

Metabolomic Strategies to Improve Chemical Information from OSMAC Studies of Endophytic Fungi



Abstract
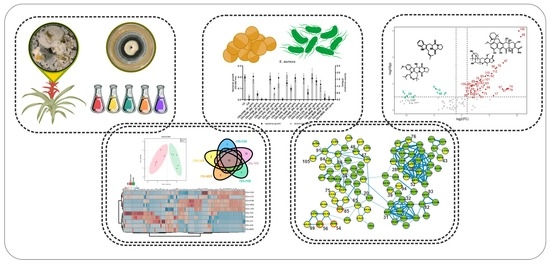
:
1. Introduction
2. Materials and Methods
2.1. Fungal Material and Identification of Strain
2.2. OSMAC-Based Cultivation of Fungi
2.3. Extraction from Broth and Mycelial Biomass
2.4. Chemical Characterization of Compounds by HPLC-DAD-MS/MS
2.5. Antibiofilm and Antibacterial Evaluations
2.6. Processing Data and Statistical Analyses
2.7. Molecular Networking and Data Analysis
2.8. Annotation of Compounds
3. Results
3.1. Identification of Fungal Strain
3.2. OSMAC Experiments from Penicillium sp. 5MP2F4
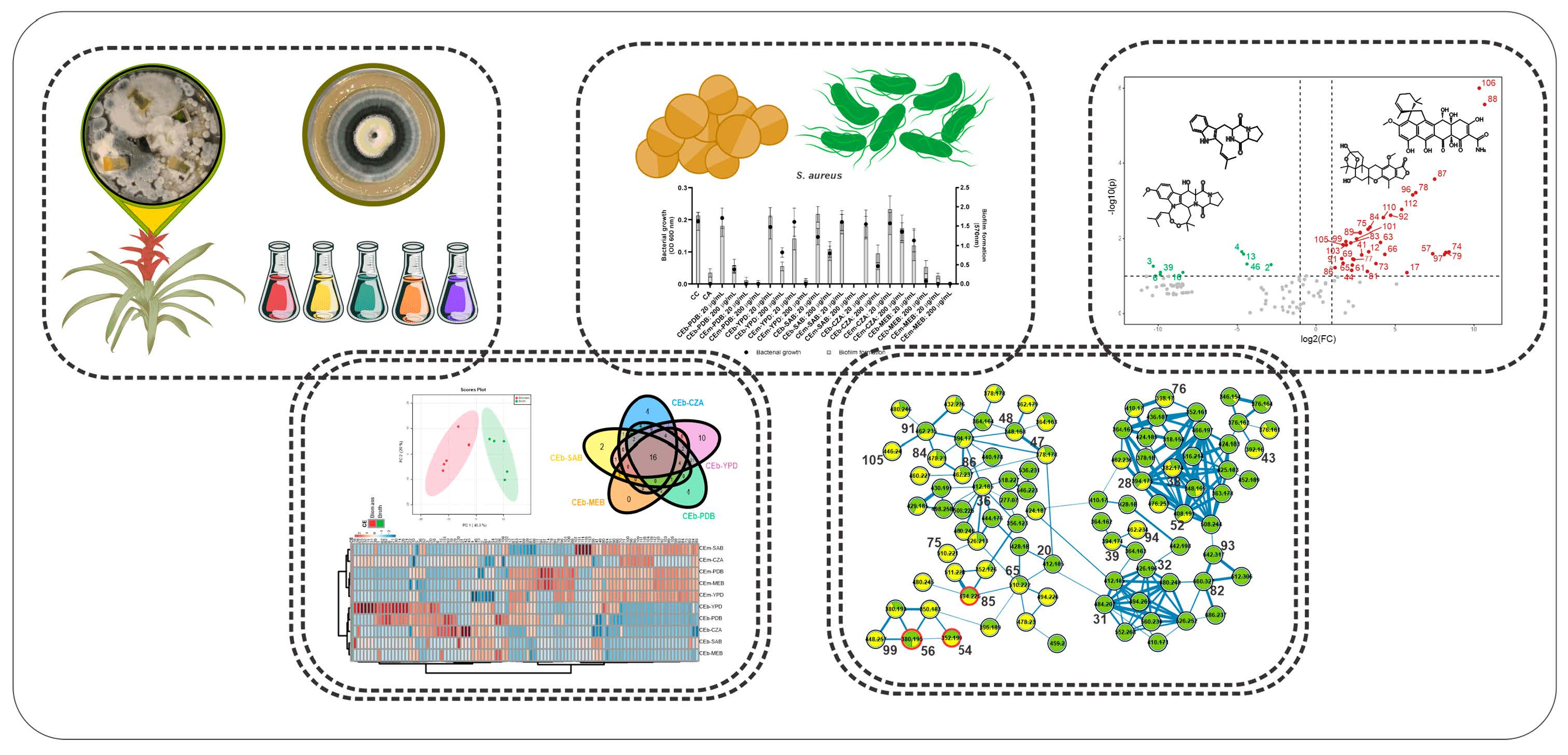
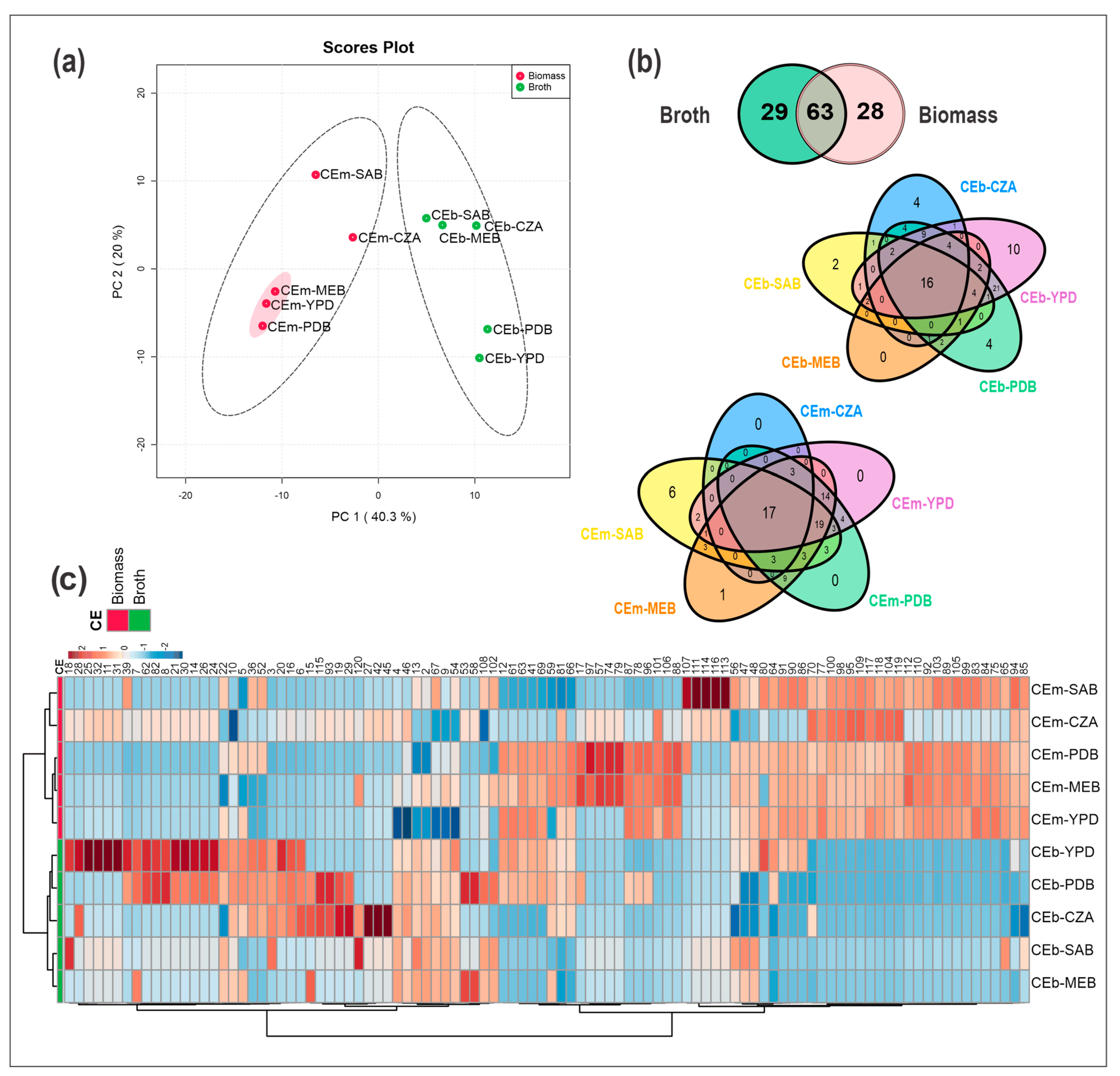
3.3. Chemical Diversity of the Crude Extracts of Broth (CEb) and Mycelial Biomass (CEm) from Penicillium sp. Strain 5MP2F4 Obtained by OSMAC Strategy
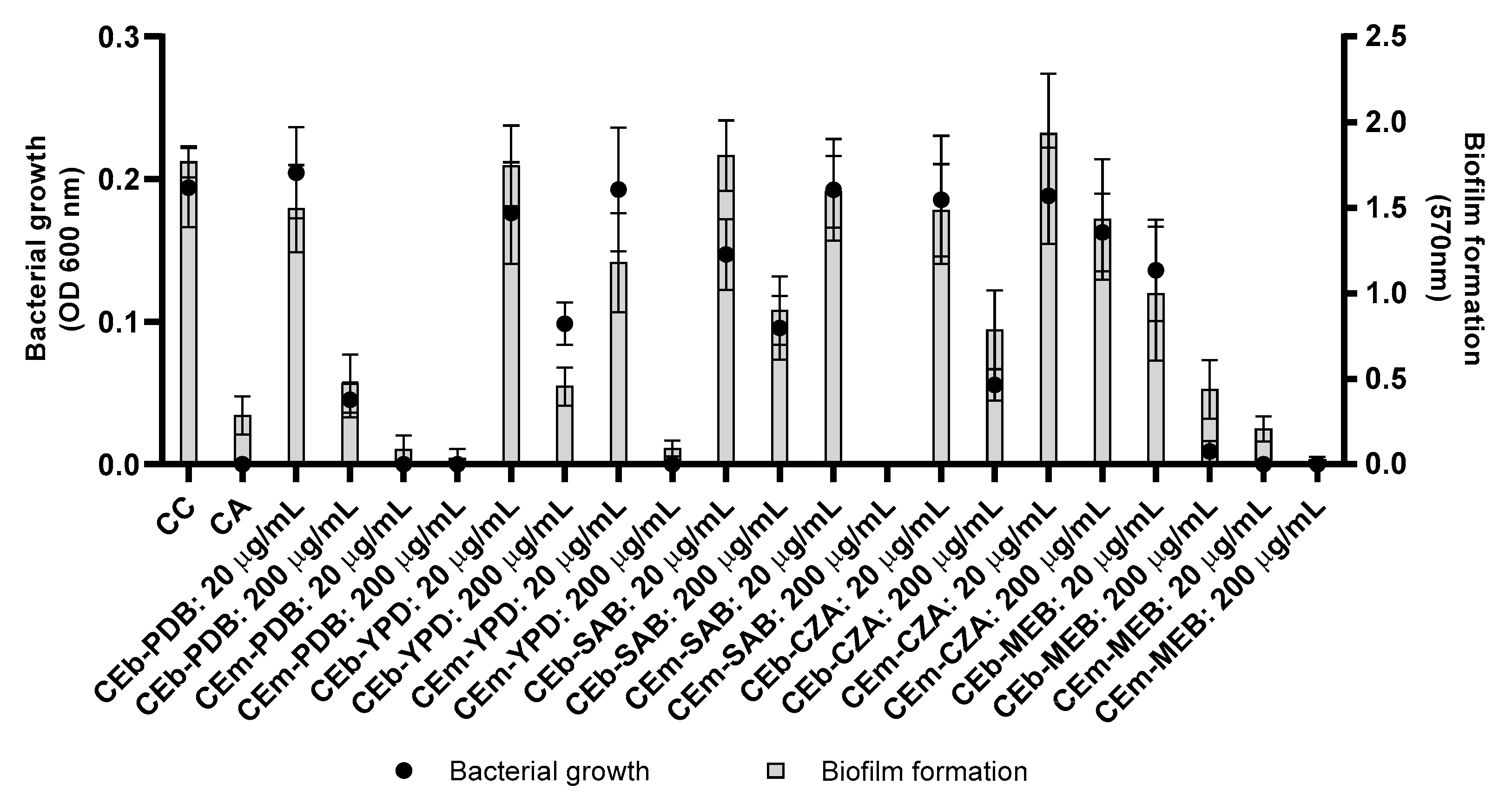
3.4. Evaluation of Antibacterial and Antibiofilm Activities of Crude Extracts of Broth (CEb) and Mycelial Biomass (CEm) from Penicillium sp. Strain 5MP2F4 Obtained by OSMAC Strategy
3.5. Metabolomics and Statistical Analyses Applied to Determine the Chemical Differences between Samples with High and Moderate Antibacterial Activity
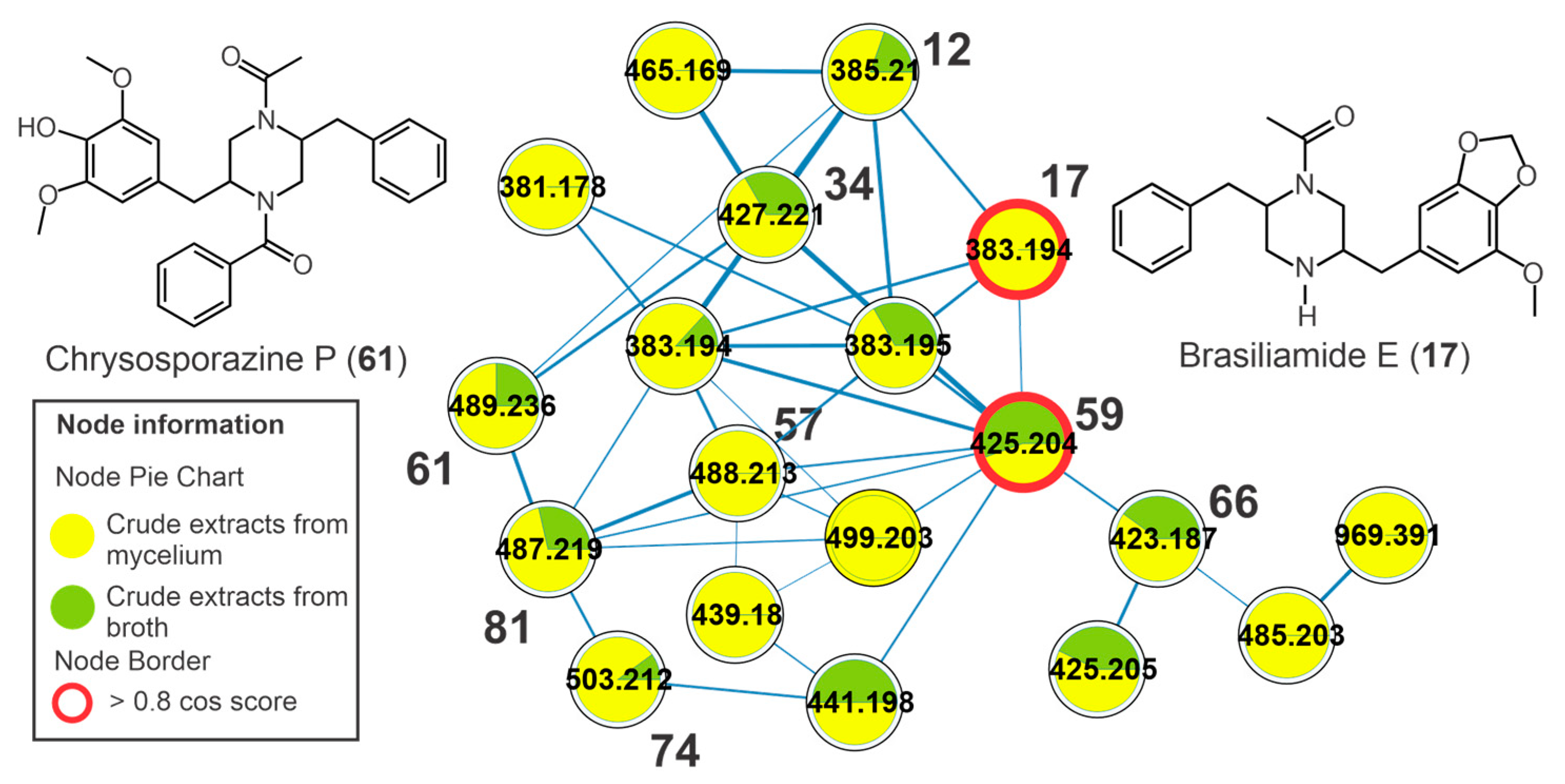
3.6. Annotation of Constituents from Crude Extracts of Broth (cEb) and Mycelial Biomass (cEm) from Penicillium sp. Strain 5MP2F4 and Molecular Networking
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Atanasov, A.G.; Zotchev, S.B.; Dirsch, V.M.; Orhan, I.E.; Banach, M.; Rollinger, J.M.; Barreca, D.; Weckwerth, W.; Bauer, R.; Bayer, E.A.; et al. Natural products in drug discovery: Advances and opportunities. Nat. Rev. Drug. Discov. 2021, 20, 200–216. [Google Scholar] [CrossRef] [PubMed]
- Gakuubi, M.M.; Munusamy, M.; Liang, Z.X.; Ng, S.B. Fungal endophytes: A promising frontier for discovery of novel bioactive compounds. J. Fungi 2021, 7, 786. [Google Scholar] [CrossRef]
- Mishra, R.; Venkateswara Sarma, V. Current perspectives of endophytic fungi in sustainable development. In Fungi and Their Role in Sustainable Development: Current Perspectives; Gehlot, P., Singh, J., Eds.; Springer: Singapore, 2018; pp. 553–584. ISBN 978-981-13-0393-7. [Google Scholar]
- Gomes, S.H.C.; Navegantes-Lima, P.W.P.; Reis, K.C.; Carvalho, J.D.E.; Konči´c, Z.K.; Tomczyk, M.; Helena, S.; Nogueira-Lima, C.; Wender, P.; Gomes, P.; et al. The roots of Deguelia Nitidula as a natural antibacterial source against Staphylococcus aureus strains. Metabolites 2022, 12, 1083. [Google Scholar] [CrossRef]
- Pham, J.V.; Yilma, M.A.; Feliz, A.; Majid, M.T.; Maffetone, N.; Walker, J.R.; Kim, E.; Cho, H.J.; Reynolds, J.M.; Song, M.C.; et al. A review of the microbial production of bioactive natural products and biologics. Front. Microbiol. 2019, 10, 1404. [Google Scholar] [CrossRef] [PubMed]
- Gupta, S.; Chaturvedi, P.; Kulkarni, M.G.; van Staden, J. A critical review on exploiting the pharmaceutical potential of plant endophytic fungi. Biotechnol. Adv. 2020, 39, 107462. [Google Scholar] [CrossRef] [PubMed]
- Romano, S.; Jackson, S.A.; Patry, S.; Dobson, A.D.W. Extending the “One Strain Many Compounds” (OSMAC) principle to marine microorganisms. Mar. Drugs 2018, 16, 244. [Google Scholar] [CrossRef]
- Bode, H.B.; Bethe, B.; Höfs, R.; Zeeck, A. Big effects from small changes: Possible ways to explore nature’s chemical diversity. ChemBioChem 2002, 3, 619–627. [Google Scholar] [CrossRef]
- Oprea, E.; Li, G.; Jian, T.; Liu, X.; Lv, Q.; Zhang, G.; Ling, J. Application of metabolomics in fungal research. Molecules 2022, 27, 7365. [Google Scholar] [CrossRef]
- Hamany Djande, C.Y.; Piater, L.A.; Steenkamp, P.A.; Tugizimana, F.; Dubery, I.A. A metabolomics approach and chemometric tools for differentiation of barley cultivars and biomarker discovery. Metabolites 2021, 11, 578. [Google Scholar] [CrossRef]
- Ye, D.; Li, X.; Shen, J.; Xia, X. Microbial metabolomics: From novel technologies to diversified applications. TrAC Trends Analyt. Chem. 2022, 148, 116540. [Google Scholar] [CrossRef]
- Belinato, J.R.; Bazioli, J.M.; Sussulini, A.; Augusto, F.; Filla, T.P. Metabolômica microbiana: Inovações e aplicações. Quim Nova 2019, 42, 546–559. [Google Scholar] [CrossRef]
- Cassemiro, N.S.; Sanches, L.B.; Kato, N.N.; Ruller, R.; Carollo, C.A.; de Mello, J.C.P.; dos Santos dos Anjos, E.; Silva, D.B. New derivatives of the iridoid specioside from fungal biotransformation. Appl. Microbiol. Biotechnol. 2021, 105, 7731–7741. [Google Scholar] [CrossRef]
- da Silva Trentin, D.; Giordani, R.B.; Zimmer, K.R.; da Silva, A.G.; da Silva, M.V.; dos Santos Correia, M.T.; Baumvol, I.J.R.; Macedo, A.J. Potential of medicinal plants from the brazilian semi-arid region (caatinga) against Staphylococcus epidermidis planktonic and biofilm lifestyles. J. Ethnopharmacol. 2011, 137, 327–335. [Google Scholar] [CrossRef]
- Zimmer, K.R.; Macedo, A.J.; Giordani, R.B.; Conceição, J.M.; Nicastro, G.G.; Boechat, A.L.; Baldini, R.L.; Abraham, W.R.; Termignoni, C. A steroidal molecule present in the egg wax of the tick rhipicephalus (boophilus) microplus inhibits bacterial biofilms. Environ. Microbiol. 2013, 15, 2008–2018. [Google Scholar] [CrossRef] [PubMed]
- Lommen, A. MetAlign: Interface-driven, versatile metabolomics tool for hyphenated full-scan mass spectrometry data preprocessing. Anal. Chem. 2009, 81, 3079–3086. [Google Scholar] [CrossRef]
- Tikunov, Y.M.; Laptenok, S.; Hall, R.D.; Bovy, A.; de Vos, R.C.H. MSClust: A tool for unsupervised mass spectra extraction of chromatography-mass spectrometry ion-wise aligned data. Metabolomics 2012, 8, 714–718. [Google Scholar] [CrossRef] [PubMed]
- Pang, Z.; Chong, J.; Zhou, G.; de Lima Morais, D.A.; Chang, L.; Barrette, M.; Gauthier, C.; Jacques, P.-É.; Li, S.; Xia, J. MetaboAnalyst 5.0: Narrowing the gap between raw spectra and functional insights. Nucleic Acids Res. 2021, 49, W388–W396. [Google Scholar] [CrossRef] [PubMed]
- Adusumilli, R.; Mallick, P. Data conversion with ProteoWizard MsConvert. Methods Mol. Biol. 2017, 1550, 339–368. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Carver, J.J.; Phelan, V.V.; Sanchez, L.M.; Garg, N.; Peng, Y.; Nguyen, D.D.; Watrous, J.; Kapono, C.A.; Luzzatto-Knaan, T.; et al. Sharing and community curation of mass spectrometry data with global natural products social molecular networking. Nat. Biotechnol. 2016, 34, 828–837. [Google Scholar] [CrossRef]
- Shannon, P.; Markiel, A.; Ozier, O.; Baliga, N.S.; Wang, J.T.; Ramage, D.; Amin, N.; Schwikowski, B.; Ideker, T. Cytoscape: A software environment for integrated models of biomolecular interaction networks. Genome Res. 2003, 13, 2498–2504. [Google Scholar] [CrossRef] [PubMed]
- Schoch, C.L.; Robbertse, B.; Robert, V.; Vu, D.; Cardinali, G.; Irinyi, L.; Meyer, W.; Nilsson, R.H.; Hughes, K.; Miller, A.N.; et al. Finding needles in haystacks: Linking scientific names, reference specimens and molecular data for fungi. Database 2014, 2014, 1–21. [Google Scholar] [CrossRef]
- Bazioli, J.M.; Amaral, L.D.S.; Fill, T.P.; Rodrigues-Filho, E. Insights into Penicillium brasilianum secondary metabolism and its biotechnological potential. Molecules 2017, 22, 858. [Google Scholar] [CrossRef] [PubMed]
- Filtenborg, O.; Frisvad, J.C.; Thrane, U. The significance of yeast extract composition on metabolite production in Penicillium. In Modern Concepts in Penicillium and Aspergillus Classification; Samson, R.A., Pitt, J.I., Eds.; Springer: Boston, MA, USA, 1990; pp. 433–441. ISBN 978-1-4899-3579-3. [Google Scholar]
- Demarque, D.P.; Dusi, R.G.; de Sousa, F.D.M.; Grossi, S.M.; Silvério, M.R.S.; Lopes, N.P.; Espindola, L.S. Mass spectrometry-based metabolomics approach in the isolation of bioactive natural products. Sci. Rep. 2020, 10, 1051. [Google Scholar] [CrossRef]
- Dewa, A.A.; Khalil, Z.G.; Elbanna, A.H.; Capon, R.J. Chrysosporazines revisited: Regioisomeric phenylpropanoid piperazine P-Glycoprotein inhibitors from Australian marine fish-derived fungi. Molecules 2022, 27, 3172. [Google Scholar] [CrossRef] [PubMed]
- Fill, T.P.; Pallini, H.F.; da Silva Amaral, L.; da Silva, J.V.; Bidóia, D.L.; Peron, F.; Garcia, F.P.; Nakamura, C.V.; Rodrigues-Filho, E. Copper and manganese cations alter secondary metabolism in the fungus Penicillium brasilianum. J. Braz. Chem. Soc. 2016, 27, 1444–1451. [Google Scholar] [CrossRef]
- Fornal, E.; Parfieniuk, E.; Czeczko, R.; Bilinska-Wielgus, N.; Frac, M. Fast and easy liquid chromatography–mass spectrometry method for evaluation of postharvest fruit safety by determination of mycotoxins: Fumitremorgin c and verruculogen. Postharvest Biol. Technol. 2017, 131, 46–54. [Google Scholar] [CrossRef]
- An, C.-Y.; Li, X.-M.; Li, C.-S.; Xu, G.-M.; Wang, B.-G. Prenylated indolediketopiperazine peroxides and related homologues from the marine sediment-derived fungus Penicillium brefeldianum SD-273. Mar. Drugs 2014, 12, 746–756. [Google Scholar] [CrossRef]
- Liu, L.; Ruan, H. An overview of natural austalides: Structure, bioactivity and synthesis. Phytochem. Lett. 2022, 47, 81–92. [Google Scholar] [CrossRef]
- Sversut, R.A.; Vieira, J.C.; Kassab, N.M.; Silva, D.B.; Salgado, H.R.N. Forced degradation behavior of two-drug combinations: Isolation and characterization of major degradation products by LC-MS. Microchem. J. 2019, 150, 104074. [Google Scholar] [CrossRef]
- de Lima, M.S.; Takahasi, A.; Damasceno-Junior, G.A.; Araujo, A.C. Checklist of the flora in ironstone outcrops at the urucum plateau, Corumbá, Mato Grosso Do Sul. Biota Neotrop. 2019, 19, 20180708. [Google Scholar] [CrossRef]
- Viana, T.F.C.; Campelo, A.P.S.; Baldani, J.I.; Fernandes-Júnior, P.I.; Baldani, V.L.D.; Silva, W.M.; Paggi, G.M.; Brasil, M.S. Cultivable bacterial diversity associated with bromeliad roots from ironstone outcrops in central Brazil. Braz. J. Biol. 2020, 80, 872–880. [Google Scholar] [CrossRef]
- Noriler, S.A.; Savi, D.C.; Ponomareva, L.V.; Rodrigues, R.; Rohr, J.; Thorson, J.S.; Glienke, C.; Shaaban, K.A. Vochysiamides A and B: Two new bioactive carboxamides produced by the new species Diaporthe vochysiae. Fitoterapia 2019, 138, 104273. [Google Scholar] [CrossRef] [PubMed]
- Noriler, S.A.; Savi, D.C.; Aluizio, R.; Palácio-Cortes, A.M.; Possiede, Y.M.; Glienke, C. Bioprospecting and structure of fungal endophyte communities found in the brazilian biomes, Pantanal, and Cerrado. Front. Microbiol. 2018, 9, 1526. [Google Scholar] [CrossRef] [PubMed]
- Iantas, J.; Savi, D.C.; Schibelbein, R.d.S.; Noriler, S.A.; Assad, B.M.; Dilarri, G.; Ferreira, H.; Rohr, J.; Thorson, J.S.; Shaaban, K.A.; et al. Endophytes of brazilian medicinal plants with activity against phytopathogens. Front. Microbiol. 2021, 12, 714750. [Google Scholar] [CrossRef] [PubMed]
- Savi, D.C.; Aluizio, R.; Glienke, C. Brazilian plants: An unexplored source of endophytes as producers of active metabolites. Planta Med. 2019, 85, 619–636. [Google Scholar] [CrossRef]
- Gubiani, J.R.; Oliveira, M.C.S.; Neponuceno, R.A.R.; Camargo, M.J.; Garcez, W.S.; Biz, A.R.; Soares, M.A.; Araujo, A.R.; Bolzani, V.d.S.; Lisboa, H.C.F.; et al. Cytotoxic prenylated indole alkaloid produced by the endophytic fungus Aspergillus terreus P63. Phytochem. Lett. 2019, 32, 162–167. [Google Scholar] [CrossRef]
- Pan, R.; Bai, X.; Chen, J.; Zhang, H.; Wang, H. Exploring structural diversity of microbe secondary metabolites using OSMAC strategy: A literature review. Front. Microbiol. 2019, 10, 294. [Google Scholar] [CrossRef]
- Hoang, T.P.T.; Roullier, C.; Boumard, M.C.; Robiou Du Pont, T.; Nazih, H.; Gallard, J.F.; Pouchus, Y.F.; Beniddir, M.A.; Grovel, O. Metabolomics-driven discovery of meroterpenoids from a mussel-derived Penicillium ubiquetum. J. Nat. Prod. 2018, 81, 2501–2511. [Google Scholar] [CrossRef]
- Xie, C.L.; Liu, Q.; He, Z.H.; Gai, Y.B.; Zou, Z.B.; Shao, Z.Z.; Liu, G.M.; Chen, H.F.; Yang, X.W. Discovery of andrastones from the deep-sea-derived Penicillium allii-sativi MCCC 3A00580 by OSMAC strategy. Bioorg. Chem. 2021, 108, 104671. [Google Scholar] [CrossRef] [PubMed]
- Meng, L.H.; Li, X.M.; Liu, Y.; Xu, G.M.; Wang, B.G. Antimicrobial alkaloids produced by the mangrove endophyte Penicillium brocae MA-231 using the OSMAC approach. RSC Adv. 2017, 7, 55026–55033. [Google Scholar] [CrossRef] [Green Version]
- Morais, L.S.; Dusi, R.G.; Demarque, D.P.; Silva, R.L.; Albernaz, L.C.; Bao, S.N.; Merten, C.; Antinarelli, L.M.R.; Coimbra, E.S.; Espindola, L.S. Antileishmanial compounds from Connarus suberosus: Metabolomics, isolation and mechanism of action. PLoS ONE 2020, 15, e0241855. [Google Scholar] [CrossRef] [PubMed]
- de Oliveira Dembogurski, D.S.; Silva Trentin, D.; Boaretto, A.G.; Rigo, G.V.; da Silva, R.C.; Tasca, T.; Macedo, A.J.; Carollo, C.A.; Silva, D.B. Brown propolis-metabolomic innovative approach to determine compounds capable of killing Staphylococcus aureus biofilm and Trichomonas vaginalis. Food Res. Int. 2018, 111, 661–673. [Google Scholar] [CrossRef]
- Deutsch, J.M.; Mandelare-Ruiz, P.; Yang, Y.; Foster, G.; Routhu, A.; Houk, J.; de La Flor, Y.T.; Ushijima, B.; Meyer, J.L.; Paul, V.J.; et al. Metabolomics approaches to dereplicate natural products from coral-derived bioactive bacteria. J. Nat. Prod. 2022, 85, 462–478. [Google Scholar] [CrossRef]
- Minarini, L.A.D.R.; de Andrade, L.N.; de Gregorio, E.; Grosso, F.; Naas, T.; Zarrilli, R.; Camargo, I.L.B.C. Antimicrobial resistance as a global public health problem: How can we address it? Front. Public Health 2020, 8, 768. [Google Scholar] [CrossRef] [PubMed]
- Singh, A.; Singh, D.K.; Kharwar, R.N.; White, J.F.; Gond, S.K.; Singh, A.; Singh, D.K.; Kharwar, R.N.; White, J.F.; Gond, S.K. Fungal endophytes as efficient sources of plant-derived bioactive compounds and their prospective applications in natural product drug discovery: Insights, avenues, and challenges. Microorganisms 2021, 9, 197. [Google Scholar] [CrossRef]
- Radić, N.; Štrukelj, B. Endophytic fungi—The treasure chest of antibacterial substances. Phytomedicine 2012, 19, 1270–1284. [Google Scholar] [CrossRef]
- Song, Z.; Hou, Y.; Yang, Q.; Li, X.; Wu, S. Structures and biological activities of diketopiperazines from marine organisms: A Review. Mar. Drugs 2021, 19, 403. [Google Scholar] [CrossRef]
- Yuan, B.; Liu, D.; Guan, X.; Yan, Y.; Zhang, J.; Zhang, Y.; Yang, D.; Ma, M.; Lin, W. Piperazine ring formation by a single-module NRPS and cleavage by an α-KG-Dependent nonheme iron dioxygenase in brasiliamide biosynthesis. Appl. Microbiol. Biotechnol. 2020, 104, 6149–6159. [Google Scholar] [CrossRef]
- Fujita, T.; Hayashi, H. New brasiliamide congeners, brasiliamides C, D and E, from Penicillium brasilianum Batista JV-379. Biosci. Biotechnol. Biochem. 2004, 68, 820–826. [Google Scholar] [CrossRef]
- Fujita, T.; Makishima, D.; Akiyama, K.; Hayashi, H. New convulsive compounds, brasiliamides A and B, from Penicillium brasilianum Batista JV-379. Biosci. Biotechnol. Biochem. 2002, 66, 1697–1705. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fill, T.P.; da Silva, B.F.; Rodrigues-Fo, E. Biosynthesis of phenylpropanoid amides by an endophytic Penicillium brasilianum found in root bark of Melia azedarach. J. Microbiol. Biotechnol. 2009, 20, 622–629. [Google Scholar] [CrossRef]
- Ma, Y.-M.; Liang, X.-A.; Kong, Y.; Jia, B. Structural diversity and biological activities of indole diketopiperazine alkaloids from fungi. J. Agric. Food Chem. 2016, 64, 6659–6671. [Google Scholar] [CrossRef]
- Zhang, H.-C.; Ma, Y.-M.; Liu, R.; Zhou, F. Endophytic fungus Aspergillus tamarii from Ficus carica L., a new source of indolyl diketopiperazines. Biochem. Syst. Ecol. 2012, 45, 31–33. [Google Scholar] [CrossRef]
- Zhang, Y.H.; Geng, C.; Zhang, X.W.; Zhu, H.J.; Shao, C.L.; Cao, F.; Wang, C.Y. Discovery of bioactive indole-diketopiperazines from the marine-derived fungus Penicillium brasilianum aided by genomic information. Mar. Drugs 2019, 17, 514. [Google Scholar] [CrossRef] [PubMed]
- Kato, N.; Suzuki, H.; Takagi, H.; Asami, Y.; Kakeya, H.; Uramoto, M.; Usui, T.; Takahashi, S.; Sugimoto, Y.; Osada, H. Identification of cytochrome P450s required for fumitremorgin biosynthesis in Aspergillus fumigatus. ChemBioChem 2009, 10, 920–928. [Google Scholar] [CrossRef]
- Knaus, H.G.; McManus, O.B.; Lee, S.H.; Schmalhofer, W.A.; Garcia-Calvo, M.; Helms, L.M.; Sanchez, M.; Giangiacomo, K.; Reuben, J.P.; Smith, A.B., 3rd. Tremorgenic indole alkaloids potently inhibit smooth muscle high-conductance calcium-activated potassium channels. Biochemistry 1994, 33, 5819–5828. [Google Scholar] [CrossRef]
- Zhou, Y.; Debbab, A.; Wray, V.; Lin, W.; Schulz, B.; Trepos, R.; Pile, C.; Hellio, C.; Proksch, P.; Aly, A.H. Marine bacterial inhibitors from the sponge-derived fungus Aspergillus sp. Tetrahedron. Lett. 2014, 55, 2789–2792. [Google Scholar] [CrossRef]
- Shang, Z.; Salim, A.A.; Khalil, Z.; Quezada, M.; Bernhardt, P.V.; Capon, R.J. Viridicatumtoxins: Expanding on a rare tetracycline antibiotic scaffold. J. Org. Chem. 2015, 80, 12501–12508. [Google Scholar] [CrossRef]
- Li, W.; Li, L.; Zhang, C.; Cai, Y.; Gao, Q.; Wang, F.; Cao, Y.; Lin, J.; Li, J.; Shang, Z.; et al. Investigations into the antibacterial mechanism of action of viridicatumtoxins. ACS Infect. Dis. 2020, 6, 1759–1769. [Google Scholar] [CrossRef] [PubMed]
- Zheng, C.J.; Yu, H.E.; Kim, E.H.; Kim, W.G. Viridicatumtoxin B, a new anti-MRSA agent from Penicillium sp. FR11. J. Antibiot. 2008, 61, 633–637. [Google Scholar] [CrossRef]
- Lebeau, J.; Venkatachalam, M.; Fouillaud, M.; Petit, T.; Vinale, F.; Dufossé, L.; Caro, Y. Production and new extraction method of polyketide red pigments produced by ascomycetous fungi from terrestrial and marine habitats. J. Fungi 2017, 3, 34. [Google Scholar] [CrossRef] [PubMed]
- Chooi, Y.H.; Cacho, R.; Tang, Y. Identification of the viridicatumtoxin and griseofulvin gene clusters from Penicillium aethiopicum. Chem. Biol. 2010, 17, 483–494. [Google Scholar] [CrossRef] [PubMed]
- Nicolaou, K.C.; Hale, C.R.H.; Nilewski, C.; Ioannidou, H.A.; Elmarrouni, A.; Nilewski, L.G.; Beabout, K.; Wang, T.T.; Shamoo, Y. Total Synthesis of Viridicatumtoxin B and Analogues Thereof: Strategy Evolution, Structural Revision, and Biological Evaluation. J. Am. Chem. Soc. 2014, 136, 12137–12160. [Google Scholar] [CrossRef]
- Newman, D.J. Old and modern antibiotic structures with potential for today’s infections. ADMET DMPK 2022, 10, 131. [Google Scholar] [CrossRef] [PubMed]
- Han, J.; Liu, M.; Jenkins, I.D.; Liu, X.; Zhang, L.; Quinn, R.J.; Feng, Y. Genome-inspired chemical exploration of marine fungus Aspergillus fumigatus MF071. Mar. Drugs 2020, 18, 352. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Peak | RT (min) | Compound | MF ** | Positive Mode (m/z) |
|---|---|---|---|---|
| [M+H]+ | ||||
| 2 | 3.89 | 5,6-Dihydro-6-hydroxypenicillic acid | C8H12O5 | 189.0761 |
| 3 | 11.17 | Unknown | C11H16O7S | 275.0582 # |
| 4 | 11.68 | Penicillic acid * | C8H10O4 | 171.0655 |
| 6 | 12.90 | Orsellinic acid | C8H8O4 | 151.0384 # |
| 8 | 15.36 | Dihydroxypropyl dihydroxy-methylbenzoate | C11H14O6 | 243.0866 |
| 10 | 16.33 | Unknown | C12H18O7 | 275.1124 |
| 11 | 16.64 | Unknown | C10H11NO3 | 194.0813 |
| 12 | 17.16 | Brasiliamide I | C22H28N2O4 | 385.2123 |
| 17 | 21.74 | Brasiliamide E | C22H26N2O4 | 383.1958 |
| 20 | 23.78 | Dihydroxyfumitremorgin C or Cyclotryprostatin A | C22H25N3O5 | 412.1863 |
| 21 | 24.64 | Unknown | C22H25N3O7 | 444.176 |
| 22 | 25.67 | Dihydroxyfumitremorgin C or Cyclotryprostatin A | C22H25N3O5 | 412.1855 |
| 23 | 26.21 | Triacetylfusarinin C | C39H60N6O15 | 853.4161 |
| 24 | 26.24 | Unknown | C24H25N7O3 | 410.1701 F |
| 25 | 26.82 | Unknown | C22H25N3O7 | 426.1664 # |
| 26 | 26.99 | Dihydroxy-spirotryprostatin A | C22H25N3O6 | 410.1705 # |
| 28 | 27.11 | Spirotryprostatin G | C23H29N3O6 | 394.1765 |
| 30 | 27.22 | Cyclotryprostatin E | C23H29N3O6 | 412.1869 M |
| 31 | 27.36 | Unknown | C25H29N3O7 | 484.2073 |
| 32 | 28.58 | Unknown | C22H23N3O6 | 426.1963 |
| 33 | 28.66 | Unknown | C22H23N3O6 | 410.1705 # |
| 34 | 28.76 | N-acetyl brasiliamide I | C24H30N2O5 | 427.2212 |
| 36 | 29.10 | Dihydroxyfumitremorgin C/Cyclotryprostatin A | C22H25N3O5 | 412.1861 |
| 37 | 29.38 | Dihydroxyfumitremorgin C/Cyclotryprostatin A | C22H25N3O5 | 394.1752 # |
| 38 | 29.76 | Cyclotryprostatin C /hydroxycycloprostatin A | C21H23N3O4 | 382.1755 |
| 39 | 30.08 | Cyclotryprostatin B | C23H27N3O5 | 394.1756 |
| 40 | 30.14 | Cyclotryprostatin C | C21H21N3O3 | 364.1561 |
| 41 | 30.16 | Unknown | C30H34N6O4 | 555.2699 |
| 43 | 30.62 | Oxo-cyclotryprostatin A | C22H23N3O5 | 392.1600 # |
| 45 | 30.66 | Abscisic acid | C15H20O4 | 265.1439 |
| 44 | 30.66 | Austalide H acid | C25H34O8 | 463.231 |
| 47 | 30.97 | Unknown | C22H25N3O4 | 378.179 # |
| 48 | 31.01 | Dehydroxycyclotryprostatin C | C21H23N3O3 | 348.1704 # |
| 50 | 31.23 | 17-O-desmethyl austalide B * | C25H32O8 | 461.2156 |
| 52 | 31.32 | Unknown | C23H25N3O4 | 408.1893 |
| 53 | 31.46 | Unknown | C22H14N4O8 | 463.087 |
| 54 | 31.49 | Tryprostatin B | C21H25N3O2 | 352.2019 |
| 55 | 31.74 | Cyclodepsipeptide (JBIR-113) | C31H41N5O7 | 596.3086 |
| 56 | 31.89 | Fumitremorgin C | C22H25N3O3 | 380.1947 |
| 57 | 32.08 | Unknown | C28H29N3O5 | 488.2171 |
| 58 | 32.13 | Unknown | C21H18O12 | 431.0599 M |
| 59 | 32.14 | Brasiliamide D | C24H28N2O5 | 425.2058 |
| 60 | 32.37 | Dihydroxyfumitremorgin C/Cyclotryprostatin A | C22H25N3O5 | 412.1854 |
| 61 | 32.43 | Chrysosporazine P | C29H32N2O5 | 489.2372 |
| 63 | 32.48 | Austalide Q acid | C25H32O7 | 445.2216 |
| 64 | 32.69 | Tryprostatin B isomer | C21H25N3O2 | 352.2014 |
| 65 | 32.82 | Oxoverrucologen (isomer) | C27H31N3O7 | 510.222 |
| 66 | 32.90 | Brasiliamide C | C24H26N2O5 | 423.1899 |
| 68 | 33.08 | Brasiliamide A | C24H26N2O6 | 439.1861 |
| 69 | 33.19 | Austalide H | C26H36O8 | 477.2460 |
| 70 | 33.29 | Unknown | C33H36N2O3 | 509.2747 |
| 71 | 33.34 | Oxoverruculogen | C27H31N3O7 | 510.2316 |
| 72 | 33.50 | Brasiliamide B | C24H26N2O5 | 423.1865 |
| 73 | 33.58 | 13-O-deacetyl austalide I/austalide J | C25H32O7 | 445.2198 |
| 74 | 33.91 | Unknown | C29H30N2O6 | 503.2177 |
| 75 | 34.07 | Oxoverruculogen (isomer) | C27H31N3O7 | 510.2231 |
| 76 | 34.45 | Indole alkaloid—diketopiperazine | C21H23N3O5 | 398.1712 |
| 78 | 34.61 | Austalide P acid | C25H34O7 | 447.2362 |
| 80 | 34.68 | Oxo-Hydroxyfumitremorgin C | C22H23N3O5 | 410.1712 |
| 81 | 34.74 | Chrysosporazine D | C29H30N2O5 | 487.2217 |
| 82 | 34.84 | Unknown | C37H46N3O8 | 660.329 |
| 83 | 35.09 | Hydroxy-verruculogen | C27H33N3O8 | 510.2213 # |
| 84 | 35.09 | Oxofumitremorgin B | C27H31N3O5 | 478.2324 |
| 85 | 35.39 | Verruculogen | C27H33N3O7 | 494.2282 # |
| 86 | 35.59 | Fumitrermorgin B (isomer) | C27H33N3O5 | 462.2375 # |
| 87 | 35.82 | Austalide L/Austalide W | C25H32O6 | 429.2259 |
| 88 | 35.94 | Helvamide | C29H28N2O5 | 485.2076 |
| 89 | 36.21 | Deoxy-fumitremorgin B | C27H33N3O4 | 446.2426 # |
| 90 | 36.35 | Hydroxyfumitremorgin B | C27H33N3O6 | 478.2325 # |
| 91 | 36.41 | Fumitrermorgin B (isomer) | C27H33N3O5 | 462.2375 # |
| 92 | 36.55 | Viridicatumtoxin A * | C30H31NO10 | 548.1909 # |
| 93 | 36.55 | Indole alkaloid | C37H43N3O7 | 642.3179 |
| 94 | 36.58 | Fumitrermorgin B | C27H33N3O5 | 462.2389 # |
| 95 | 36.68 | Linoleoyl-sn-glycero-3-phosphoethanolamine (isomer) | C23H44NO7P | 478.2869 |
| 96 | 36.78 | Austalide P | C26H36O7 | 443.2418 # |
| 97 | 36.83 | Viridicatum toxin A (isomer) | C30H31NO10 | 548.1909 # |
| 98 | 37.02 | Linoleoyl-sn-glycero-3-phosphoethanolamine (isomer) | C23H44NO7P | 478.29 |
| 99 | 37.05 | Di-deoxy-fumitremorgin B | C27H33N3O3 | 448.2574 |
| 100 | 37.26 | Linoleoyl-sn-glycero-3-phosphocholine | C26H50NO7P | 520.3381 |
| 101 | 37.33 | Dihydroaustalide K | C25H34O5 | 415.2484 |
| 102 | 37.43 | GameXPeptide C | C28H57NO9 | 552.4114 |
| 104 | 37.75 | Palmitoyl-sn-glycerophosphatidylethanolamine | C21H44NO7P | 454.2941 |
| 105 | 37.93 | Indole alkaloid—diketopiperazine | C27H31N3O3 | 446.2434 |
| 106 | 38.04 | Austalide K | C25H32O5 | 413.24 |
| 107 | 38.07 | Unknown | C27H52N3O3P | 498.3781 |
| 108 | 38.13 | GameXPeptide D | C30H55N5O5 | 566.4266 |
| 109 | 38.29 | Lysophosphatidylethanolamine (18:1) | C23H47NO7P | 480.3082 |
| 110 | 38.40 | Spirohexaline | C31H32O10 | 547.1949 # |
| 111 | 38.46 | LDGTS 18:2 | C28H52NO6 | 498.3711 |
| 112 | 38.73 | Previridicatumtoxin | C30H33NO10 | 550.2062 # |
| 113 | 39.36 | LDGTS 16:0 | C26H52NO6 | 474.375 |
| 116 | 39.86 | LDGTS 18:1 | C28H54NO6 | 500.3881 |
| 119 | 42.29 | Octadecadienoic acid | C18H32O2 | 303.2282 N |
| 120 | 42.30 | DGTS 18:2-18:2 | C46H81NO7 | 760.6068 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
da Silva, F.M.R.; Paggi, G.M.; Brust, F.R.; Macedo, A.J.; Silva, D.B. Metabolomic Strategies to Improve Chemical Information from OSMAC Studies of Endophytic Fungi. Metabolites 2023, 13, 236. https://doi.org/10.3390/metabo13020236
da Silva FMR, Paggi GM, Brust FR, Macedo AJ, Silva DB. Metabolomic Strategies to Improve Chemical Information from OSMAC Studies of Endophytic Fungi. Metabolites. 2023; 13(2):236. https://doi.org/10.3390/metabo13020236
Chicago/Turabian Styleda Silva, Fernanda Motta Ribeiro, Gecele Matos Paggi, Flávia Roberta Brust, Alexandre José Macedo, and Denise Brentan Silva. 2023. "Metabolomic Strategies to Improve Chemical Information from OSMAC Studies of Endophytic Fungi" Metabolites 13, no. 2: 236. https://doi.org/10.3390/metabo13020236
APA Styleda Silva, F. M. R., Paggi, G. M., Brust, F. R., Macedo, A. J., & Silva, D. B. (2023). Metabolomic Strategies to Improve Chemical Information from OSMAC Studies of Endophytic Fungi. Metabolites, 13(2), 236. https://doi.org/10.3390/metabo13020236