
Targeting Longevity Gene SLC13A5: A Novel Approach to Prevent Age-Related Bone Fragility and Osteoporosis

 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Mendelian Randomization
2.2. Causal Biomarker Analysis
2.3. Animals
2.4. Serum Analysis
2.5. High-Resolution Micro-Computed Tomographic Imaging (Micro-CT)
2.6. Whole Bone Biomechanical Testing
3. Results
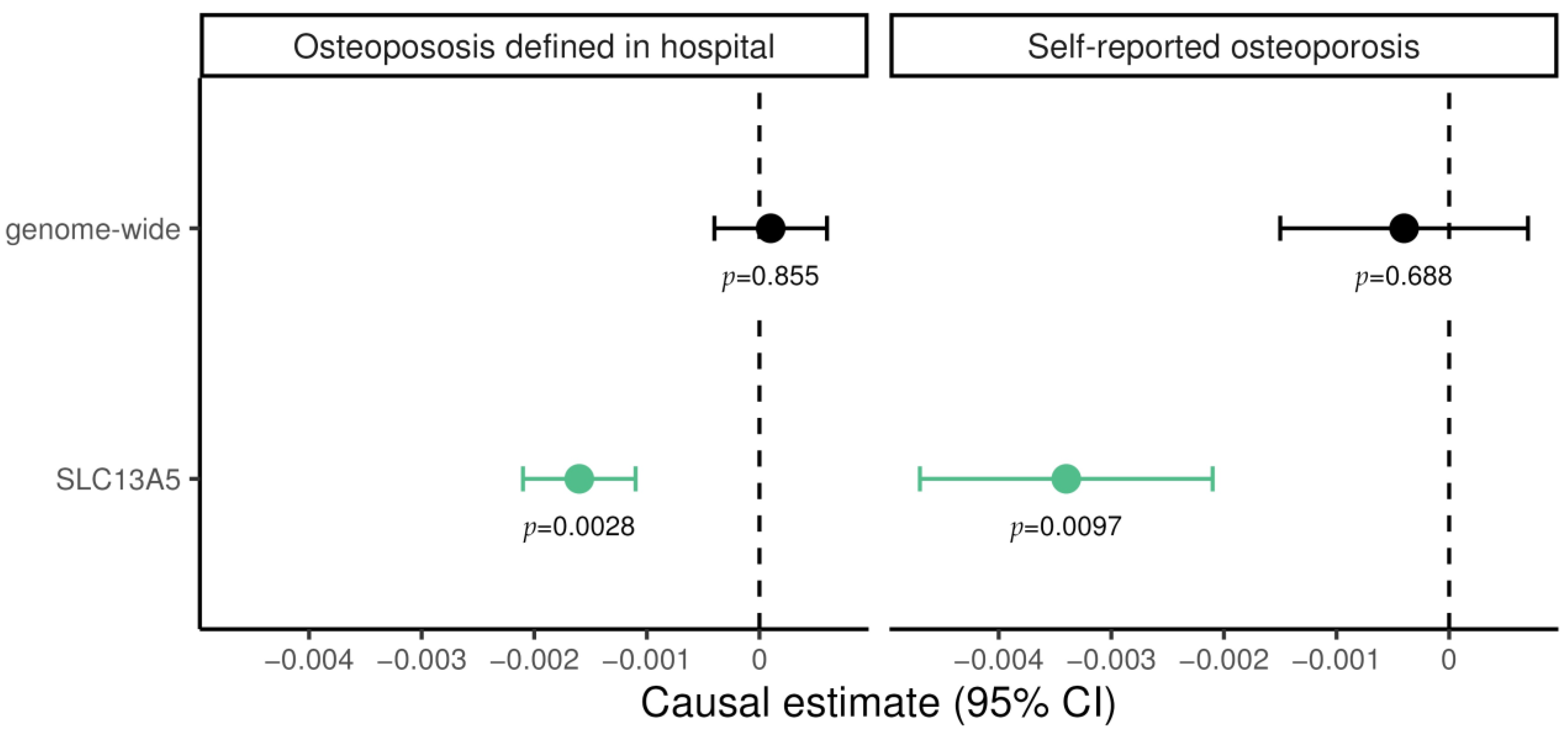
3.1. Mendelian Randomization
3.2. Causal Biomarker Analysis
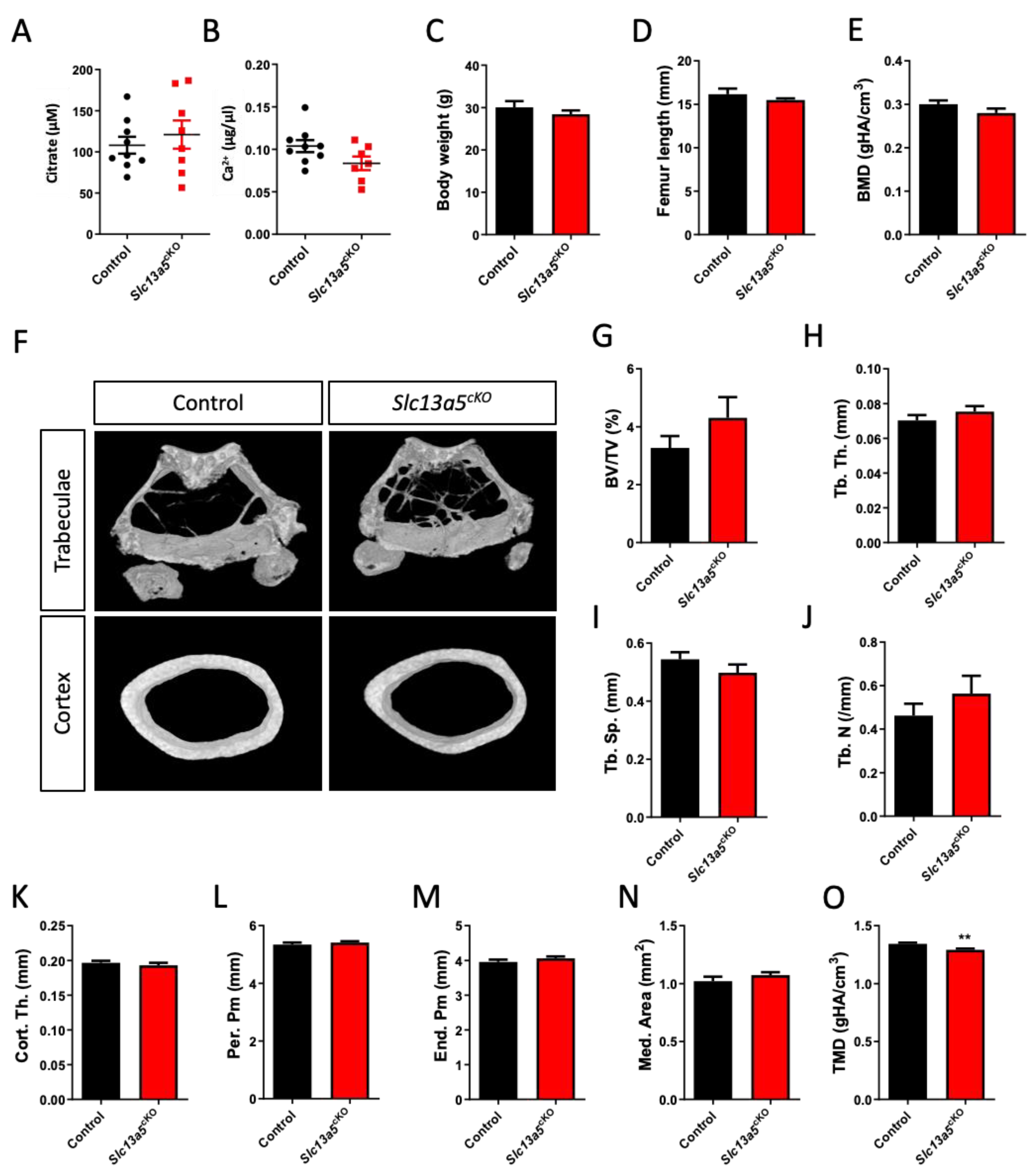
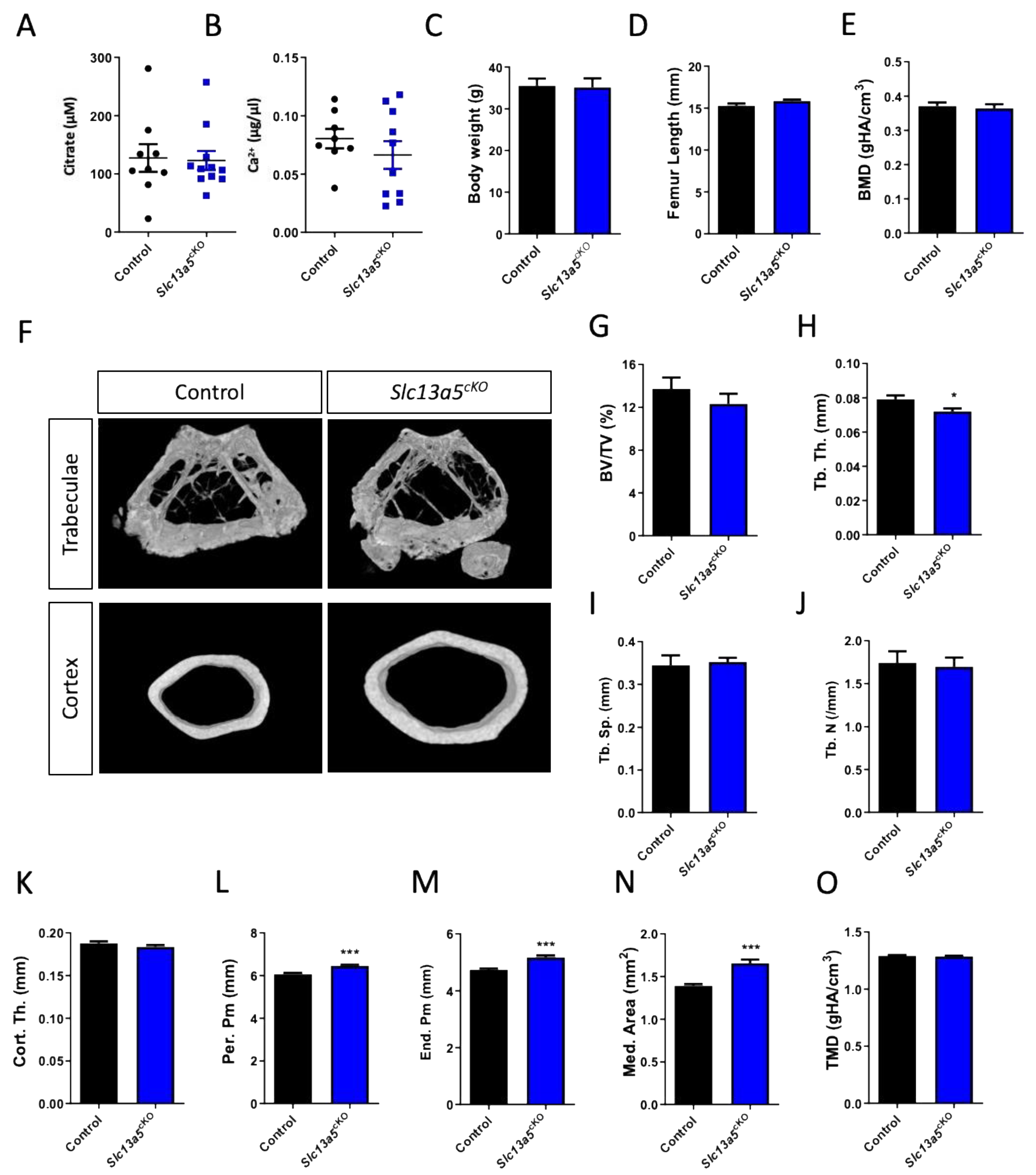
3.3. Age-Induced Changes in Bone Morphology and Mechanical Properties in Skeletal-Specific Slc13a5cKO Female and Male Mice
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Rogina, B.; Reenan, R.A.; Nilsen, S.P.; Helfand, S.L. Extended Life-Span Conferred by Cotransporter Gene Mutations in Drosophila. Science 2000, 290, 2137–2140. [Google Scholar] [CrossRef]
- Inoue, K.; Zhuang, L.; Maddox, D.M.; Smith, S.B.; Ganapathy, V. Structure, Function, and Expression Pattern of a Novel Sodium-Coupled Citrate Transporter (NaCT) Cloned from Mammalian Brain. J. Biol. Chem. 2002, 277, 39469–39476. [Google Scholar] [CrossRef]
- Birkenfeld, A.L.; Lee, H.-Y.; Guebre-Egziabher, F.; Alves, T.C.; Jurczak, M.J.; Jornayvaz, F.R.; Zhang, D.; Hsiao, J.J.; Martin-Montalvo, A.; Fischer-Rosinsky, A.; et al. Deletion of the Mammalian INDY Homolog Mimics Aspects of Dietary Restriction and Protects against Adiposity and Insulin Resistance in Mice. Cell Metab. 2011, 14, 184–195. [Google Scholar] [CrossRef] [PubMed]
- Willmes, D.M.; Daniels, M.; Kurzbach, A.; Lieske, S.; Bechmann, N.; Schumann, T.; Henke, C.; El-Agroudy, N.N.; Da Costa Goncalves, A.C.; Peitzsch, M.; et al. The Longevity Gene mIndy (I’m Not Dead, Yet) Affects Blood Pressure through Sympathoadrenal Mechanisms. JCI Insight 2021, 6, e136083. [Google Scholar] [CrossRef] [PubMed]
- Huard, K.; Brown, J.; Jones, J.C.; Cabral, S.; Futatsugi, K.; Gorgoglione, M.; Lanba, A.; Vera, N.B.; Zhu, Y.; Yan, Q.; et al. Discovery and Characterization of Novel Inhibitors of the Sodium-Coupled Citrate Transporter (NaCT or SLC13A5). Sci. Rep. 2015, 5, 17391. [Google Scholar] [CrossRef] [PubMed]
- Pesta, D.; Jordan, J. INDY as a Therapeutic Target for Cardio-Metabolic Disease. Metabolites 2022, 12, 244. [Google Scholar] [CrossRef]
- Zahn, G.; Willmes, D.M.; El-Agroudy, N.N.; Yarnold, C.; Jarjes-Pike, R.; Schaertl, S.; Schreiter, K.; Gehrmann, W.; Wong, A.K.C.; Zordan, T.; et al. A Novel and Cross-Species Active Mammalian INDY (NaCT) Inhibitor Ameliorates Hepatic Steatosis in Mice with Diet-Induced Obesity. Metabolites 2022, 12, 732. [Google Scholar] [CrossRef] [PubMed]
- Irizarry, A.R.; Yan, G.; Zeng, Q.; Lucchesi, J.; Hamang, M.J.; Ma, Y.L.; Rong, J.X. Defective Enamel and Bone Development in Sodium-Dependent Citrate Transporter (NaCT) Slc13a5 Deficient Mice. PLoS ONE 2017, 12, e0175465. [Google Scholar] [CrossRef] [PubMed]
- Dirckx, N.; Zhang, Q.; Chu, E.Y.; Tower, R.J.; Li, Z.; Guo, S.; Yuan, S.; Khare, P.A.; Zhang, C.; Verardo, A.; et al. A Specialized Metabolic Pathway Partitions Citrate in Hydroxyapatite to Impact Mineralization of Bones and Teeth. Proc. Natl. Acad. Sci. USA 2022, 119, e2212178119. [Google Scholar] [CrossRef]
- Thevenon, J.; Milh, M.; Feillet, F.; St-Onge, J.; Duffourd, Y.; Jugé, C.; Roubertie, A.; Héron, D.; Mignot, C.; Raffo, E.; et al. Mutations in SLC13A5 Cause Autosomal-Recessive Epileptic Encephalopathy with Seizure Onset in the First Days of Life. Am. J. Hum. Genet. 2014, 95, 113–120. [Google Scholar] [CrossRef]
- Hardies, K.; de Kovel, C.G.F.; Weckhuysen, S.; Asselbergh, B.; Geuens, T.; Deconinck, T.; Azmi, A.; May, P.; Brilstra, E.; Becker, F.; et al. Recessive Mutations in SLC13A5 Result in a Loss of Citrate Transport and Cause Neonatal Epilepsy, Developmental Delay and Teeth Hypoplasia. Brain 2015, 138, 3238–3250. [Google Scholar] [CrossRef]
- von Loeffelholz, C.; Lieske, S.; Neuschäfer-Rube, F.; Willmes, D.M.; Raschzok, N.; Sauer, I.M.; König, J.; Fromm, M.F.; Horn, P.; Chatzigeorgiou, A.; et al. The Human Longevity Gene Homolog INDY and Interleukin-6 Interact in Hepatic Lipid Metabolism. Hepatology 2017, 66, 616–630. [Google Scholar] [CrossRef]
- Kopel, J.J.; Bhutia, Y.D.; Sivaprakasam, S.; Ganapathy, V. Consequences of NaCT/SLC13A5/mINDY Deficiency: Good versus Evil, Separated Only by the Blood–brain Barrier. Biochem. J. 2021, 478, 463–486. [Google Scholar] [CrossRef]
- Gopal, E.; Babu, E.; Ramachandran, S.; Bhutia, Y.D.; Prasad, P.D.; Ganapathy, V. Species-Specific Influence of Lithium on the Activity of SLC13A5 (NaCT): Lithium-Induced Activation Is Specific for the Transporter in Primates. J. Pharmacol. Exp. Ther. 2015, 353, 17–26. [Google Scholar] [CrossRef]
- Dickens, F. The Citric Acid Content of Animal Tissues, with Reference to Its Occurrence in Bone and Tumour. Biochem. J. 1941, 35, 1011–1023. [Google Scholar] [CrossRef]
- Costello, L.C.; Franklin, R.B.; Reynolds, M.A.; Chellaiah, M. The Important Role of Osteoblasts and Citrate Production in Bone Formation: “Osteoblast Citration” as a New Concept for an Old Relationship. Open Bone J. 2012, 4, 27–34. [Google Scholar] [CrossRef]
- Ma, C.; Tian, X.; Kim, J.P.; Xie, D.; Ao, X.; Shan, D.; Lin, Q.; Hudock, M.R.; Bai, X.; Yang, J. Citrate-Based Materials Fuel Human Stem Cells by Metabonegenic Regulation. Proc. Natl. Acad. Sci. USA 2018, 115, E11741–E11750. [Google Scholar] [CrossRef]
- Granchi, D.; Baldini, N.; Ulivieri, F.M.; Caudarella, R. Role of Citrate in Pathophysiology and Medical Management of Bone Diseases. Nutrients 2019, 11, 2576. [Google Scholar] [CrossRef]
- Katan, M. Apoupoprotein e isoforms, serum cholesterol, and cancer. Lancet 1986, 327, 507–508. [Google Scholar] [CrossRef]
- Baukmann, H.A.; Cope, J.L.; Ravarani, C.N.J.; Bannard, C.; Lamparter, M.R.J.; Schwinges, A.R.E.C.; Klinger, J.E.; Schmidt, M.F. Neutrophils Cause Critical Illness in COVID-19 and Reveal CDK6 Inhibitors as Potential Treatment. medRxiv 2021. [Google Scholar] [CrossRef]
- Burgess, S.; Mason, A.M.; Grant, A.J.; Slob, E.A.W.; Gkatzionis, A.; Zuber, V.; Patel, A.; Tian, H.; Liu, C.; Haynes, W.G.; et al. Using Genetic Association Data to Guide Drug Discovery and Development: Review of Methods and Applications. Am. J. Hum. Genet. 2023, 110, 195–214. [Google Scholar] [CrossRef]
- Gill, D.; Georgakis, M.K.; Walker, V.M.; Schmidt, A.F.; Gkatzionis, A.; Freitag, D.F.; Finan, C.; Hingorani, A.D.; Howson, J.M.M.; Burgess, S.; et al. Mendelian Randomization for Studying the Effects of Perturbing Drug Targets. Wellcome Open Res. 2021, 6, 16. [Google Scholar] [CrossRef]
- Gill, D.; Zagkos, L.; Gill, R.; Benzing, T.; Jordan, J.; Birkenfeld, A.L.; Burgess, S.; Zahn, G. The Citrate Transporter SLC13A5 as a Therapeutic Target for Kidney Disease: Evidence from Mendelian Randomization to Inform Drug Development. medRxiv 2023. [Google Scholar] [CrossRef]
- Purcell, S.; Neale, B.; Todd-Brown, K.; Thomas, L.; Ferreira, M.A.R.; Bender, D.; Maller, J.; Sklar, P.; de Bakker, P.I.W.; Daly, M.J.; et al. PLINK: A Tool Set for Whole-Genome Association and Population-Based Linkage Analyses. Am. J. Hum. Genet. 2007, 81, 559–575. [Google Scholar] [CrossRef]
- Bowden, J.; Del Greco, M.F.; Minelli, C.; Davey Smith, G.; Sheehan, N.; Thompson, J. A Framework for the Investigation of Pleiotropy in Two-Sample Summary Data Mendelian Randomization. Stat. Med. 2017, 36, 1783–1802. [Google Scholar] [CrossRef]
- Hemani, G.; Zheng, J.; Elsworth, B.; Wade, K.H.; Haberland, V.; Baird, D.; Laurin, C.; Burgess, S.; Bowden, J.; Langdon, R.; et al. The MR-Base Platform Supports Systematic Causal Inference across the Human Phenome. Elife 2018, 7, e34408. [Google Scholar] [CrossRef]
- Bycroft, C.; Freeman, C.; Petkova, D.; Band, G.; Elliott, L.T.; Sharp, K.; Motyer, A.; Vukcevic, D.; Delaneau, O.; O’Connell, J.; et al. The UK Biobank Resource with Deep Phenotyping and Genomic Data. Nature 2018, 562, 203–209. [Google Scholar] [CrossRef]
- Thrailkill, K.M.; Nyman, J.S.; Bunn, R.C.; Uppuganti, S.; Thompson, K.L.; Lumpkin, C.K., Jr.; Kalaitzoglou, E.; Fowlkes, J.L. The Impact of SGLT2 Inhibitors, Compared with Insulin, on Diabetic Bone Disease in a Mouse Model of Type 1 Diabetes. Bone 2017, 94, 141–151. [Google Scholar] [CrossRef]
- Imai, K.; Van Dyk, D.A. Causal Inference with General Treatment Regimes: Generalizing the Propensity Score. J. Am. Stat. Assoc. 2004, 29, 854–886. [Google Scholar] [CrossRef]
- Kar, S.P.; Brenner, H.; Giles, G.G.; Huo, D.; Milne, R.L.; Rennert, G.; Simard, J.; Zheng, W.; Burgess, S.; Pharoah, P.D.P. Body Mass Index and the Association between Low-Density Lipoprotein Cholesterol as Predicted by HMGCR Genetic Variants and Breast Cancer Risk. Int. J. Epidemiol. 2019, 48, 1727–1730. [Google Scholar] [CrossRef]
- Orho-Melander, M.; Hindy, G.; Borgquist, S.; Schulz, C.-A.; Manjer, J.; Melander, O.; Stocks, T. Blood Lipid Genetic Scores, the HMGCR Gene and Cancer Risk: A Mendelian Randomization Study. Int. J. Epidemiol. 2018, 47, 495–505. [Google Scholar] [CrossRef]
- Nethander, M.; Coward, E.; Reimann, E.; Grahnemo, L.; Gabrielsen, M.E.; Wibom, C.; Estonian Biobank Research Team; Mägi, R.; Funck-Brentano, T.; Hoff, M.; et al. Assessment of the Genetic and Clinical Determinants of Hip Fracture Risk: Genome-Wide Association and Mendelian Randomization Study. Cell Rep. Med. 2022, 3, 100776. [Google Scholar] [CrossRef]
- Trajanoska, K.; Morris, J.A.; Oei, L.; Zheng, H.-F.; Evans, D.M.; Kiel, D.P.; Ohlsson, C.; Richards, J.B.; Rivadeneira, F.; GEFOS/GENOMOS Consortium and the 23andMe Research Team. Assessment of the Genetic and Clinical Determinants of Fracture Risk: Genome Wide Association and Mendelian Randomisation Study. BMJ 2018, 362, k3225. [Google Scholar] [CrossRef]
- Tang, Y.; Wei, F.; Yu, M.; Zhou, H.; Wang, Y.; Cui, Z.; Liu, X. Absence of Causal Association between Vitamin D and Bone Mineral Density across the Lifespan: A Mendelian Randomization Study. Sci. Rep. 2022, 12, 10408. [Google Scholar] [CrossRef]
- Zhu, H.; Li, Z.; Zhou, Y.; Zheng, R.; Diao, C.; Li, K.; Feng, Q. Neutrophil-Lymphocyte Ratio as a Risk Factor for Osteoporotic Vertebrae Fractures and Femoral Neck Fractures. Medicine 2022, 101, e32125. [Google Scholar] [CrossRef]
- Yilmaz, H.; Uyfun, M.; Yilmaz, T.S.; Namuslu, M.; Inan, O.; Taskin, A.; Cakmak, M.; Bilgic, M.A.; Bavbek, N.; Akcay, A.; et al. Neutrophil-Lymphocyte Ratio May Be Superior to C-Reactive Protein for Predicting the Occurrence of Postmenopausal Osteoporosis. Endocr. Regul. 2014, 48, 25–33. [Google Scholar] [CrossRef]
- Yu, X.-Y.; Li, X.-S.; Li, Y.; Liu, T.; Wang, R.-T. Neutrophil-Lymphocyte Ratio Is Associated with Arterial Stiffness in Postmenopausal Women with Osteoporosis. Arch. Gerontol. Geriatr. 2015, 61, 76–80. [Google Scholar] [CrossRef]
- Bainbridge, M.N.; Cooney, E.; Miller, M.; Kennedy, A.D.; Wulff, J.E.; Donti, T.; Jhangiani, S.N.; Gibbs, R.A.; Elsea, S.H.; Porter, B.E.; et al. Analyses of SLC13A5-Epilepsy Patients Reveal Perturbations of TCA Cycle. Mol. Genet. Metab. 2017, 121, 314–319. [Google Scholar] [CrossRef]
- Henke, C.; Töllner, K.; van Dijk, R.M.; Miljanovic, N.; Cordes, T.; Twele, F.; Bröer, S.; Ziesak, V.; Rohde, M.; Hauck, S.M.; et al. Disruption of the Sodium-Dependent Citrate Transporter SLC13A5 in Mice Causes Alterations in Brain Citrate Levels and Neuronal Network Excitability in the Hippocampus. Neurobiol. Dis. 2020, 143, 105018. [Google Scholar] [CrossRef]
- Li, Z.; Erion, D.M.; Maurer, T.S. Model-Based Assessment of Plasma Citrate Flux into the Liver: Implications for NaCT as a Therapeutic Target. CPT Pharmacomet. Syst. Pharmacol. 2016, 5, 132–139. [Google Scholar] [CrossRef]
- Costello, L.C.; Franklin, R.B. The Implications of the Hypocitricemic Response to Surgery and the Role of Liver Function and Hepatocyte Metabolism: An Important, but Neglected, Clinical Relationship. J. Liver Res. Disord. Ther. 2018, 4, 114–119. [Google Scholar] [CrossRef]
- Ho, H.T.B.; Ko, B.C.B.; Cheung, A.K.H.; Lam, A.K.M.; Tam, S.; Chung, S.K.; Chung, S.S.M. Generation and Characterization of Sodium-Dicarboxylate Cotransporter-Deficient Mice. Kidney Int. 2007, 72, 63–71. [Google Scholar] [CrossRef]
- Dewulf, J.P.; Wiame, E.; Dorboz, I.; Elmaleh-Bergès, M.; Imbard, A.; Dumitriu, D.; Rak, M.; Bourillon, A.; Helaers, R.; Malla, A.; et al. SLC13A3 Variants Cause Acute Reversible Leukoencephalopathy and α-Ketoglutarate Accumulation. Ann. Neurol. 2019, 85, 385–395. [Google Scholar] [CrossRef]
- Wang, Q.; Dhindsa, R.S.; Carss, K.; Harper, A.R.; Nag, A.; Tachmazidou, I.; Vitsios, D.; Deevi, S.V.V.; Mackay, A.; Muthas, D.; et al. Rare Variant Contribution to Human Disease in 281,104 UK Biobank Exomes. Nature 2021, 597, 527–532. [Google Scholar] [CrossRef]
- Moffatt, P.; Boraschi-Diaz, I.; Marulanda, J.; Bardai, G.; Rauch, F. Calvaria Bone Transcriptome in Mouse Models of Osteogenesis Imperfecta. Int. J. Mol. Sci. 2021, 22, 5290. [Google Scholar] [CrossRef]
- Fernandez-Fuente, G.; Overmyer, K.A.; Lawton, A.J.; Kasza, I.; Shapiro, S.L.; Gallego-Muñoz, P.; Coon, J.J.; Denu, J.M.; Alexander, C.M.; Puglielli, L. The Citrate Transporters SLC13A5 and SLC25A1 Elicit Different Metabolic Responses and Phenotypes in the Mouse. Commun. Biol. 2023, 6, 926. [Google Scholar] [CrossRef]
- Manolagas, S.C.; O’Brien, C.A.; Almeida, M. The Role of Estrogen and Androgen Receptors in Bone Health and Disease. Nat. Rev. Endocrinol. 2013, 9, 699–712. [Google Scholar] [CrossRef]
- Chen, H.; Wang, Y.; Dai, H.; Tian, X.; Cui, Z.-K.; Chen, Z.; Hu, L.; Song, Q.; Liu, A.; Zhang, Z.; et al. Bone and Plasma Citrate Is Reduced in Osteoporosis. Bone 2018, 114, 189–197. [Google Scholar] [CrossRef]
- Brown, T.L.; Nye, K.L.; Porter, B.E. Growth and Overall Health of Patients with SLC13A5 Citrate Transporter Disorder. Metabolites 2021, 11, 746. [Google Scholar] [CrossRef]
- Mishra, D.; Kannan, K.; Meadows, K.; Macro, J.; Li, M.; Frankel, S.; Rogina, B. INDY—From Flies to Worms, Mice, Rats, Non-Human Primates, and Humans. Front. Aging 2021, 2, 782162. [Google Scholar] [CrossRef]
- Inoue, K.; Fei, Y.-J.; Zhuang, L.; Gopal, E.; Miyauchi, S.; Ganapathy, V. Functional Features and Genomic Organization of Mouse NaCT, a Sodium-Coupled Transporter for Tricarboxylic Acid Cycle Intermediates. Biochem. J. 2004, 378, 949–957. [Google Scholar] [CrossRef]
- Hsu, W.-B.; Hsu, W.-H.; Hung, J.-S.; Shen, W.-J.; Hsu, R.W.-W. Transcriptome Analysis of Osteoblasts in an Ovariectomized Mouse Model in Response to Physical Exercise. Bone Joint. Res. 2018, 7, 601–608. [Google Scholar] [CrossRef]
- Zhai, T.; Chen, Q.; Xu, J.; Jia, X.; Xia, P. Prevalence and Trends in Low Bone Density, Osteopenia and Osteoporosis in U.S. Adults with Non-Alcoholic Fatty Liver Disease, 2005–2014. Front. Endocrinol. 2021, 12, 825448. [Google Scholar] [CrossRef]
- Brent, M.B. Pharmaceutical Treatment of Bone Loss: From Animal Models and Drug Development to Future Treatment Strategies. Pharmacol. Ther. 2023, 244, 108383. [Google Scholar] [CrossRef]
- Stoecker, W.V.; Carson, A.; Nguyen, V.H.; Willis, A.B.; Cole, J.G.; Rader, R.K. Addressing the Crisis in the Treatment of Osteoporosis: Better Paths Forward. J. Bone Miner. Res. 2017, 32, 1386–1387. [Google Scholar] [CrossRef]
- Reid, I.R.; Billington, E.O. Drug Therapy for Osteoporosis in Older Adults. Lancet 2022, 399, 1080–1092. [Google Scholar] [CrossRef]
- Pan, B.; Cai, J.; Zhao, P.; Liu, J.; Fu, S.; Jing, G.; Niu, Q.; Li, Q. Relationship between Prevalence and Risk of Osteoporosis or Osteoporotic Fracture with Non-Alcoholic Fatty Liver Disease: A Systematic Review and Meta-Analysis. Osteoporos. Int. 2022, 33, 2275–2286. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Trait | Estimate | Standard Error | p Value |
|---|---|---|---|
| Heel BMD | −5.2377 | 0.205 | 2.314 × 10−177 |
| Vitamin D | 0.0127 | 0.001 | 2.323 × 10−69 |
| Neutrophil/ lymphocyte ratio | 0.1446 | 0.018 | 2.546 × 10−30 |
| 6 Weeks Females | 1 Year Females | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Control | Slc13a5cKO | p-Value | Control | Slc13a5cKO | p-Value | |||||
| Mean | StDev | Mean | StDev | Mean | StDev | Mean | StDev | |||
| Ultimate Moment (Nmm) | 16.306 | 0.681 | 14.853 | 1.279 | 0.010 | 22.754 | 2.613 | 22.960 | 2.226 | 0.864 |
| Bending Rigidity (Nmm2) | 340.344 | 51.487 | 309.758 | 50.248 | 0.212 | 671.574 | 149.997 | 728.670 | 121.604 | 0.406 |
| Ultimate Stress (MPa) | 80.458 | 5.095 | 68.330 | 8.026 | 0.003 | 480.285 | 86.127 | 208.860 | 206.639 | 0.003 |
| Young’s Modulus (MPa) | 3023.970 | 574.458 | 2446.854 | 479.900 | 0.035 | 43,622.800 | 10,573.766 | 19,982.679 | 24,105.944 | 0.017 |
| Ultimate Displacement (mm) | 1.540 | 0.514 | 2.445 | 1.052 | 0.051 | 0.434 | 0.180 | 0.485 | 0.044 | 0.451 |
| Pre-Yield Displacement (mm) | 0.197 | 0.027 | 0.224 | 0.042 | 0.126 | 0.101 | 0.025 | 0.102 | 0.020 | 0.902 |
| Post-Yield Displacement (mm) | 1.345 | 0.533 | 2.221 | 1.041 | 0.058 | 0.334 | 0.166 | 0.383 | 0.048 | 0.433 |
| Ultimate Strain (mm/mm) | 0.220 | 0.078 | 0.349 | 0.146 | 0.048 | 0.034 | 0.015 | 0.065 | 0.019 | 0.002 |
| Pre-Yield Strain (mm/mm) | 0.028 | 0.004 | 0.032 | 0.006 | 0.127 | 0.008 | 0.002 | 0.014 | 0.006 | 0.010 |
| Post-Yield Strain (mm/mm) | 0.243 | 0.161 | 0.317 | 0.145 | 0.305 | 0.026 | 0.013 | 0.051 | 0.014 | 0.002 |
| Ultimate Bending Energy (J) | 10.567 | 3.991 | 11.761 | 3.127 | 0.474 | 6.323 | 1.805 | 5.122 | 0.785 | 0.103 |
| Pre-Yield Energy (J) | 1.069 | 0.214 | 1.133 | 0.303 | 0.618 | 0.585 | 0.213 | 0.588 | 0.215 | 0.978 |
| Post-Yield Energy (J) | 9.498 | 4.033 | 10.628 | 3.058 | 0.496 | 5.738 | 1.916 | 4.534 | 0.674 | 0.113 |
| Toughness (J/mm3) | 10.926 | 2.555 | 13.662 | 4.403 | 0.158 | 19.121 | 9.140 | 8.446 | 6.779 | 0.016 |
| Pre-Yield Toughness (J/mm3) | 1.283 | 0.342 | 1.317 | 0.429 | 0.857 | 1.678 | 0.660 | 0.934 | 0.770 | 0.049 |
| Post-Yield Toughness (J/mm3) | 9.664 | 2.622 | 12.345 | 4.209 | 0.153 | 17.443 | 9.362 | 7.512 | 6.051 | 0.022 |
| 10 Weeks Males | 1 Year Males | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Control | Slc13a5cKO | p-Value | Control | Slc13a5cKO | p-Value | |||||
| Mean | StDev | Mean | StDev | Mean | StDev | Mean | StDev | |||
| Ultimate Moment (Nmm) | 25.723 | 4.099 | 25.756 | 5.869 | 0.989 | 24.204 | 3.372 | 27.381 | 2.881 | 0.035 |
| Bending Rigidity (Nmm2) | 643.472 | 111.209 | 587.465 | 174.759 | 0.437 | 714.243 | 123.466 | 709.637 | 143.268 | 0.940 |
| Ultimate Stress (MPa) | 69.083 | 13.541 | 58.256 | 20.525 | 0.214 | 234.489 | 185.895 | 347.755 | 43.898 | 0.065 |
| Young’s Modulus (MPa) | 563.832 | 157.208 | 434.808 | 176.560 | 0.132 | 16,368.289 | 14,239.261 | 23,053.527 | 5815.719 | 0.171 |
| Ultimate Displacement (mm) | 1.256 | 0.604 | 1.553 | 0.662 | 0.349 | 0.689 | 0.367 | 0.688 | 0.422 | 0.995 |
| Pre-Yield Displacement (mm) | 0.124 | 0.032 | 0.118 | 0.048 | 0.760 | 0.108 | 0.017 | 0.134 | 0.047 | 0.137 |
| Post-Yield Displacement (mm) | 1.131 | 0.587 | 1.435 | 0.664 | 0.333 | 0.581 | 0.362 | 0.554 | 0.400 | 0.878 |
| Ultimate Strain (mm/mm) | 0.953 | 0.465 | 1.144 | 0.467 | 0.412 | 0.089 | 0.051 | 0.067 | 0.043 | 0.305 |
| Pre-Yield Strain (mm/mm) | 0.095 | 0.023 | 0.086 | 0.029 | 0.528 | 0.014 | 0.004 | 0.013 | 0.005 | 0.758 |
| Post-Yield Strain (mm/mm) | 0.859 | 0.452 | 1.058 | 0.478 | 0.392 | 0.076 | 0.049 | 0.054 | 0.040 | 0.293 |
| Ultimate Bending Energy (J) | 12.364 | 3.020 | 14.518 | 4.497 | 0.259 | 7.321 | 3.489 | 7.429 | 4.005 | 0.950 |
| Pre-Yield Energy (J) | 0.717 | 0.265 | 0.725 | 0.374 | 0.956 | 0.710 | 0.179 | 1.046 | 0.625 | 0.138 |
| Post-Yield Energy (J) | 11.647 | 2.995 | 13.792 | 4.654 | 0.271 | 6.611 | 3.421 | 6.383 | 3.686 | 0.889 |
| Toughness (J/mm3) | 45.566 | 18.311 | 43.979 | 22.559 | 0.875 | 12.918 | 12.023 | 16.019 | 8.771 | 0.513 |
| Pre-Yield Toughness (J/mm3) | 2.587 | 0.897 | 2.238 | 1.488 | 0.561 | 1.279 | 0.983 | 2.262 | 1.442 | 0.099 |
| Post-Yield Toughness (J/mm3) | 42.979 | 17.866 | 41.741 | 22.015 | 0.900 | 11.639 | 11.189 | 13.757 | 7.953 | 0.627 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zahn, G.; Baukmann, H.A.; Wu, J.; Jordan, J.; Birkenfeld, A.L.; Dirckx, N.; Schmidt, M.F. Targeting Longevity Gene SLC13A5: A Novel Approach to Prevent Age-Related Bone Fragility and Osteoporosis. Metabolites 2023, 13, 1186. https://doi.org/10.3390/metabo13121186
Zahn G, Baukmann HA, Wu J, Jordan J, Birkenfeld AL, Dirckx N, Schmidt MF. Targeting Longevity Gene SLC13A5: A Novel Approach to Prevent Age-Related Bone Fragility and Osteoporosis. Metabolites. 2023; 13(12):1186. https://doi.org/10.3390/metabo13121186
Chicago/Turabian StyleZahn, Grit, Hannes A. Baukmann, Jasmine Wu, Jens Jordan, Andreas L. Birkenfeld, Naomi Dirckx, and Marco F. Schmidt. 2023. "Targeting Longevity Gene SLC13A5: A Novel Approach to Prevent Age-Related Bone Fragility and Osteoporosis" Metabolites 13, no. 12: 1186. https://doi.org/10.3390/metabo13121186
APA StyleZahn, G., Baukmann, H. A., Wu, J., Jordan, J., Birkenfeld, A. L., Dirckx, N., & Schmidt, M. F. (2023). Targeting Longevity Gene SLC13A5: A Novel Approach to Prevent Age-Related Bone Fragility and Osteoporosis. Metabolites, 13(12), 1186. https://doi.org/10.3390/metabo13121186