Scholar] [CrossRef]
- Ramos, Y.F.M.; den Hollander, W.; Bovée, J.V.M.G.; Bomer, N.; van der Breggen, R.; Lakenberg, N.; Keurentjes, J.C.; Goeman, J.J.; Slagboom, P.E.; Nelissen, R.G.H.H.; et al. Genes Involved in the Osteoarthritis Process Identified through Genome Wide Expression Analysis in Articular Cartilage; the RAAK Study. PLoS ONE 2014, 9, e103056. [Google Scholar] [CrossRef]
- So-Osman, C.; Nelissen, R.G.H.H.; Koopman-van Gemert, A.W.M.M.; Kluyver, E.; Pöll, R.G.; Onstenk, R.; Van Hilten, J.A.; Jansen-Werkhoven, T.M.; van den Hout, W.B.; Brand, R.; et al. Patient Blood Management in Elective Total Hip- and Knee-Replacement Surgery (Part 1): A Randomized Controlled Trial on Erythropoietin and Blood Salvage as Transfusion Alternatives Using a Restrictive Transfusion Policy in Erythropoietin-Eligible Patients. Anesthesiology 2014, 120, 839–851. [Google Scholar] [CrossRef] [PubMed]
- Ikram, M.A.; Brusselle, G.G.O.; Murad, S.D.; van Duijn, C.M.; Franco, O.H.; Goedegebure, A.; Klaver, C.C.W.; Nijsten, T.E.C.; Peeters, R.P.; Stricker, B.H.; et al. The Rotterdam Study: 2018 Update on Objectives, Design and Main Results. Eur. J. Epidemiol. 2017, 32, 807–850. [Google Scholar] [CrossRef] [PubMed]
- van der Flier, W.M.; Pijnenburg, Y.A.L.; Prins, N.; Lemstra, A.W.; Bouwman, F.H.; Teunissen, C.E.; van Berckel, B.N.M.; Stam, C.J.; Barkhof, F.; Visser, P.J.; et al. Optimizing Patient Care and Research: The Amsterdam Dementia Cohort. J. Alzheimers Dis. 2014, 41, 313–327. [Google Scholar] [CrossRef] [PubMed]
- Eppinga, R.N.; Kofink, D.; Dullaart, R.P.F.; Dalmeijer, G.W.; Lipsic, E.; van Veldhuisen, D.J.; van der Horst, I.C.C.; Asselbergs, F.W.; van der Harst, P. Effect of Metformin on Metabolites and Relation with Myocardial Infarct Size and Left Ventricular Ejection Fraction After Myocardial Infarction. Circ. Cardiovasc. Genet. 2017, 10, e001564. [Google Scholar] [CrossRef]
- Gijsberts, C.M.; Agostoni, P.; Hoefer, I.E.; Asselbergs, F.W.; Pasterkamp, G.; Nathoe, H.; Appelman, Y.E.; de Kleijn, D.P.V.; den Ruijter, H.M. Gender Differences in Health-Related Quality of Life in Patients Undergoing Coronary Angiography. Open Heart 2015, 2, e000231. [Google Scholar] [CrossRef]
- van Beijsterveldt, C.E.M.; Groen-Blokhuis, M.; Hottenga, J.J.; Franić, S.; Hudziak, J.J.; Lamb, D.; Huppertz, C.; de Zeeuw, E.; Nivard, M.; Schutte, N.; et al. The Young Netherlands Twin Register (YNTR): Longitudinal Twin and Family Studies in over 70,000 Children. Twin Res. Hum. Genet. 2013, 16, 252–267. [Google Scholar] [CrossRef]
- Bizzarri, D.; Reinders, M.J.T.; Beekman, M.; Slagboom, P.E.; van den Akker, E.B. MiMIR: R-Shiny Application to Infer Risk Factors and Endpoints from Nightingale Health’s 1H-NMR Metabolomics Data. Bioinformatics 2022, 38, 3847–3849. [Google Scholar] [CrossRef]
- Horvath, S. DNA Methylation Age of Human Tissues and Cell Types. Genome Biol. 2013, 14, R115. [Google Scholar] [CrossRef]
- Lu, A.T.; Quach, A.; Wilson, J.G.; Reiner, A.P.; Aviv, A.; Raj, K.; Hou, L.; Baccarelli, A.A.; Li, Y.; Stewart, J.D.; et al. DNA Methylation GrimAge Strongly Predicts Lifespan and Healthspan. Aging 2019, 11, 303–327. [Google Scholar] [CrossRef] [PubMed]
- Julkunen, H.; Cichońska, A.; Tiainen, M.; Koskela, H.; Nybo, K.; Mäkelä, V.; Nokso-Koivisto, J.; Kristiansson, K.; Perola, M.; Salomaa, V.; et al. Atlas of Plasma NMR Biomarkers for Health and Disease in 118,461 Individuals from the UK Biobank. Nat. Commun. 2023, 14, 604. [Google Scholar] [CrossRef] [PubMed]
- Lever, J.; Krzywinski, M.; Altman, N. Regularization. Nat. Methods 2016, 13, 803–804. [Google Scholar] [CrossRef]
- Peng, R.D.; Dominici, F.; Zeger, S.L. Reproducible Epidemiologic Research. Am. J. Epidemiol. 2006, 163, 783–789. [Google Scholar] [CrossRef]
- Schober, P.; Vetter, T.R. Repeated Measures Designs and Analysis of Longitudinal Data: If at First You Do Not Succeed—Try, Try Again. Anesth. Analg. 2018, 127, 569–575. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Waves | N. Samples | N. Biobanks | Platform Version |
|---|---|---|---|
| First wave | 24,994 | 26 | Version 2014 |
| Second wave | 9880 | 10 | Version 2016 |
| Re-quantifications | 34,015 | 28 | Version 2020 |
| LLS-PAROFFS [30–79 years old] | ||||||
| Wave | Platform version first measure | Re-quantification | Total N. samples | N. samples after QC | Drop rate (%) | |
| IOP1 | First Wave | Version 2014 | Version 2020 | 2313 | 1925 | 16.77 |
| IOP2 | Second Wave | Version 2016 | Version 2020 | 670 | 604 | 9.85 |
| IOP3 | Third Wave | Version 2016 | Version 2020 | 498 | 400 | 19.68 |
| LLS-SIBS [89–103 years old] | ||||||
| IOP1 | First Wave | Version 2014 | Version 2020 | 998 | 948 | 5.01 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bizzarri, D.; Reinders, M.J.T.; Beekman, M.; Slagboom, P.E.; van den Akker, E.B.; on behalf of the BbmriNl. Technical Report: A Comprehensive Comparison between Different Quantification Versions of Nightingale Health’s 1H-NMR Metabolomics Platform. Metabolites 2023, 13, 1181. https://doi.org/10.3390/metabo13121181
Bizzarri D, Reinders MJT, Beekman M, Slagboom PE, van den Akker EB, on behalf of the BbmriNl. Technical Report: A Comprehensive Comparison between Different Quantification Versions of Nightingale Health’s 1H-NMR Metabolomics Platform. Metabolites. 2023; 13(12):1181. https://doi.org/10.3390/metabo13121181
Chicago/Turabian StyleBizzarri, Daniele, Marcel J. T. Reinders, Marian Beekman, P. Eline Slagboom, Erik B. van den Akker, and on behalf of the BbmriNl. 2023. "Technical Report: A Comprehensive Comparison between Different Quantification Versions of Nightingale Health’s 1H-NMR Metabolomics Platform" Metabolites 13, no. 12: 1181. https://doi.org/10.3390/metabo13121181
APA StyleBizzarri, D., Reinders, M. J. T., Beekman, M., Slagboom, P. E., van den Akker, E. B., & on behalf of the BbmriNl. (2023). Technical Report: A Comprehensive Comparison between Different Quantification Versions of Nightingale Health’s 1H-NMR Metabolomics Platform. Metabolites, 13(12), 1181. https://doi.org/10.3390/metabo13121181