

Metabolic Signatures Elucidate the Effect of Body Mass Index on Type 2 Diabetes

 ,
,  ,
,  ,
,  , ,
, ,
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Study Subjects and Sampling
2.2. Metabolite Quantification and Normalization
2.2.1. Multivariable Linear Regression and Logistic Regression
2.2.2. Sobel Mediation Test
2.2.3. Mendelian Randomization
3. Results
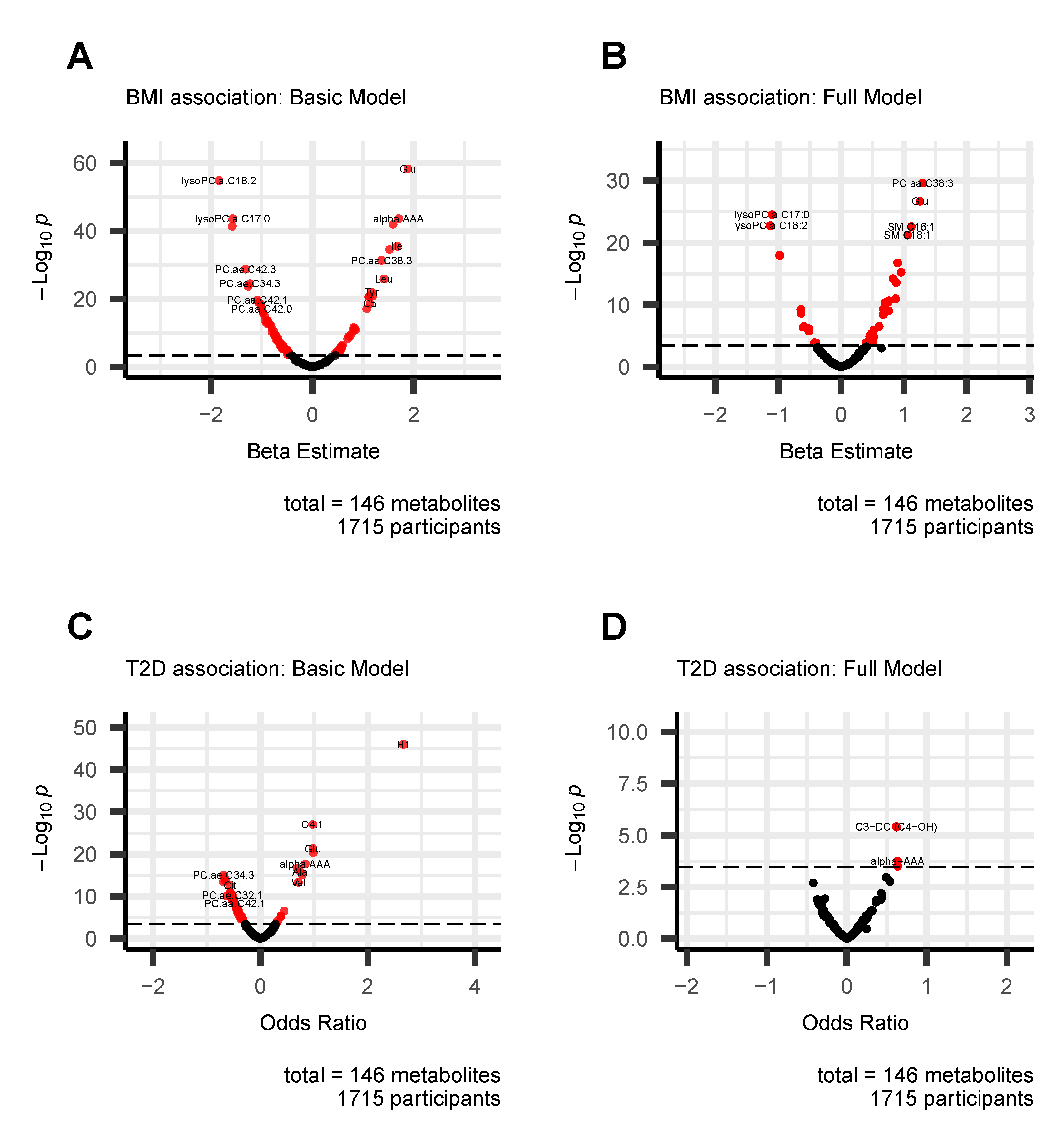
3.1. Associations of Metabolites with BMI and T2D
3.1.1. Characteristics of the KORA FF4 Participants
3.1.2. Metabolites Associated with BMI and T2D
- (1)
- We have observed that all diacyl phosphatidylcholines (PC aa), acylcarnitines, biogenic amines, and sphingomyelins (SM) were positively associated with BMI. In particular, PC aa C38:3 was the strongest metabolite associated with BMI (1.301 [1.082–1.520], q-value = 3.65 × 10−28. Glutamate (1.255 [1.032–1.478], q-value = 3.05 × 10−25), SM C16:1 (1.118 [0.901–1.336], q-value = 3.87 × 10−21), alpha-AAA (0.955 [0.726–1.184], q-value = 8.04 × 10−14), and C0 (0.672 [0.462–0.882], q-value = 6.13 × 10−8) were those with the strongest association in each category;
- (2)
- Some amino acids were positively correlated with BMI. Among them, glutamate (1.255 [1.032–1.478], q-value = 3.05 × 10−25) and Tyrosine (0.901 [0.695–1.106], q-value = 2.51 × 10−15) have the strongest association. Others were inversely associated with BMI: Asparagine (−0.642 [−0.843–−0.44], q-value = 7.73 × 10−8) and Glycine (−0.515 [−0.724–−0.305], q-value = 2.34 × 10−4);
- (3)
- Three acylalkylphosphatidylcholine (PC ae) were positively associated with BMI, PC ae C36:5 (0.502 [0.29–0.713], q-value = 5.09 × 10−4), PC ae C36:4 (0.457 [0.254–0.66], q-value = 1.56 × 10−3), and PC ae C32:2 (0.506 [0.258–0.754], q-value = 9.52 × 10−3); whereas others PC aes were negatively associated with BMI: PC ae C42:3 (−0.594 [−0.821–−0.368], q-value = 4.29 × 10−5), PC ae C36:2 (−0.607 [−0.84–−0.373], q-value = 5.48 × 10−5), PC ae C40:6 (−0.424 [−0.639–−0.209], q-value = 1.66 × 10−2), and PC ae C38:2 (−0.406 [−0.613–−0.199], q-value = 1.80 × 10−2);
- (4)
- All lysophosphatidylcholines (lyso PC) were negatively associated with BMI. In particular, lysoPC a C17:0 (−1.1 [−1.305-−0.896], q-value = 4.20 × 10−23) was the strongest.
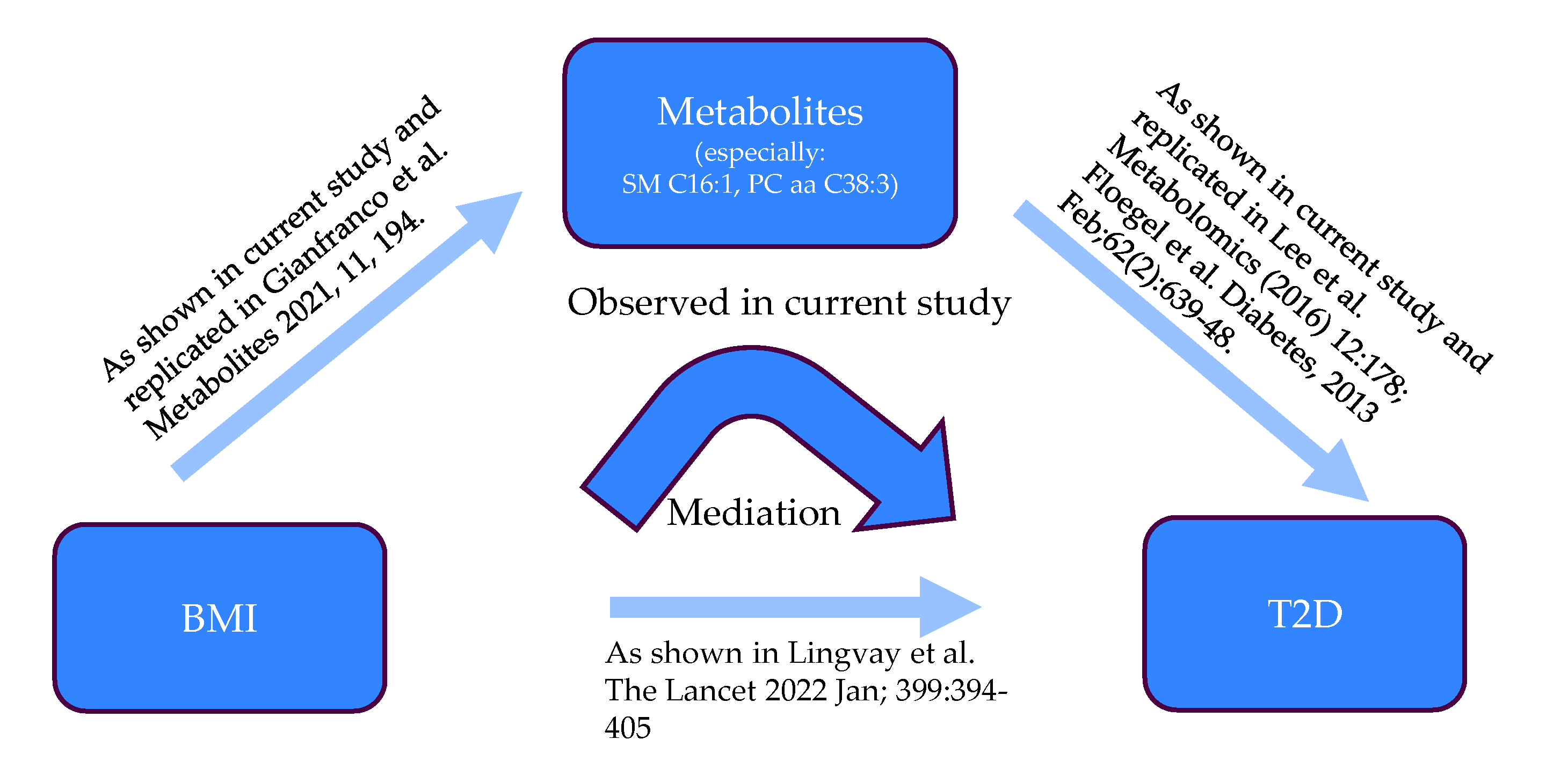
3.2. Sobel Mediation Test
3.3. Mendelian Randomization
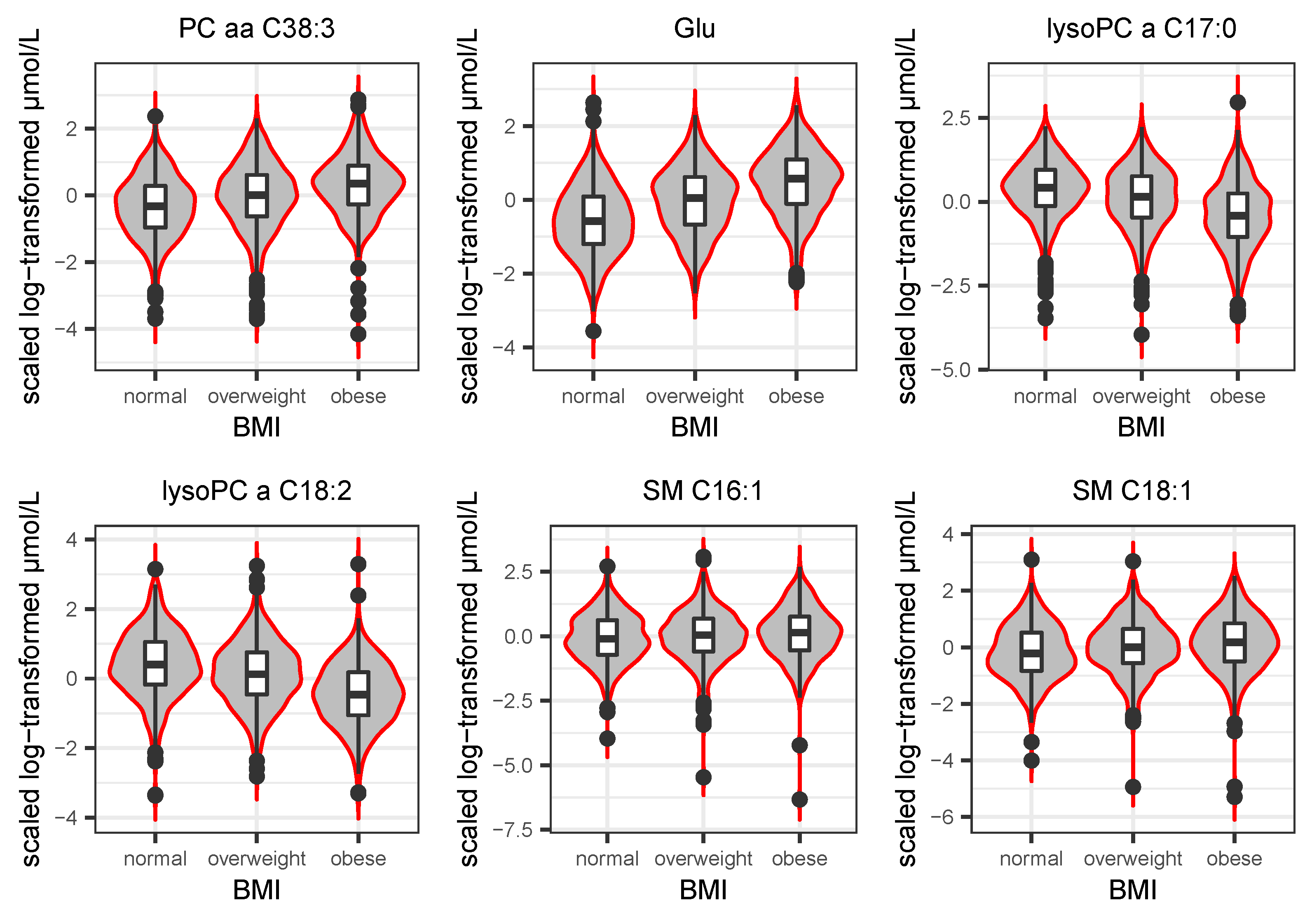
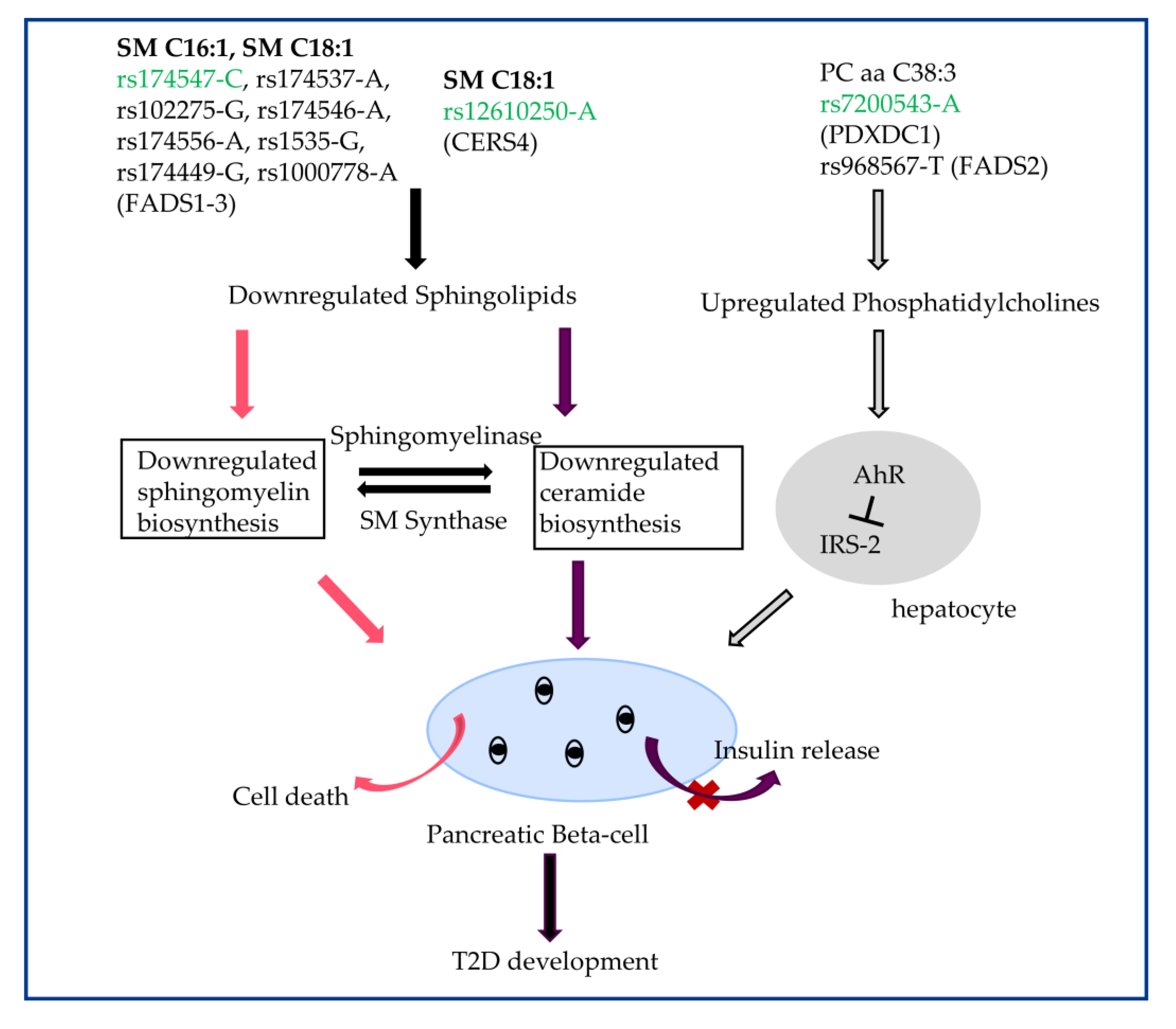
3.4. The Biological Role of SM C16:1, SM C18:1, and PC aa C38:3 in Transition to T2D
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- World Health Organization. Available online: https://www.who.int/news/item/04-03-2022-world-obesity-day-2022-accelerating-action-to-stop-obesity (accessed on 4 March 2022).
- Lingvay, I.; Sumithran, P.; Cohen, R.V.; le Roux, C.W. Obesity management as a primary treatment goal for type 2 diabetes: Time to reframe the conversation. Lancet 2022, 399, 394–405. [Google Scholar] [CrossRef] [PubMed]
- Lauby-Secretan, B.; Scoccianti, C.; Loomis, D.; Grosse, Y.; Bianchini, F.; Straif, K.; Handbook, I.A.R.C. Body Fatness and Cancer—Viewpoint of the IARC Working Group. N. Engl. J. Med. 2016, 375, 794–798. [Google Scholar] [CrossRef]
- Singh, G.M.; Danaei, G.; Farzadfar, F.; Stevens, G.A.; Woodward, M.; Wormser, D.; Kaptoge, S.; Whitlock, G.; Qiao, Q.; Lewington, S.; et al. The Age-Specific Quantitative Effects of Metabolic Risk Factors on Cardiovascular Diseases and Diabetes: A Pooled Analysis. PLoS ONE 2013, 8, e0065174. [Google Scholar] [CrossRef]
- Okamura, T.; Hashimoto, Y.; Hamaguchi, M.; Obora, A.; Kojima, T.; Fukui, M. Ectopic fat obesity presents the greatest risk for incident type 2 diabetes: A population-based longitudinal study. Int. J. Obesity 2019, 43, 139–148. [Google Scholar] [CrossRef] [PubMed]
- Varemo, L.; Nookaew, I.; Nielsen, J. Novel insights into obesity and diabetes through genome-scale metabolic modeling. Front. Physiol. 2013, 4, 92. [Google Scholar] [CrossRef] [PubMed]
- Muoio, D.M.; Newgard, C.B. Obesity-related derangements in metabolic regulation. Annu. Rev. Biochem. 2006, 75, 367–401. [Google Scholar] [CrossRef]
- Park, S.; Sadanala, K.C.; Kim, E.K. A Metabolomic Approach to Understanding the Metabolic Link between Obesity and Diabetes. Mol. Cells 2015, 38, 587–596. [Google Scholar] [CrossRef] [PubMed]
- Stevens, V.L.; Carter, B.D.; McCullough, M.L.; Campbell, P.T.; Wang, Y. Metabolomic Profiles Associated with BMI, Waist Circumference, and Diabetes and Inflammation Biomarkers in Women. Obesity 2020, 28, 187–196. [Google Scholar] [CrossRef]
- Tulipani, S.; Palau-Rodriguez, M.; Alonso, A.M.; Cardona, F.; Marco-Ramell, A.; Zonja, B.; de Alda, M.L.; Munoz-Garach, A.; Sanchez-Pla, A.; Tinahones, F.J.; et al. Biomarkers of Morbid Obesity and Prediabetes by Metabolomic Profiling of Human Discordant Phenotypes. Clin. Chim. Acta 2016, 463, 53–61. [Google Scholar] [CrossRef]
- Rauschert, S.; Uhl, O.; Koletzko, B.; Kirchberg, F.; Mori, T.A.; Huang, R.C.; Beilin, L.J.; Hellmuth, C.; Oddy, W.H. Lipidomics Reveals Associations of Phospholipids With Obesity and Insulin Resistance in Young Adults. J. Clin. Endocr. Metab. 2016, 101, 871–879. [Google Scholar] [CrossRef]
- Sokolowska, E.; Blachnio-Zabielska, A. The Role of Ceramides in Insulin Resistance. Front. Endocrinol. 2019, 10, 577. [Google Scholar] [CrossRef] [PubMed]
- Chang, W.G.; Hatch, G.M.; Wang, Y.; Yu, F.; Wang, M. The relationship between phospholipids and insulin resistance: From clinical to experimental studies. J. Cell Mol. Med. 2019, 23, 702–710. [Google Scholar] [CrossRef]
- Borkman, M.; Storlien, L.H.; Pan, D.A.; Jenkins, A.B.; Chisholm, D.J.; Campbell, L.V. The Relation between Insulin Sensitivity and the Fatty-Acid Composition of Skeletal-Muscle Phospholipids. N. Engl. J. Med. 1993, 328, 238–244. [Google Scholar] [CrossRef] [PubMed]
- Wahl, S.; Drong, A.; Lehne, B.; Loh, M.; Scott, W.R.; Kunze, S.; Tsai, P.C.; Ried, J.S.; Zhang, W.H.; Yang, Y.W.; et al. Epigenome-wide association study of body mass index, and the adverse outcomes of adiposity. Nature 2017, 541, 81. [Google Scholar] [CrossRef]
- Yengo, L.; Sidorenko, J.; Kemper, K.E.; Zheng, Z.; Wood, A.R.; Weedon, M.N.; Frayling, T.M.; Hirschhorn, J.; Yang, J.; Visscher, P.M.; et al. Meta-analysis of genome-wide association studies for height and body mass index in approximately 700000 individuals of European ancestry. Hum. Mol. Genet. 2018, 27, 3641–3649. [Google Scholar] [CrossRef] [PubMed]
- Mahajan, A.; Spracklen, C.N.; Zhang, W.; Ng, M.C.Y.; Petty, L.E.; Kitajima, H.; Yu, G.Z.; Rueger, S.; Speidel, L.; Kim, Y.J.; et al. Multi-ancestry genetic study of type 2 diabetes highlights the power of diverse populations for discovery and translation. Nat. Genet. 2022, 54, 560–572. [Google Scholar] [CrossRef] [PubMed]
- Lanznaster, D.; Veyrat-Durebex, C.; Vourc’h, P.; Andres, C.R.; Blasco, H.; Corcia, P. Metabolomics: A Tool to Understand the Impact of Genetic Mutations in Amyotrophic Lateral Sclerosis. Genes 2020, 11, 537. [Google Scholar] [CrossRef]
- Gieger, C.; Geistlinger, L.; Altmaier, E.; Hrabe de Angelis, M.; Kronenberg, F.; Meitinger, T.; Mewes, H.W.; Wichmann, H.E.; Weinberger, K.M.; Adamski, J.; et al. Genetics meets metabolomics: A genome-wide association study of metabolite profiles in human serum. PLoS Genet. 2008, 4, e1000282. [Google Scholar] [CrossRef]
- Suhre, K.; Raffler, J.; Kastenmuller, G. Biochemical insights from population studies with genetics and metabolomics. Arch. Biochem. Biophys. 2016, 589, 168–176. [Google Scholar] [CrossRef]
- Wang-Sattler, R.; Yu, Z.; Herder, C.; Messias, A.C.; Floegel, A.; He, Y.; Heim, K.; Campillos, M.; Holzapfel, C.; Thorand, B.; et al. Novel biomarkers for pre-diabetes identified by metabolomics. Mol. Syst. Biol. 2012, 8, 615. [Google Scholar] [CrossRef]
- Haid, M.; Muschet, C.; Wahl, S.; Romisch-Margl, W.; Prehn, C.; Moller, G.; Adamski, J. Long-Term Stability of Human Plasma Metabolites during Storage at -80 degrees C. J. Proteome Res. 2018, 17, 203–211. [Google Scholar] [CrossRef] [PubMed]
- Zukunft, S.; Prehn, C.; Rohring, C.; Moller, G.; Hrabe de Angelis, M.; Adamski, J.; Tokarz, J. High-throughput extraction and quantification method for targeted metabolomics in murine tissues. Metabolomics 2018, 14, 18. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Covic, M.; Huth, C.; Rommel, M.; Adam, J.; Zukunft, S.; Prehn, C.; Wang, L.; Nano, J.; Scheerer, M.F.; et al. Validation of Candidate Phospholipid Biomarkers of Chronic Kidney Disease in Hyperglycemic Individuals and Their Organ-Specific Exploration in Leptin Receptor-Deficient db/db Mouse. Metabolites 2021, 11, 89. [Google Scholar] [CrossRef]
- Preacher, K.J.; Hayes, A.F. Asymptotic and resampling strategies for assessing and comparing indirect effects in multiple mediator models. Behav. Res. Methods 2008, 40, 879–891. [Google Scholar] [CrossRef]
- Sobel, M.E. Asymptotic confidence intervals for indirect effects in structural equation models. Sociol. Methodol. 1982, 13, 290–312. [Google Scholar] [CrossRef]
- Hemani, G.; Zhengn, J.; Elsworth, B.; Wade, K.H.; Haberland, V.; Baird, D.; Laurin, C.; Burgess, S.; Bowden, J.; Langdon, R.; et al. The MR-Base platform supports systematic causal inference across the human phenome. eLife 2018, 7, e34408. [Google Scholar] [CrossRef]
- Draisma, H.H.M.; Pool, R.; Kobl, M.; Jansen, R.; Petersen, A.K.; Vaarhorst, A.A.M.; Yet, I.; Haller, T.; Demirkan, A.; Esko, T.; et al. Genome-wide association study identifies novel genetic variants contributing to variation in blood metabolite levels. Nat. Commun. 2015, 6, 7208. [Google Scholar] [CrossRef] [PubMed]
- Xue, A.; Wu, Y.; Zhu, Z.; Zhang, F.; Kemper, K.E.; Zheng, Z.; Yengo, L.; Lloyd-Jones, L.R.; Sidorenko, J.; Wu, Y.; et al. Genome-wide association analyses identify 143 risk variants and putative regulatory mechanisms for type 2 diabetes. Nat. Commun. 2018, 9, 2941. [Google Scholar] [CrossRef]
- Frigerio, G.; Favero, C.; Savino, D.; Mercadante, R.; Albetti, B.; Dioni, L.; Vigna, L.; Bollati, V.; Pesatori, A.C.; Fustinoni, S. Plasma Metabolomic Profiling in 1391 Subjects with Overweight and Obesity from the SPHERE Study. Metabolites 2021, 11, 194. [Google Scholar] [CrossRef]
- Lee, H.-S.; Xu, T.; Lee, Y.; Kim, N.-H.; Kim, Y.-J.; Kim, J.-M.; Cho, S.Y.; Kim, K.-Y.; Nam, M.; Adamski, J.; et al. Identification of putative biomarkers for type 2 diabetes using metabolomics in the Korea Association REsource (KARE) cohort. Metabolomics 2016, 12, 178. [Google Scholar] [CrossRef]
- Floegel, A.; Stefan, N.; Yu, Z.; Muhlenbruch, K.; Drogan, D.; Joost, H.G.; Fritsche, A.; Haring, H.U.; Hrabe de Angelis, M.; Peters, A.; et al. Identification of serum metabolites associated with risk of type 2 diabetes using a targeted metabolomic approach. Diabetes 2013, 62, 639–648. [Google Scholar] [CrossRef] [PubMed]
- Hicks, A.A.; Pramstaller, P.P.; Johansson, A.; Vitart, V.; Rudan, I.; Ugocsai, P.; Aulchenko, Y.; Franklin, C.S.; Liebisch, G.; Erdmann, J.; et al. Genetic determinants of circulating sphingolipid concentrations in European populations. PLoS Genet. 2009, 5, e1000672. [Google Scholar] [CrossRef] [PubMed]
- Straczkowski, M.; Kowalska, I.; Nikolajuk, A.; Dzienis-Straczkowska, S.; Kinalska, I.; Baranowski, M.; Zendzian-Piotrowska, M.; Brzezinska, Z.; Gorski, J. Relationship between insulin sensitivity and sphingomyelin signaling pathway in human skeletal muscle. Diabetes 2004, 53, 1215–1221. [Google Scholar] [CrossRef] [PubMed]
- Subathra, M.; Qureshi, A.; Luberto, C. Sphingomyelin Synthases Regulate Protein Trafficking and Secretion. PLoS ONE 2011, 6, e23644. [Google Scholar] [CrossRef] [PubMed]
- Khan, S.R.; Manialawy, Y.; Obersterescu, A.; Cox, B.J.; Gunderson, E.P.; Wheeler, M.B. Diminished Sphingolipid Metabolism, a Hallmark of Future Type 2 Diabetes Pathogenesis, Is Linked to Pancreatic beta Cell Dysfunction. iScience 2020, 23, 101566. [Google Scholar] [CrossRef]
- Alexaki, A.; Clarke, B.A.; Gavrilova, O.; Ma, Y.; Zhu, H.; Ma, X.; Xu, L.; Tuymetova, G.; Larman, B.C.; Allende, M.L.; et al. De Novo Sphingolipid Biosynthesis Is Required for Adipocyte Survival and Metabolic Homeostasis. J. Biol. Chem. 2017, 292, 3929–3939. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Sundaram, K.; Mu, J.Y.; Dryden, G.W.; Sriwastva, M.K.; Lei, C.; Zhang, L.F.; Qiu, X.L.; Xu, F.Y.; Yan, J.; et al. High-fat diet-induced upregulation of exosomal phosphatidylcholine contributes to insulin resistance. Nat. Commun. 2021, 12. [Google Scholar] [CrossRef] [PubMed]
- Lynch, C.J.; Adams, S.H. Branched-chain amino acids in metabolic signalling and insulin resistance. Nat. Rev. Endocrinol. 2014, 10, 723–736. [Google Scholar] [CrossRef]
- Yoon, M.S. The Emerging Role of Branched-Chain Amino Acids in Insulin Resistance and Metabolism. Nutrients 2016, 8, 405. [Google Scholar] [CrossRef]
- Wang, S.M.; Yang, R.Y.; Wang, M.; Ji, F.S.; Li, H.X.; Tang, Y.M.; Chen, W.X.; Dong, J. Identification of serum metabolites associated with obesity and traditional risk factors for metabolic disease in Chinese adults. Nutr. Metab. Cardiovas. 2018, 28, 112–118. [Google Scholar] [CrossRef]
- Bagheri, M.; Djazayery, A.; Farzadfar, F.; Qi, L.; Yekaninejad, M.S.; Aslibekyan, S.; Chamari, M.; Hassani, H.; Koletzko, B.; Uhl, O. Plasma metabolomic profiling of amino acids and polar lipids in Iranian obese adults. Lipids Health Dis. 2019, 18, 94. [Google Scholar] [CrossRef] [PubMed]
- Rangel-Huerta, O.D.; Pastor-Villaescusa, B.; Gil, A. Are we close to defining a metabolomic signature of human obesity? A systematic review of metabolomics studies. Metabolomics 2019, 15, 93. [Google Scholar] [CrossRef]
- Libert, D.M.; Nowacki, A.S.; Natowicz, M.R. Metabolomic analysis of obesity, metabolic syndrome, and type 2 diabetes: Amino acid and acylcarnitine levels change along a spectrum of metabolic wellness. PeerJ 2018, 6, e5410. [Google Scholar] [CrossRef] [PubMed]
- Siddik, M.A.; Shin, A.C. Recent Progress on Branched-Chain Amino Acids in Obesity, Diabetes, and Beyond. Endocrinol. Metab. 2019, 34, 234–246. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Han, Q.; Liu, Y.J.; Sun, C.L.; Gang, X.K.; Wang, G.X. The Relationship between Branched-Chain Amino Acid Related Metabolomic Signature and Insulin Resistance: A Systematic Review. J. Diabetes Res. 2016, 2016, 2794591. [Google Scholar] [CrossRef]
- Adams, S.H. Emerging Perspectives on Essential Amino Acid Metabolism in Obesity and the Insulin-Resistant State. Adv. Nutr. 2011, 2, 445–456. [Google Scholar] [CrossRef]
- Fikri, A.M.; Smyth, R.; Kumar, V.; Al-Abadla, Z.; Abusnana, S.; Munday, M.R. Pre-diagnostic biomarkers of type 2 diabetes identified in the UAE’s obese national population using targeted metabolomics. Sci. Rep. 2020, 10, 17616. [Google Scholar] [CrossRef] [PubMed]
- Adams, S.H.; Hoppel, C.L.; Lok, K.H.; Zhao, L.; Wong, S.W.; Minkler, P.E.; Hwang, D.H.; Newman, J.W.; Garvey, W.T. Plasma acylcarnitine profiles suggest incomplete long-chain fatty acid beta-oxidation and altered tricarboxylic acid cycle activity in type 2 diabetic African-American women. J. Nutr. 2009, 139, 1073–1081. [Google Scholar] [CrossRef]
- Wijekoon, E.P.; Skinner, C.; Brosnan, M.E.; Brosnan, J.T. Amino acid metabolism in the Zucker diabetic fatty rat: Effects of insulin resistance and of type 2 diabetes. Can. J. Physiol. Pharm. 2004, 82, 506–514. [Google Scholar] [CrossRef]
- Carayol, M.; Leitzmann, M.F.; Ferrari, P.; Zamora-Ros, R.; Achaintre, D.; Stepien, M.; Schmidt, J.A.; Travis, R.C.; Overvad, K.; Tjonneland, A.; et al. Blood Metabolic Signatures of Body Mass Index: A Targeted Metabolomics Study in the EPIC Cohort. J. Proteome Res. 2017, 16, 3137–3146. [Google Scholar] [CrossRef]
- Cole, L.K.; Vance, J.E.; Vance, D.E. Phosphatidylcholine biosynthesis and lipoprotein metabolism. BBA Mol. Cell. Biol. Lipids 2012, 1821, 754–761. [Google Scholar] [CrossRef]
- Matsumoto, T.; Kobayashi, T.; Kamata, K. Role of lysophosphatidylcholine (LPC) in atherosclerosis. Curr. Med. Chem. 2007, 14, 3209–3220. [Google Scholar] [CrossRef]
- Pacana, T.; Cazanave, S.; Verdianelli, A.; Patel, V.; Min, H.K.; Mirshahi, F.; Quinlivan, E.; Sanyal, A.J. Dysregulated Hepatic Methionine Metabolism Drives Homocysteine Elevation in Diet-Induced Nonalcoholic Fatty Liver Disease. PLoS ONE 2015, 10, e0136822. [Google Scholar] [CrossRef] [PubMed]
- Wei, X.C.; Song, H.W.; Yin, L.; Rizzo, M.G.; Sidhu, R.; Covey, D.F.; Ory, D.S.; Semenkovich, C.F. Fatty acid synthesis configures the plasma membrane for inflammation in diabetes. Nature 2016, 539, 294. [Google Scholar] [CrossRef] [PubMed]
- van der Veen, J.N.; Lingrell, S.; McCloskey, N.; LeBlond, N.D.; Galleguillos, D.; Zhao, Y.Y.; Curtis, J.M.; Sipione, S.; Fullerton, M.D.; Vance, D.E.; et al. A role for phosphatidylcholine and phosphatidylethanolamine in hepatic insulin signaling. FASEB J. 2019, 33, 5045–5057. [Google Scholar] [CrossRef]
- Kim, Y.C.; Seok, S.; Byun, S.; Kong, B.; Zhang, Y.; Guo, G.; Xie, W.; Ma, J.; Kemper, B.; Kemper, J.K. AhR and SHP regulate phosphatidylcholine and S-adenosylmethionine levels in the one-carbon cycle. Nat. Commun. 2018, 9, 540. [Google Scholar] [CrossRef]
- Park, T.S.; Panek, R.L.; Mueller, S.B.; Hanselman, J.C.; Rosebury, W.S.; Robertson, A.W.; Kindt, E.K.; Homan, R.; Karathanasis, S.K.; Rekhter, M.D. Inhibition of sphingomyelin synthesis reduces atherogenesis in apolipoprotein E-knockout mice. Circulation 2004, 110, 3465–3471. [Google Scholar] [CrossRef]
- Russo, S.B.; Ross, J.S.; Cowart, L.A. Sphingolipids in Obesity, Type 2 Diabetes, and Metabolic Disease; Springer: Vienna, Austria, 2013; pp. 373–401. [Google Scholar] [CrossRef]
- Hammad, S.M.; Pierce, J.S.; Soodavar, F.; Smith, K.J.; Al Gadban, M.M.; Rembiesa, B.; Klein, R.L.; Hannun, Y.A.; Bielawski, J.; Bielawska, A. Blood sphingolipidomics in healthy humans: Impact of sample collection methodology. J. Lipid Res. 2010, 51, 3074–3087. [Google Scholar] [CrossRef]
- Shimabukuro, M.; Zhou, Y.T.; Levi, M.; Unger, R.H. Fatty acid-induced beta cell apoptosis: A link between obesity and diabetes. Proc. Natl. Acad. Sci. USA 1998, 95, 2498–2502. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, W.; Ross, J.; Geng, T.Y.; Brice, S.E.; Cowart, L.A. Differential Regulation of Dihydroceramide Desaturase by Palmitate versus Monounsaturated Fatty Acids IMPLICATIONS FOR INSULIN RESISTANCE. J. Biol. Chem. 2011, 286, 16596–16605. [Google Scholar] [CrossRef]
- Holland, W.L.; Bikman, B.T.; Wang, L.P.; Yuguang, G.; Sargent, K.M.; Bulchand, S.; Knotts, T.A.; Shui, G.H.; Clegg, D.J.; Wenk, M.R.; et al. Lipid-induced insulin resistance mediated by the proinflammatory receptor TLR4 requires saturated fatty acid-induced ceramide biosynthesis in mice. J. Clin. Investig. 2011, 121, 1858–1870. [Google Scholar] [CrossRef]
- Samad, F.; Hester, K.D.; Yang, G.; Hannun, Y.A.; Bielawski, J. Altered adipose and plasma sphingolipid metabolism in obesity - A potential mechanism for cardiovascular and metabolic risk. Diabetes 2006, 55, 2579–2587. [Google Scholar] [CrossRef]
- Sas, K.M.; Nair, V.; Byun, J.; Kayampilly, P.; Zhang, H.; Saha, J.; Brosius, F.C., 3rd; Kretzler, M.; Pennathur, S. Targeted Lipidomic and Transcriptomic Analysis Identifies Dysregulated Renal Ceramide Metabolism in a Mouse Model of Diabetic Kidney Disease. J. Proteom. Bioinform. 2015, 2015 (Suppl. 14), 2. [Google Scholar] [CrossRef]
- Lee, S.Y.; Lee, H.Y.; Song, J.H.; Kim, G.T.; Jeon, S.; Song, Y.J.; Lee, J.S.; Hur, J.H.; Oh, H.H.; Park, S.Y.; et al. Adipocyte-Specific Deficiency of De Novo Sphingolipid Biosynthesis Leads to Lipodystrophy and Insulin Resistance. Diabetes 2017, 66, 2596–2609. [Google Scholar] [CrossRef] [PubMed]
- Park, J.W.; Park, W.J.; Kuperman, Y.; Boura-Halfon, S.; Pewzner-Jung, Y.; Futerman, A.H. Ablation of Very Long Acyl Chain Sphingolipids Causes Hepatic Insulin Resistance in Mice Due to Altered Detergent-Resistant Membranes. Hepatology 2013, 57, 525–532. [Google Scholar] [CrossRef] [PubMed]
- Lemaitre, R.N.; Tanaka, T.; Tang, W.H.; Manichaikul, A.; Foy, M.; Kabagambe, E.K.; Nettleton, J.A.; King, I.B.; Weng, L.C.; Bhattacharya, S.; et al. Genetic Loci Associated with Plasma Phospholipid n-3 Fatty Acids: A Meta-Analysis of Genome-Wide Association Studies from the CHARGE Consortium. PloS Genet. 2011, 7, e1002193. [Google Scholar] [CrossRef] [PubMed]
- Guan, W.H.; Steffen, B.T.; Lemaitre, R.N.; Wu, J.H.Y.; Tanaka, T.; Manichaikul, A.; Foy, M.; Rich, S.S.; Wang, L.; Nettleton, J.A.; et al. Genome-Wide Association Study of Plasma N6 Polyunsaturated Fatty Acids Within the Cohorts for Heart and Aging Research in Genomic Epidemiology Consortium. Circ. Cardiovasc. Genet. 2014, 7, 321–331. [Google Scholar] [CrossRef]
- Meierhofer, D.; Weidner, C.; Sauer, S. Integrative Analysis of Transcriptomics, Proteomics, and Metabolomics Data of White Adipose and Liver Tissue of High-Fat Diet and Rosiglitazone-Treated Insulin-Resistant Mice Identified Pathway Alterations and Molecular Hubs. J. Proteome Res. 2014, 13, 5592–5602. [Google Scholar] [CrossRef]
- Lattka, E.; Illig, T.; Koletzko, B.; Heinrich, J. Genetic variants of the FADS1 FADS2 gene cluster as related to essential fatty acid metabolism. Curr. Opin. Lipidol. 2010, 21, 64–69. [Google Scholar] [CrossRef]
- Reardon, H.T.; Hsieh, A.T.; Park, W.J.; Kothapalli, K.S.D.; Anthony, J.C.; Nathanielsz, P.W.; Brenna, J.T. Dietary long-chain polyunsaturated fatty acids upregulate expression of FADS3 transcripts. Prostag. Leukotr. Ess. 2013, 88, 15–19. [Google Scholar] [CrossRef]
- Dupuis, J.; Langenberg, C.; Prokopenko, I.; Saxena, R.; Soranzo, N.; Jackson, A.U.; Wheeler, E.; Glazer, N.L.; Bouatia-Naji, N.; Gloyn, A.L.; et al. New genetic loci implicated in fasting glucose homeostasis and their impact on type 2 diabetes risk (vol 42, pg 105, 2010). Nat. Genet. 2010, 42, 464. [Google Scholar] [CrossRef]
- Martinelli, N.; Girelli, D.; Malerba, G.; Guarini, P.; Illig, T.; Trabetti, E.; Sandri, M.; Friso, S.; Pizzolo, F.; Schaeffer, L.; et al. FADS genotypes and desaturase activity estimated by the ratio of arachidonic acid to linoleic acid are associated with inflammation and coronary artery disease. Am. J. Clin. Nutr. 2008, 88, 941–949. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Athinarayanan, S.; Jiang, G.; Chalasani, N.; Zhang, M.; Liu, W. Fatty acid desaturase 1 gene polymorphisms control human hepatic lipid composition. Hepatology 2015, 61, 119–128. [Google Scholar] [CrossRef] [PubMed]
- Meldrum, S.J.; Li, Y.C.; Zhang, G.C.; Heaton, A.E.M.; D’Vaz, N.; Manz, J.; Reischl, E.; Koletzko, B.V.; Prescott, S.L.; Simmer, K. Can polymorphisms in the fatty acid desaturase (FADS) gene cluster alter the effects of fish oil supplementation on plasma and erythrocyte fatty acid profiles? An exploratory study. Eur. J. Nutr. 2018, 57, 2583–2594. [Google Scholar] [CrossRef] [PubMed]
- Brayner, B.; Kaur, G.; Keske, M.A.; Livingstone, K.M. FADS Polymorphism, Omega-3 Fatty Acids and Diabetes Risk: A Systematic Review. Nutrients 2018, 10, 758. [Google Scholar] [CrossRef] [PubMed]
- Ralston, J.C.; Matravadia, S.; Gaudio, N.; Holloway, G.P.; Mutch, D.M. Polyunsaturated Fatty Acid Regulation of Adipocyte FADS1 and FADS2 Expression and Function. Obesity 2015, 23, 725–728. [Google Scholar] [CrossRef]
- Stancakova, A.; Paananen, J.; Soininen, P.; Kangas, A.J.; Bonnycastle, L.L.; Morken, M.A.; Collins, F.S.; Jackson, A.U.; Boehnke, M.L.; Kuusisto, J.; et al. Effects of 34 risk loci for type 2 diabetes or hyperglycemia on lipoprotein subclasses and their composition in 6,580 nondiabetic Finnish men. Diabetes 2011, 60, 1608–1616. [Google Scholar] [CrossRef]
- Khamlaoui, W.; Mehri, S.; Hammami, S.; Hammouda, S.; Chraeif, I.; Elosua, R.; Hammami, M. Association Between Genetic Variants in FADS1-FADS2 and ELOVL2 and Obesity, Lipid Traits, and Fatty Acids in Tunisian Population. Clin. Appl.Thromb.-Hem. 2020, 26, 1076029620915286. [Google Scholar] [CrossRef] [PubMed]
- Bult, C.J.; Blake, J.A.; Smith, C.L.; Kadin, J.A.; Richardson, J.E.; Anagnostopoulos, A.; Asabor, R.; Baldarelli, R.M.; Beal, J.S.; Bello, S.M.; et al. Mouse Genome Database (MGD) 2019. Nucleic Acids Res. 2019, 47, D801–D806. [Google Scholar] [CrossRef] [Green Version]
- Yang, X.; Schadt, E.E.; Wang, S.; Wang, H.; Arnold, A.P.; Ingram-Drake, L.; Drake, T.A.; Lusis, A.J. Tissue-specific expression and regulation of sexually dimorphic genes in mice. Genome Res. 2006, 16, 995–1004. [Google Scholar] [CrossRef]
- Langfelder, P.; Horvath, S. WGCNA: An R package for weighted correlation network analysis. BMC Bioinform. 2008, 9, 559. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- James, G.; Witten, D.; Hastie, T.; Tibshirani, R. An Introduction to Statistical Learning: With Applications in R, Uncorrected ed.; Springer: New York, NY, USA, 2013. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| BMI | Overall | Non-Obese (BMI < 30 kg/m2) | Obese (BMI ≥ 30 kg/m2) | p Value |
|---|---|---|---|---|
| Sample size | 1715 | 1276 | 439 | |
| Age mean (SD) | 59.0 (12.2) | 58.1 (12.1) | 62.0 (12.0) | <0.001 |
| Sex woman (%) | 904 (52.7) | 683 (53.5) | 221 (50.3) | 0.268 |
| Weight (kg) mean (SD) | 78.6 (16.0) | 72.7 (11.8) | 95.7 (14.2) | <0.001 |
| Height (cm) mean (SD) | 169.1 (9.6) | 169.5 (9.6) | 167.7 (9.7) | <0.001 |
| Alcohol (g/day) mean (SD) | 14.2 (19.4) | 14.5 (18.2) | 13.5 (22.5) | 0.392 |
| Waist (cm) mean (SD) | 95.6 (14.0) | 90.2 (10.5) | 111.6 (10.3) | <0.001 |
| Waist-hip-ratio mean (SD) | 0.9 (0.1) | 0.88 (0.1) | 0.96 (0.1) | <0.001 |
| Fasting glucose (mmol/L) mean (SD) | 5.6 (1.3) | 5.4 (1.0) | 6.3 (1.7) | <0.001 |
| 2 h post glucose (mmol/L) mean (SD) | 5.8 (2.2) | 5.5 (1.7) | 6.9 (3.2) | <0.001 |
| Systolic blood pressure (mmHg) mean (SD) | 117.9 (17.2) | 116.4 (16.6) | 122.5 (18.1) | <0.001 |
| Diastolic blood pressure (mmHg) mean (SD) | 72.7 (9.5) | 72.2 (9.1) | 74.0 (10.3) | 0.001 |
| Smoking (%) | <0.001 | |||
| Smoker | 267 (15.6) | 221 (17.3) | 46 (10.5) | |
| Ex-smoker | 658 (38.4) | 461 (36.1) | 197 (44.9) | |
| Never-smoker | 790 (46.1) | 594 (46.6) | 196 (44.6) | |
| Physical activities inactive (%) | 702 (40.9) | 456 (35.7) | 246 (56.0) | <0.001 |
| HDL cholesterol (mmol/L) mean (SD) | 1.7 (0.5) | 1.8 (0.5) | 1.5 (0.4) | <0.001 |
| LDL cholesterol (mmol/L) mean (SD) | 3.5 (0.9) | 3.4 (0.9) | 3.6 (0.9) | 0.048 |
| Triglycerides (mmol/L) mean (SD) | 1.4 (0.8) | 1.25 (0.8) | 1.6 (0.9) | <0.001 |
| HbA1c (%) mean (SD) | 5.5 (0.7) | 5.4 (0.6) | 5.8 (0.9) | <0.001 |
| Total cholesterol (mmol/L) mean (SD) | 5.6 (1.00) | 5.6 (1.0) | 5.5 (1.0) | 0.409 |
| C-reactive protein (mg/L) mean (SD) | 2.3 (4.4) | 1.7 (3.8) | 3.9 (5.5) | <0.001 |
| Type 2 diabetesy (%) | 300 (17.5) | 136 (10.7) | 164 (37.4) | <0.001 |
| Diabetes | Overall | T2D (No) | T2D (Yes) | p Value |
|---|---|---|---|---|
| Sample size | 1715 | 1415 | 300 | |
| Age mean (SD) | 59.0 (12.2) | 59.7 (12.2) | 69.5 (10.0) | <0.001 |
| Sex woman (%) | 904 (52.7) | 784 (55.4) | 120 (40.0) | <0.001 |
| Weight (kg) mean (SD) | 78.6 (16.0) | 76.8 (15.3) | 87.2 (16.5) | <0.001 |
| Height (cm) mean (SD) | 169.1 (9.6) | 169.4 (9.7) | 167.2 (9.1) | <0.001 |
| Alcohol (g/day) mean (SD) | 14.2 (19.4) | 13.9 (18.1) | 15.8 (24.7) | 0.115 |
| Waist (cm) mean (SD) | 95.6 (14.0) | 93.1 (12.9) | 107.8 (12.7) | <0.001 |
| Waist-hip-ratio mean (SD) | 0.9 (0.1) | 0.9 (0.1) | 1.0 (0.1) | <0.001 |
| Fasting glucose (mmol/L) mean (SD) | 5.6 (1.3) | 5.2 (0.4) | 7.6 (2.0) | <0.001 |
| 2 h post glucose (mmol/L) mean (SD) | 5.8 (2.2) | 5.4 (1.1) | 12.6 (3.5) | <0.001 |
| Systolic blood pressure (mmHg) mean (SD) | 117.9 (17.2) | 116.1 (16.2) | 126.7 (18.8) | <0.001 |
| Diastolic blood pressure (mmHg) mean (SD) | 72.7 (9.5) | 72.8 (9.1) | 72.0 (11.1) | 0.201 |
| Smoking (%) | <0.001 | |||
| Smoker | 267 (15.6) | 243 (17.2) | 24 (8.0) | |
| Ex-smoker | 658 (38.4) | 524 (37.0) | 134 (44.7) | |
| Never-smoker | 790 (46.1) | 648 (45.8) | 142 (47.3) | |
| Physical activities inactive (%) | 702 (40.9) | 512 (36.2) | 190 (63.3) | |
| HDL cholesterol (mmol/L) mean (SD) | 1.72 (0.5) | 1.76 (0.5) | 1.48 (0.4) | <0.001 |
| LDL cholesterol (mmol/L) mean (SD) | 3.5 (0.9) | 3.5 (0.9) | 3.3 (0.9) | <0.001 |
| Triglycerides (mmol/L) mean (SD) | 1.4 (0.8) | 1.3 (0.8) | 1.8 (1.0) | <0.001 |
| HbA1c (%) mean (SD) | 5.5 (0.7) | 5.3 (0.3) | 6.5 (1.0) | <0.001 |
| Total cholesterol (mmol/L) mean (SD) | 5.6 (1.0) | 5.6 (1.0) | 5.3 (1.1) | <0.001 |
| C-reactive protein (mg/L) mean (SD) | 2.3 (4.4) | 2.1 (4.3) | 3.4 (4.6) | <0.001 |
| BMI = Obese (%) | 439 (25.6) | 275 (19.4) | 164 (54.7) | <0.001 |
| Positively Associated | ||||
|---|---|---|---|---|
| Category | Metabolite | Beta Estimate (95% CI) | p-value | q-value |
| PC aa | PC aa C38:3 | 1.301 (1.082–1.520) | 2.50 × 10−30 | 3.65 × 10−28 |
| PC aa | PC aa C38:4 | 0.728 (0.514–0.943) | 3.74 × 10−11 | 5.47 × 10−9 |
| PC aa | PC aa C40:4 | 0.692 (0.471–0.913) | 9.89 × 10−11 | 1.44 × 10−7 |
| PC aa | PC aa C32:1 | 0.606 (0.375–0.837) | 2.93 × 10−7 | 4.28 × 10−5 |
| PC aa | PC aa C40:5 | 0.505 (0.279–0.730) | 1.19 × 10−5 | 1.74 × 10−3 |
| PC aa | PC aa C36:3 | 0.512 (0.281–0.742) | 1.41 × 10−5 | 2.06 × 10−3 |
| PC aa | PC aa C36:4 | 0.426 (0.207–0.644) | 1.38 × 10−4 | 2.01 × 10−2 |
| Amino Acids | Glutamate (Glu) | 1.255 (1.032–1.478) | 2.09 × 10−27 | 3.05 × 10−25 |
| Amino Acids | Tyrosine (Tyr) | 0.901 (0.695–1.106) | 1.72 × 10−17 | 2.51 × 10−15 |
| Amino Acids | Phenylalanine (Phe) | 0.823 (0.618–1.027) | 6.11 × 10−15 | 8.92 × 10−13 |
| Amino Acids | Valine (Val) | 0.876 (0.652–1.100) | 2.60 × 10−14 | 3.80 × 10−12 |
| Amino Acids | Isoleucine (Ile) | 0.866 (0.618–1.114) | 1.05 × 10−11 | 1.53 × 10−9 |
| Amino Acids | Leucine (Leu) | 0.755 (0.515–0.995) | 9.02 × 10−10 | 1.32 × 10−7 |
| Amino Acids | Alanine (Ala) | 0.458 (0.242–0.673) | 3.27 × 10−5 | 4.78 × 10−3 |
| Amino Acids | Ornithine (Orn) | 0.399 (0.195–0.603) | 1.30 × 10−4 | 1.90 × 10−2 |
| SM | SM C16:1 | 1.118 (0.901–1.336) | 2.65 × 10−23 | 3.87 × 10−21 |
| SM | SM C18:1 | 1.061 (0.848–1.273) | 5.81 × 10−22 | 8.48 × 10−20 |
| SM | SM C20:2 | 0.763 (0.541–0.985) | 2.14 × 10−11 | 3.12 × 10−9 |
| SM | SM C18:0 | 0.697 (0.490–0.903) | 4.52 × 10−11 | 6.60 × 10−9 |
| SM | SM C24:1 | 0.518 (0.310–0.726) | 1.16 × 10−6 | 1.69 × 10−4 |
| Biogenic Amines | Alpha-Amino acid (alpha-AAA) | 0.955 (0.726–1.184) | 5.51 × 10−16 | 8.04 × 10−14 |
| Biogenic Amines | Kynurenine | 0.743 (0.524–0.962) | 3.81 × 10−11 | 5.57 × 10−9 |
| Biogenic Amines | 4-Hydroxyproline (t4-OH-Pro) | 0.485 (0.279–0.691) | 4.13 × 10−6 | 6.02 × 10−4 |
| Acylcarnitines | Carnitine (C0) | 0.672 (0.462 -0.882) | 4.20 × 10−10 | 6.13 × 10−8 |
| Acylcarnitines | Valerylcarnitine (C5) | 0.700 (0.478–0.922) | 7.96 × 10−10 | 1.16 × 10−7 |
| Acylcarnitines | Propionylcarnitine (C3) | 0.670 (0.449–0.891) | 3.50 × 10−9 | 5.11 × 10−7 |
| Acylcarnitines | Butyrylcarnitine (C4) | 0.457 (0.247–0.667) | 2.15 × 10−5 | 3.14 × 10−3 |
| PC ae | PC ae C36:5 | 0.502 (0.290–0.713) | 3.49 × 10−6 | 5.09 × 10−4 |
| PC ae | PC ae C36:4 | 0.457 (0.254–0.660) | 1.07 × 10−5 | 1.56 × 10−3 |
| PC ae | PC ae C32:2 | 0.506 (0.258–0.754) | 6.52 × 10−5 | 9.52 × 10−3 |
| Negatively Associated | ||||
| Category | Metabolite | Beta Estimate (95% CI) | p-value | q-value |
| lysoPC | lysoPC a C17:0 | −1.1 (−1.305–−0.896) | 2.88 × 10−25 | 4.20 × 10−23 |
| lysoPC | lysoPC a C18:2 | −1.129 (−1.348–−0.911) | 1.72 × 10−23 | 2.51 × 10−21 |
| lysoPC | lysoPC a C18:1 | −0.978 (−1.193–−0.763) | 1.08 × 10−18 | 8.72 × 10−15 |
| lysoPC | lysoPC a C16:0 | −0.640 (−0.849–−0.432) | 2.19 × 10−9 | 3.20 × 10−7 |
| lysoPC | lysoPC a C18:0 | −0.521 (−0.725–−0.316) | 6.48 × 10−7 | 9.46 × 10−5 |
| lysoPC | lysoPC a C20:4 | −0.415 (−0.627–−0.203) | 1.28 × 10−4 | 1.86 × 10−2 |
| Amino Acids | Asparagine (Asn) | −0.642 (−0.843–−0.44) | 5.30 × 10−10 | 7.73 × 10−8 |
| Amino Acids | Glycine (Gly) | −0.515 (−0.724–−0.305) | 1.60 × 10−6 | 2.34 × 10−4 |
| PC ae | PC ae C42:3 | −0.594 (−0.821–−0.368) | 2.94 × 10−7 | 4.29 × 10−5 |
| PC ae | PC ae C36:2 | −0.607 (−0.840–−0.373) | 3.75 × 10−7 | 5.48 × 10−5 |
| PC ae | PC ae C40:6 | −0.424 (−0.639–−0.209) | 1.14 × 10−4 | 1.66 × 10−2 |
| PC ae | PC ae C38:2 | −0.406 (−0.613–−0.199) | 1.23 × 10−4 | 1.80 × 10−2 |
| Category | Metabolite | Odds Ratios (95% CI) | p-Value | q-Value |
|---|---|---|---|---|
| Acylcarnitines | Hydroxybutyrylcarnitine (C3-DC (C4-OH)) | 0.619 (0.363–0.888) | 3.79 × 10−6 | 5.54 × 10−4 |
| Biogenic Amines | Alpha-Amino acid (alpha-AAA) | 0.638 (0.308–0.977) | 1.77 × 10−4 | 2.58 × 10−2 |
| Amino Acids | Isoleucine (Ile) | 0.637 (0.293–0.987) | 3.08 × 10−4 | 4.50 × 10−2 |
| Sobel Test (Metabolite, BMI, Fasting Glucose) | Sobel Test (Metabolite, BMI, HbA1c) | ||||
|---|---|---|---|---|---|
| Metabolite | p-Value | q-Value | Metabolite | p-Value | q-Value |
| Sum of hexoses (H1) | 1.49 × 10−16 | 2.18 × 10−14 | Sum of hexoses (H1) | 1.14 × 10−15 | 1.66 × 10−13 |
| SM C16:1 | 2.88 × 10−7 | 4.20 × 10−5 | Isoleucine (Ile) | 1.08 × 10−5 | 1.58 × 10−3 |
| Glutamate (Glu) | 1.27 × 10−6 | 1.85 × 10−4 | SM C16:1 | 1.40 × 10−5 | 2.04 × 10−3 |
| PC aa C38:3 | 2.62 × 10−6 | 3.82 × 10−4 | lysoPC a C18:0 | 5.56 × 10−5 | 8.11 × 10−3 |
| lysoPC a C17:0 | 1.31 × 10−5 | 1.91 × 10−3 | Leucine (Leu) | 1.05 × 10−4 | 1.53 × 10−2 |
| Alpha-Amino acid (alpha-AAA) | 1.58 × 10−5 | 2.3 × 10−3 | Glutamate (Glu) | 1.06 × 10−4 | 1.55 × 10−2 |
| Isoleucine (Ile) | 1.95 × 10−5 | 2.84 × 10−3 | lysoPC a C16:0 | 1.12 × 10−4 | 1.63 × 10−2 |
| lysoPC a C18:0 | 5.00 × 10−5 | 7.30 × 10−3 | Alpha-Amino acid (alpha-AAA) | 1.48 × 10−4 | 2.16 × 10−2 |
| Alanine (Ala) | 6.94 × 10−5 | 1.01 × 10−2 | PC aa C38:3 | 3.14 × 10−4 | 4.59 × 10−2 |
| SM C18:1 | 1.33 × 10−4 | 1.94 × 10−2 | |||
| Leucine (Leu) | 1.48 × 10−4 | 2.16 × 10−2 | |||
| SM C20:2 | 2.91 × 10−4 | 4.24 × 10−2 | |||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dong, Q.; Sidra, S.; Gieger, C.; Wang-Sattler, R.; Rathmann, W.; Prehn, C.; Adamski, J.; Koenig, W.; Peters, A.; Grallert, H.; et al. Metabolic Signatures Elucidate the Effect of Body Mass Index on Type 2 Diabetes. Metabolites 2023, 13, 227. https://doi.org/10.3390/metabo13020227
Dong Q, Sidra S, Gieger C, Wang-Sattler R, Rathmann W, Prehn C, Adamski J, Koenig W, Peters A, Grallert H, et al. Metabolic Signatures Elucidate the Effect of Body Mass Index on Type 2 Diabetes. Metabolites. 2023; 13(2):227. https://doi.org/10.3390/metabo13020227
Chicago/Turabian StyleDong, Qiuling, Sidra Sidra, Christian Gieger, Rui Wang-Sattler, Wolfgang Rathmann, Cornelia Prehn, Jerzy Adamski, Wolfgang Koenig, Annette Peters, Harald Grallert, and et al. 2023. "Metabolic Signatures Elucidate the Effect of Body Mass Index on Type 2 Diabetes" Metabolites 13, no. 2: 227. https://doi.org/10.3390/metabo13020227
APA StyleDong, Q., Sidra, S., Gieger, C., Wang-Sattler, R., Rathmann, W., Prehn, C., Adamski, J., Koenig, W., Peters, A., Grallert, H., & Sharma, S. (2023). Metabolic Signatures Elucidate the Effect of Body Mass Index on Type 2 Diabetes. Metabolites, 13(2), 227. https://doi.org/10.3390/metabo13020227