

Metabolomics in Corneal Diseases: A Narrative Review from Clinical Aspects


Abstract
:1. Introduction
2. Principles of Metabolomic Experiments
3. Metabolism and Metabolomic Profiles in Corneas
4. Metabolomic Studies in Corneal Diseases
Dry Eye Disease
5. Sjogren’s Syndrome
6. Contact Lens Wearing
7. Refractive Surgery
8. Keratoconus
9. Diabetic Corneas
10. Conclusions and Future Direction
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sun, Y.V.; Hu, Y.J. Integrative Analysis of Multi-omics Data for Discovery and Functional Studies of Complex Human Diseases. Adv. Genet. 2016, 93, 147–190. [Google Scholar]
- Trifonova, O.; Lokhov, P.; Archakov, A. Postgenomics diagnostics: Metabolomics approaches to human blood profiling. OMICS 2013, 17, 550–559. [Google Scholar] [CrossRef] [PubMed]
- Rinschen, M.M.; Ivanisevic, J.; Giera, M.; Siuzdak, G. Identification of bioactive metabolites using activity metabolomics. Nat. Rev. Mol. Cell Biol. 2019, 20, 353–367. [Google Scholar] [CrossRef]
- Chen, L.; Cheng, C.-Y.; Choi, H.; Ikram, M.K.; Sabanayagam, C.; Tan, G.S.W.; Tian, D.; Zhang, L.; Venkatesan, G.; Tai, E.S.; et al. Plasma Metabonomic Profiling of Diabetic Retinopathy. Diabetes 2016, 65, 1099–1108. [Google Scholar] [CrossRef] [Green Version]
- Lains, I.; Mendez, K.; Nigalye, A.; Katz, R.; Douglas, V.P.; Kelly, R.S.; Kim, I.K.; Miller, J.B.; Vavvas, D.G.; Liang, L.; et al. Plasma Metabolomic Profiles Associated with Three-Year Progression of Age-Related Macular Degeneration. Metabolites 2022, 12, 32. [Google Scholar] [CrossRef]
- Barbosa Breda, J.; Croitor Sava, A.; Himmelreich, U.; Somers, A.; Matthys, C.; Rocha Sousa, A.; Vandewalle, E.; Stalmans, I. Metabolomic profiling of aqueous humor from glaucoma patients—The metabolomics in surgical ophthalmological patients (MISO) study. Exp. Eye Res. 2020, 201, 108268. [Google Scholar] [CrossRef]
- Xu, J.; Su, G.; Huang, X.; Chang, R.; Chen, Z.; Ye, Z.; Cao, Q.; Kijlstra, A.; Yang, P. Metabolomic Analysis of Aqueous Humor Identifies Aberrant Amino Acid and Fatty Acid Metabolism in Vogt-Koyanagi-Harada and Behcet’s Disease. Front. Immunol. 2021, 12, 587393. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Hou, X.W.; Liang, G.; Pan, C.W. Metabolomics in Glaucoma: A Systematic Review. Investig. Ophthalmol. Vis. Sci. 2021, 62, 9. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Gao, Y.; Wang, L.Z.; Cheung, N.; Tan, G.S.; Cheung, G.C.M.; Beuerman, R.W.; Wong, T.Y.; Chan, E.C.Y.; Zhou, L. Recent advances in the applications of metabolomics in eye research. Anal. Chim. Acta 2018, 1037, 28–40. [Google Scholar] [CrossRef]
- Chen, X.; Zhang, C.; Tian, L.; Wu, L.; Jie, Y.; Wang, N.; Liu, R.; Wang, L. In situ metabolic profile and spatial distribution of ocular tissues: New insights into dry eye disease. Ocul. Surf. 2022, 24, 51–63. [Google Scholar] [CrossRef]
- Daphne Teh, A.L.; Jayapalan, J.J.; Loke, M.F.; Wan Abdul Kadir, A.J.; Subrayan, V. Identification of potential serum metabolic biomarkers for patient with keratoconus using untargeted metabolomics approach. Exp. Eye Res. 2021, 211, 108734. [Google Scholar] [CrossRef]
- Ju, U.H.; Liu, F.C.; Lin, C.S.; Huang, W.Y.; Lin, T.Y.; Shen, C.H.; Chou, Y.C.; Lin, C.L.; Lin, K.T.; Kao, C.H.; et al. Risk of Parkinson disease in Sjögren syndrome administered ineffective immunosuppressant therapies: A nationwide population-based study. Medicine 2019, 98, e14984. [Google Scholar] [CrossRef]
- Li, M.; Liu, L.; Shi, Y.; Sun, L.; Ma, X.; Zou, J. Age-related differences in corneal nerve regeneration after SMILE and the mechanism revealed by metabolomics. Exp. Eye Res. 2021, 209, 108665. [Google Scholar] [CrossRef] [PubMed]
- Willcox, M.D. Tear film, contact lenses and tear biomarkers. Clin. Exp. Optom. 2019, 102, 350–363. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Padberg, I.; Peter, E.; González-Maldonado, S.; Witt, H.; Mueller, M.; Weis, T.; Bethan, B.; Liebenberg, V.; Wiemer, J.; Katus, H.A.; et al. A new metabolomic signature in type-2 diabetes mellitus and its pathophysiology. PLoS ONE 2014, 9, e85082. [Google Scholar] [CrossRef] [Green Version]
- Tan, S.Z.; Begley, P.; Mullard, G.; Hollywood, K.A.; Bishop, P.N. Introduction to metabolomics and its applications in ophthalmology. Eye 2016, 30, 773–783. [Google Scholar] [CrossRef] [Green Version]
- Schmidt, D.R.; Patel, R.; Kirsch, D.G.; Lewis, C.A.; Vander Heiden, M.G.; Locasale, J.W. Metabolomics in cancer research and emerging applications in clinical oncology. CA Cancer J. Clin. 2021, 71, 333–358. [Google Scholar] [CrossRef] [PubMed]
- Wishart, D.S.; Guo, A.; Oler, E.; Wang, F.; Anjum, A.; Peters, H.; Dizon, R.; Sayeeda, Z.; Tian, S.; Lee, B.L.; et al. HMDB 5.0: The Human Metabolome Database for 2022. Nucleic Acids Res. 2022, 50, D622–D631. [Google Scholar] [CrossRef]
- Bothwell, J.H.; Griffin, J.L. An introduction to biological nuclear magnetic resonance spectroscopy. Biol. Rev. Camb. Philos. Soc. 2011, 86, 493–510. [Google Scholar] [CrossRef]
- Emwas, A.H. The strengths and weaknesses of NMR spectroscopy and mass spectrometry with particular focus on metabolomics research. Methods Mol. Biol. 2015, 1277, 161–193. [Google Scholar]
- Amberg, A.; Riefke, B.; Schlotterbeck, G.; Ross, A.; Senn, H.; Dieterle, F.; Keck, M. NMR and MS Methods for Metabolomics. Methods Mol. Biol. 2017, 1641, 229–258. [Google Scholar] [PubMed]
- Rubakhin, S.S.; Sweedler, J.V. A mass spectrometry primer for mass spectrometry imaging. Methods Mol. Biol. 2010, 656, 21–49. [Google Scholar]
- Pitt, J.J. Principles and applications of liquid chromatography-mass spectrometry in clinical biochemistry. Clin. Biochem. Rev. 2009, 30, 19–34. [Google Scholar]
- Gika, H.G.; Theodoridis, G.A.; Plumb, R.S.; Wilson, I.D. Current practice of liquid chromatography-mass spectrometry in metabolomics and metabonomics. J. Pharm. Biomed. Anal. 2014, 87, 12–25. [Google Scholar] [CrossRef] [PubMed]
- Fatt, I.; Bieber, M.T.; Pye, S.D. Steady state distribution of oxygen and carbon dioxide in the in vivo cornea of an eye covered by a gas-permeable contact lens. Am. J. Optom. Arch. Am. Acad. Optom. 1969, 46, 3–14. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Li, H.; Ogando, D.G.; Li, S.; Feng, M.; Price, F.W.; Tennessen, J.M.; Bonanno, J.A. Glutaminolysis is Essential for Energy Production and Ion Transport in Human Corneal Endothelium. EBioMedicine 2017, 16, 292–301. [Google Scholar] [CrossRef] [Green Version]
- Fatt, I.; Weissman, B.A. 7—Cornea II: Metabolism, Oxygen, Carbon Dioxide, and Contact Lens Wear. In Physiology of the Eye, 2nd ed.; Fatt, I., Weissman, B.A., Eds.; Butterworth-Heinemann: Oxford, UK, 1992; pp. 151–204. [Google Scholar]
- Geroski, D.H.; Eldelhauser, H.F.; O’Brien, W.J. Hexose-monophosphate shunt response to diamide in the component layers of the cornea. Exp. Eye Res. 1978, 26, 611–619. [Google Scholar] [CrossRef]
- Moreno, V.C.; Aguilella-Arzo, M.; Del Castillo, R.M.; Espinós, F.J.; Del Castillo, L.F. A refined model on flow and oxygen consumption in the human cornea depending on the oxygen tension at the interface cornea/post lens tear film during contact lens wear. J. Optom. 2022, 15, 160–174. [Google Scholar] [CrossRef]
- Leung, B.K.; Bonanno, J.A.; Radke, C.J. Oxygen-deficient metabolism and corneal edema. Prog. Retin Eye Res. 2011, 30, 471–492. [Google Scholar] [CrossRef] [Green Version]
- Kryczka, T.; Ehlers, N.; Nielsen, K.; Midelfart, A. Impact of organ culturing on metabolic profile of human corneas: Preliminary results. Acta Ophthalmol. 2012, 90, 761–767. [Google Scholar] [CrossRef]
- Craig, J.P.; Nichols, K.K.; Akpek, E.K.; Caffery, B.; Dua, H.S.; Joo, C.-K.; Liu, Z.; Nelson, J.D.; Nichols, J.J.; Tsubota, K.; et al. TFOS DEWS II Definition and Classification Report. Ocul. Surf. 2017, 15, 276–283. [Google Scholar] [CrossRef] [PubMed]
- Rantamaki, A.H.; Seppanen-Laakso, T.; Oresic, M.; Jauhiainen, M.; Holopainen, J.M. Human tear fluid lipidome: From composition to function. PLoS ONE 2011, 6, e19553. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jones, L.; Downie, L.E.; Korb, D.; Benitez-Del-Castillo, J.M.; Dana, R.; Deng, S.X.; Dong, P.N.; Geerling, G.; Hida, R.Y.; Liu, Y.; et al. TFOS DEWS II Management and Therapy Report. Ocul Surf. 2017, 15, 575–628. [Google Scholar]
- Magny, R.; Kessal, K.; Regazzetti, A.; Ben Yedder, A.; Baudouin, C.; Parsadaniantz, S.M.; Brignole-Baudouin, F.; Laprévote, O.; Auzeil, N. Lipidomic analysis of epithelial corneal cells following hyperosmolarity and benzalkonium chloride exposure: New insights in dry eye disease. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2020, 1865, 158728. [Google Scholar] [CrossRef]
- Garrett, Q.; Khandekar, N.; Shih, S.; Flanagan, J.L.; Simmons, P.; Vehige, J.; Willcox, M.D. Betaine stabilizes cell volume and protects against apoptosis in human corneal epithelial cells under hyperosmotic stress. Exp. Eye Res. 2013, 108, 33–41. [Google Scholar] [CrossRef]
- Hua, X.; Su, Z.; Deng, R.; Lin, J.; Li, D.Q.; Pflugfelder, S.C. Effects of L-carnitine, erythritol and betaine on pro-inflammatory markers in primary human corneal epithelial cells exposed to hyperosmotic stress. Curr. Eye Res. 2015, 40, 657–667. [Google Scholar] [CrossRef]
- Abbate, I.; Zappulla, C.; Santonocito, M.; Viola, S.; La Rosa, L.R.; De Pasquale, G.; Caviola, E.; Meloni, M.; Curatolo, M.C.; Mazzone, M.G. Preclinical study of a new matrix to help the ocular surface in dry eye disease. Exp. Eye Res. 2022, 222, 109168. [Google Scholar] [CrossRef]
- Nakatsukasa, M.; Sotozono, C.; Shimbo, K.; Ono, N.; Miyano, H.; Okano, A.; Hamuro, J.; Kinoshita, S. Amino Acid profiles in human tear fluids analyzed by high-performance liquid chromatography and electrospray ionization tandem mass spectrometry. Am. J. Ophthalmol. 2011, 151, 799–808.e1. [Google Scholar] [CrossRef]
- Arribas-Lopez, E.; Zand, N.; Ojo, O.; Snowden, M.J.; Kochhar, T. The Effect of Amino Acids on Wound Healing: A Systematic Review and Meta-Analysis on Arginine and Glutamine. Nutrients 2021, 13, 2498. [Google Scholar] [CrossRef]
- Charkoftaki, G.; Jester, J.V.; Thompson, D.C.; Vasiliou, V. Nitrogen mustard-induced corneal injury involves the sphingomyelin-ceramide pathway. Ocul. Surf. 2018, 16, 154–162. [Google Scholar] [CrossRef]
- Shtein, R.M.; Shen, J.F.; Kuo, A.N.; Hammersmith, K.M.; Li, J.Y.; Weikert, M.P. Autologous Serum-Based Eye Drops for Treatment of Ocular Surface Disease: A Report by the American Academy of Ophthalmology. Ophthalmology 2020, 127, 128–133. [Google Scholar] [CrossRef]
- Quartieri, E.; Marraccini, C.; Merolle, L.; Pulcini, S.; Buzzi, M.; Guardi, M.; Schiroli, D.; Baricchi, R.; Pertinhez, T.A. Metabolomics comparison of cord and peripheral blood-derived serum eye drops for the treatment of dry eye disease. Transfus. Apher. Sci. 2021, 60, 103155. [Google Scholar] [CrossRef]
- Choi, J.J.; Hwang, J.S.; Shin, Y.J. Effect of Oral Choline Alfoscerate on Patients with Keratoconjunctivitis Sicca. Nutrients 2020, 12, 1526. [Google Scholar] [CrossRef]
- Dickinson, H.; Bain, E.; Wilkinson, D.; Middleton, P.; Crowther, C.A.; Walker, D.W. Creatine for women in pregnancy for neuroprotection of the fetus. Cochrane. Database Syst. Rev. 2013, 2014, CD010846. [Google Scholar] [CrossRef]
- Nakamura, S.; Shibuya, M.; Nakashima, H.; Imagawa, T.; Uehara, M.; Tsubota, K. D-beta-hydroxybutyrate protects against corneal epithelial disorders in a rat dry eye model with jogging board. Investig. Ophthalmol. Vis. Sci. 2005, 46, 2379–2387. [Google Scholar] [CrossRef] [Green Version]
- Kawakita, T.; Uchino, M.; Fukagawa, K.; Yoshino, K.; Shimazaki, S.; Toda, I.; Tanaka, M.; Arai, H.; Sakatani, K.; Hata, S.; et al. Randomized, Multicenter, Double-Blind Study of the Safety and Efficacy of 1%D-3-Hydroxybutyrate eye drops for Dry Eye Disease. Sci. Rep. 2016, 6, 20855. [Google Scholar] [CrossRef] [Green Version]
- Hong, J.H.; Li, Q.; Kim, M.S.; Shin, D.M.; Feske, S.; Birnbaumer, L.; Cheng, K.T.; Ambudkar, I.S.; Muallem, S. Polarized but differential localization and recruitment of STIM1, Orai1 and TRPC channels in secretory cells. Traffic 2011, 12, 232–245. [Google Scholar] [CrossRef] [Green Version]
- Teos, L.Y.; Zhang, Y.; Cotrim, A.P.; Swaim, W.; Won, J.H.; Ambrus, J.; Shen, L.; Bebris, L.; Grisius, M.; Jang, S.-I.; et al. IP3R deficit underlies loss of salivary fluid secretion in Sjögren’s Syndrome. Sci. Rep. 2015, 5, 13953. [Google Scholar] [CrossRef] [Green Version]
- Inaba, T.; Hisatsune, C.; Sasaki, Y.; Ogawa, Y.; Ebisui, E.; Ogawa, N.; Matsui, M.; Takeuchi, T.; Mikoshiba, K.; Tsubota, K. Mice lacking inositol 1,4,5-trisphosphate receptors exhibit dry eye. PLoS ONE 2014, 9, e99205. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.D.; Kim, H.Y.; Park, J.J.; Oh, S.B.; Goo, H.; Cho, K.J.; Kim, S.; Kim, K.-B. Metabolomics approach to biomarkers of dry eye disease using (1)H-NMR in rats. J. Toxicol. Environ. Health Part A 2021, 84, 313–330. [Google Scholar] [CrossRef]
- Infantino, V.; Iacobazzi, V.; Menga, A.; Avantaggiati, M.L.; Palmieri, F. A key role of the mitochondrial citrate carrier (SLC25A1) in TNFα- and IFNγ-triggered inflammation. Biochim. Biophys. Acta 2014, 1839, 1217–1225. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.; Rao, J.; Zheng, Z.; Yu, Y.; Lou, S.; Liu, L.; He, Q.; Wu, L.; Sun, X. Integrated Tear Proteome and Metabolome Reveal Panels of Inflammatory-Related Molecules via Key Regulatory Pathways in Dry Eye Syndrome. J. Proteome Res. 2019, 18, 2321–2330. [Google Scholar] [CrossRef]
- Calder, P.C. Polyunsaturated fatty acids and inflammatory processes: New twists in an old tale. Biochimie 2009, 91, 791–795. [Google Scholar] [CrossRef]
- Galbis-Estrada, C.; Pinazo-Durán, M.D.; Martínez-Castillo, S.; Morales, J.M.; Monleón, D.; Zanon-Moreno, V. A metabolomic approach to dry eye disorders. The role of oral supplements with antioxidants and omega 3 fatty acids. Mol. Vis. 2015, 21, 555–567. [Google Scholar]
- Hwang, J.S.; Shin, Y.J. Role of Choline in Ocular Diseases. Int. J. Mol. Sci. 2021, 22, 4733. [Google Scholar] [CrossRef]
- Urbanski, G.; Assad, S.; Chabrun, F.; Chao de la Barca, J.M.; Blanchet, O.; Simard, G.; Lenaers, G.; Prunier-Mirebeau, D.; Gohier, P.; Lavigne, C.; et al. Tear metabolomics highlights new potential biomarkers for differentiating between Sjogren’s syndrome and other causes of dry eye. Ocul. Surf. 2021, 22, 110–116. [Google Scholar] [CrossRef]
- Wei, Y.; Asbell, P.A. sPLA(2)-IIa participates in ocular surface inflammation in humans with dry eye disease. Exp. Eye Res. 2020, 201, 108209. [Google Scholar] [CrossRef]
- Chhabra, M.; Prausnitz, J.M.; Radke, C.J. Modeling corneal metabolism and oxygen transport during contact lens wear. Optom. Vis. Sci. 2009, 86, 454–466. [Google Scholar] [CrossRef]
- Butovich, I.A. Tear film lipids. Exp. Eye Res. 2013, 117, 4–27. [Google Scholar] [CrossRef] [Green Version]
- Gronert, K.; Modi, A.; Asfaha, K.; Chen, S.; Dow, E.; Joslin, S.; Chemaly, M.; Fadli, Z.; Sonoda, L.; Liang, B. Silicone hydrogel contact lenses retain and document ocular surface lipid mediator profiles. Clin. Exp. Optom. 2022, 5, 1–9. [Google Scholar] [CrossRef]
- Masoudi, S.; Willcox, M. A method for studying lipid adsorption to silicone hydrogel contact lenses. Biofouling 2021, 37, 862–878. [Google Scholar] [CrossRef] [PubMed]
- Panaser, A.; Tighe, B.J. Evidence of lipid degradation during overnight contact lens wear: Gas chromatography mass spectrometry as the diagnostic tool. Investig. Ophthalmol. Vis. Sci. 2014, 55, 1797–1804. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Panaser, A.; Tighe, B.J. Function of lipids—Their fate in contact lens wear: An interpretive review. Cont. Lens Anterior. Eye. 2012, 35, 100–111. [Google Scholar] [CrossRef] [PubMed]
- Brown, S.H.J.; Kunnen, C.M.E.; Duchoslav, E.; Dolla, N.K.; Kelso, M.J.; Papas, E.B.; de la Jara, P.L.; Willcox, M.D.P.; Blanksby, S.J.; Mitchell, T.W. A comparison of patient matched meibum and tear lipidomes. Investig. Ophthalmol. Vis. Sci. 2013, 54, 7417–7424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, M.; Liu, L.; Qu, C.; Shi, Y.; Sun, L.; Zhou, X.; Zou, J. Metabolomic Analysis in Corneal Lenticules from Contact Lens Wearers. J. Refract. Surg. 2020, 36, 317–325. [Google Scholar] [CrossRef]
- Yang, L.W.Y.; Mehta, J.S.; Liu, Y.C. Corneal neuromediator profiles following laser refractive surgery. Neural Regen. Res. 2021, 16, 2177–2283. [Google Scholar]
- Liu, Y.C.; Jung, A.S.J.; Chin, J.Y.; Yang, L.W.Y.; Mehta, J.S. Cross-sectional Study on Corneal Denervation in Contralateral Eyes Following SMILE Versus LASIK. J. Refract. Surg. 2020, 36, 653–660. [Google Scholar] [CrossRef]
- Gao, Y.; Qi, Y.; Huang, Y.; Li, X.; Zhou, L.; Zhao, S. Lipidomics Analysis of the Tears in the Patients Receiving LASIK, FS-LASIK, or SBK Surgery. Front. Med. 2021, 8, 731462. [Google Scholar] [CrossRef]
- Bucolo, C.; Fidilio, A.; Platania, C.B.M.; Geraci, F.; Lazzara, F.; Drago, F. Antioxidant and Osmoprotecting Activity of Taurine in Dry Eye Models. J. Ocul. Pharmacol. Ther. 2018, 34, 188–194. [Google Scholar] [CrossRef]
- Chen, J.; Lan, J.; Liu, D.; Backman, L.J.; Zhang, W.; Zhou, Q.; Danielson, P. Ascorbic Acid Promotes the Stemness of Corneal Epithelial Stem/Progenitor Cells and Accelerates Epithelial Wound Healing in the Cornea. Stem. Cells Transl. Med. 2017, 6, 1356–1365. [Google Scholar] [CrossRef]
- Qiu, Y.; Yang, X.; Wang, L.; Gao, K.; Jiang, Z. L-Arginine Inhibited Inflammatory Response and Oxidative Stress Induced by Lipopolysaccharide via Arginase-1 Signaling in IPEC-J2 Cells. Int. J. Mol. Sci. 2019, 20, 1800. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Y.H.; Lee, C.H.; Hsiao, C.C.; Hsu, L.S.; Lo, H.C. Diverse effects of parenteral arginine on systemic and local oxidant-antioxidant homeostasis and nitrosative stress in rats with subacute peritonitis. J. Surg. Res. 2014, 187, 134–141. [Google Scholar] [CrossRef]
- Teo, A.W.J.; Mansoor, H.; Sim, N.; Lin, M.T.; Liu, Y.C. In Vivo Confocal Microscopy Evaluation in Patients with Keratoconus. J. Clin. Med. 2022, 11, 393. [Google Scholar] [CrossRef]
- Karamichos, D.; Hutcheon, A.E.; Rich, C.B.; Trinkaus-Randall, V.; Asara, J.M.; Zieske, J.D. In vitro model suggests oxidative stress involved in keratoconus disease. Sci. Rep. 2014, 4, 4608. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karamichos, D.; Zieske, J.D.; Sejersen, H.; Sarker-Nag, A.; Asara, J.M.; Hjortdal, J. Tear metabolite changes in keratoconus. Exp. Eye Res. 2015, 132, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Wojakowska, A.; Pietrowska, M.; Widlak, P.; Dobrowolski, D.; Wylegala, E.; Tarnawska, D. Metabolomic Signature Discriminates Normal Human Cornea from Keratoconus-A Pilot GC/MS Study. Molecules 2020, 25, 2933. [Google Scholar] [CrossRef] [PubMed]
- Kryczka, T.; Ehlers, N.; Nielsen, K.; Wylegala, E.; Dobrowolski, D.; Midelfart, A. Metabolic profile of keratoconic cornea. Curr. Eye Res. 2013, 38, 305–309. [Google Scholar] [CrossRef] [PubMed]
- Snytnikova, O.A.; Yanshole, L.V.; Iskakov, I.A.; Yanshole, V.V.; Chernykh, V.V.; Stepakov, D.A.; Novoselov, V.P.; Tsentalovich, Y.P. Quantitative metabolomic analysis of the human cornea and aqueous humor. Metabolomics 2017, 13, 152. [Google Scholar] [CrossRef]
- Karamichos, D.; Zareian, R.; Guo, X.; Hutcheon, A.E.; Ruberti, J.W.; Zieske, J.D. Novel in Vitro Model for Keratoconus Disease. J. Funct. Biomater. 2012, 3, 760–775. [Google Scholar] [CrossRef] [Green Version]
- Albaugh, V.L.; Mukherjee, K.; Barbul, A. Proline Precursors and Collagen Synthesis: Biochemical Challenges of Nutrient Supplementation and Wound Healing. J. Nutr. 2017, 147, 2011–2017. [Google Scholar] [CrossRef] [Green Version]
- Song, Y.; Overmass, M.; Fan, J.; Hodge, C.; Sutton, G.; Lovicu, F.J.; You, J. Application of Collagen I and IV in Bioengineering Transparent Ocular Tissues. Front. Surg. 2021, 8, 639500. [Google Scholar] [CrossRef] [PubMed]
- McKay, T.B.; Priyadarsini, S.; Rowsey, T.; Karamichos, D. Arginine Supplementation Promotes Extracellular Matrix and Metabolic Changes in Keratoconus. Cells 2021, 10, 2076. [Google Scholar] [CrossRef] [PubMed]
- Wollensak, G.; Spoerl, E.; Seiler, T. Riboflavin/ultraviolet-a-induced collagen crosslinking for the treatment of keratoconus. Am. J. Ophthalmol. 2003, 135, 620–627. [Google Scholar] [CrossRef] [PubMed]
- Sharif, R.; Sejersen, H.; Frank, G.; Hjortdal, J.; Karamichos, D. Effects of collagen cross-linking on the keratoconus metabolic network. Eye 2018, 32, 1271–1281. [Google Scholar] [CrossRef] [Green Version]
- Sağlik, A.; Koyuncu, İ.; Soydan, A.; Sağlik, F.; Gönel, A. Tear Organic Acid Analysis After Corneal Collagen Crosslinking in Keratoconus. Eye Contact Lens. 2020, 46 (Suppl 2), S122–S128. [Google Scholar] [CrossRef]
- Liu, Y.-C.; So, W.; Wong, N.Q.; Tan, H.; Lin, M.Y.; Lee, I.Y.; Mehta, J. Diabetic corneal neuropathy as a surrogate marker for diabetic peripheral neuropathy. Neural Regen. Res. 2022, 17, 2172–2178. [Google Scholar] [CrossRef]
- Leszczynska, A.; Kulkarni, M.; Ljubimov, A.V.; Saghizadeh, M. Exosomes from normal and diabetic human corneolimbal keratocytes differentially regulate migration, proliferation and marker expression of limbal epithelial cells. Sci. Rep. 2018, 8, 15173. [Google Scholar] [CrossRef] [Green Version]
- Brunmair, J.; Bileck, A.; Schmidl, D.; Hagn, G.; Meier-Menches, S.M.; Hommer, N.; Schlatter, A.; Gerner, C.; Garhöfer, G. Metabolic phenotyping of tear fluid as a prognostic tool for personalised medicine exemplified by T2DM patients. EPMA J. 2022, 13, 107–123. [Google Scholar] [CrossRef]
- Sun, Y.; Gao, H.Y.; Fan, Z.Y.; He, Y.; Yan, Y.X. Metabolomics Signatures in Type 2 Diabetes: A Systematic Review and Integrative Analysis. J. Clin. Endocrinol. Metab. 2020, 105, 1000–1008. [Google Scholar] [CrossRef]
- Wang, T.J.; Larson, M.G.; Vasan, R.S.; Cheng, S.; Rhee, E.P.; McCabe, E.; Lewis, G.D.; Fox, C.S.; Jacques, P.F.; Fernandez, C.; et al. Metabolite profiles and the risk of developing diabetes. Nat. Med. 2011, 17, 448–453. [Google Scholar] [CrossRef] [Green Version]
- Yoon, M.S. The Emerging Role of Branched-Chain Amino Acids in Insulin Resistance and Metabolism. Nutrients 2016, 8, 405. [Google Scholar] [CrossRef] [Green Version]
- King, C.; Lanaspa, M.A.; Jensen, T.; Tolan, D.R.; Sánchez-Lozada, L.G.; Johnson, R.J. Uric Acid as a Cause of the Metabolic Syndrome. Contrib. Nephrol. 2018, 192, 88–102. [Google Scholar] [PubMed]
- ChenZhuo, L.; Murube, J.; Latorre, A.; del Río, R.M. Different concentrations of amino acids in tears of normal and human dry eyes. Adv. Exp. Med. Biol. 2002, 506, 617–621. [Google Scholar] [PubMed]
- He, F.; Zhao, Z.; Liu, Y.; Lu, L.; Fu, Y. Assessment of Ocular Surface Damage during the Course of Type 2 Diabetes Mellitus. J. Ophthalmol. 2018, 2018, 1206808. [Google Scholar] [CrossRef]
- Sak, D.; Erdenen, F.; Müderrisoglu, C.; Altunoglu, E.; Sozer, V.; Gungel, H.; Guler, P.A.; Sak, T.; Uzun, H. The Relationship between Plasma Taurine Levels and Diabetic Complications in Patients with Type 2 Diabetes Mellitus. Biomolecules 2019, 9, 96. [Google Scholar] [CrossRef] [Green Version]
- Priyadarsini, S.; McKay, T.B.; Sarker-Nag, A.; Allegood, J.; Chalfant, C.; Ma, J.-X.; Karamichos, D. Complete metabolome and lipidome analysis reveals novel biomarkers in the human diabetic corneal stroma. Exp. Eye Res. 2016, 153, 90–100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guitton, J.; Bandet, C.L.; Mariko, M.L.; Tan-Chen, S.; Bourron, O.; Benomar, Y.; Hajduch, E.; Le Stunff, H. Sphingosine-1-Phosphate Metabolism in the Regulation of Obesity/Type 2 Diabetes. Cells 2020, 9, 1682. [Google Scholar] [CrossRef]
- Matysik-Woźniak, A.; Turski, W.; Turska, M.; Paduch, R.; Łańcut, M.; Piwowarczyk, P.; Czuczwar, M.; Rejdak, R. Kynurenic Acid Accelerates Healing of Corneal Epithelium In Vitro and In Vivo. Pharmaceuticals 2021, 14, 753. [Google Scholar] [CrossRef]
- Matysik-Wozniak, A.; Paduch, R.; Turski, W.A.; Maciejewski, R.; Junemann, A.G.; Rejdak, R. Effects of tryptophan, kynurenine and kynurenic acid exerted on human reconstructed corneal epithelium in vitro. Pharmacol. Rep. 2017, 69, 722–729. [Google Scholar] [CrossRef]
- Serbecic, N.; Lahdou, I.; Scheuerle, A.; Höftberger, R.; Aboul-Enein, F. Function of the tryptophan metabolite, L-kynurenine, in human corneal endothelial cells. Mol. Vis. 2009, 15, 1312–1324. [Google Scholar]
- Karamichos, D.; Guo, X.Q.; Hutcheon, A.E.; Zieske, J.D. Human corneal fibrosis: An in vitro model. Investig. Ophthalmol. Vis. Sci. 2010, 51, 1382–1388. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Whelchel, A.E.; Nicholas, S.E.; Ma, J.X.; Karamichos, D. Nerve influence on the metabolism of type I and type II diabetic corneal stroma: An in vitro study. Sci. Rep. 2021, 11, 13627. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
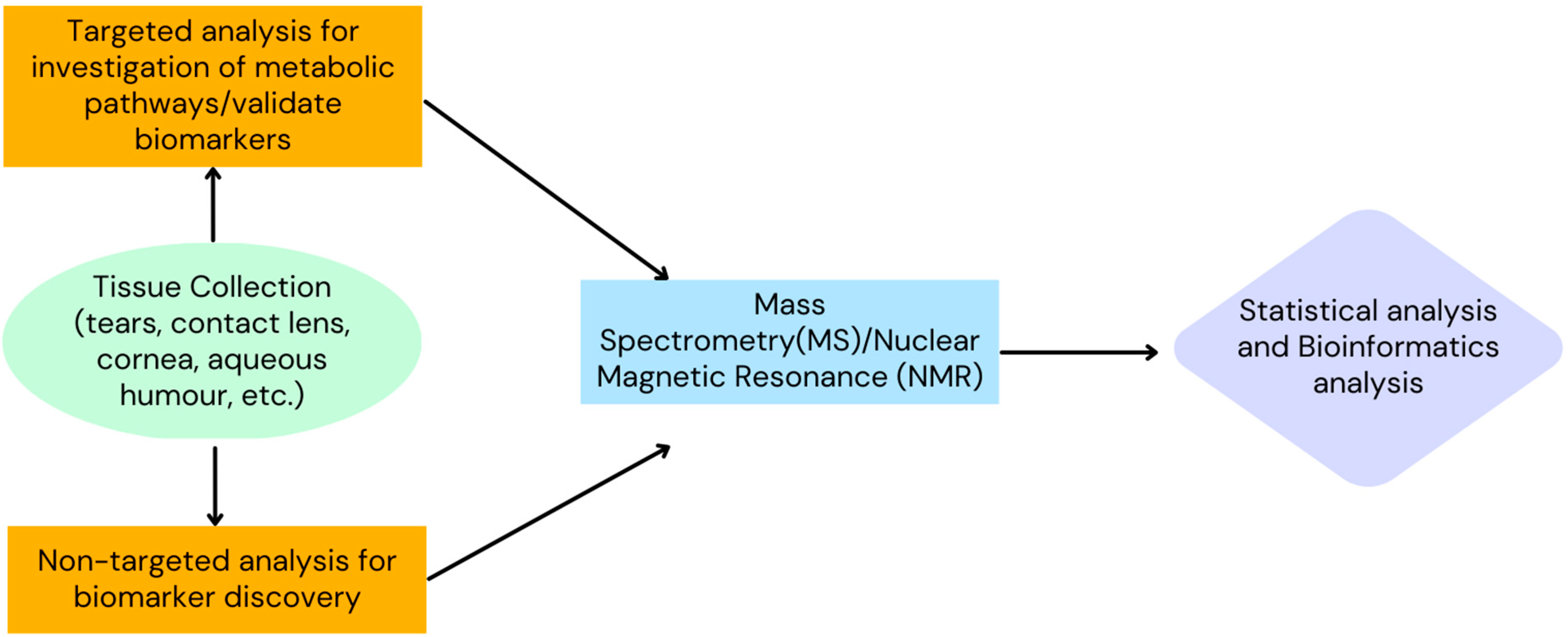{kind=link}
| Author | Metabolomic Assessment Method | Sample Source | Findings | Clinical Implications |
|---|---|---|---|---|
| Dry Eye Disease (DED) | ||||
| Lee et al. [51] | Nuclear magnetic resonance (NMR) | Rat tears, plasma, and urine | There was an increase in pro-inflammatory cytokines, such as IL-6, IL-1β, and TNF-α. 2-hydroxybutyrate, citrate and succinate, which play key roles in inflammatory pathways, had elevated levels in plasma. | Identified metabolites may serve as potential biomarkers for DED. |
| Chen et al. [10] | Matrix-assisted laser desorption ionization mass spectrometry imaging (MALDI-MSI) | Cornea, conjunctiva, and aqueous humor from rats | Glycerophospholipid and phenylalanine metabolism were altered, implicating their roles in signal transduction and tear film stability.Increase in betaine suggests early cytoprotective mechanism against dry eye. | MALDI-MSI can accurately analyze the different metabolic responses of complex eye components in DED, and can potentially be used to individualize treatment. |
| Quartieri et al. [43] | NMR | CBS and PBS eye drops | Metabolites measured in CBS and PBS eye drops were higher than tears. However, CBS drops contained higher amounts of myoinositol, choline, glutamine, creatine, and beta-hydroxybutyrate, which serve as anti-oxidants, and work in pathways to ameliorate DED. | Serum eye drops contain important metabolites to influence cellular signaling in hopes of alleviating DED. |
| Galbis-Estrada et al. [55] | NMR | Tears | There was an increase in choline after essential polyunsaturated fatty acids (EPUFA) supplementation—this stabilizes the tear film and has anti-inflammatory effects on ocular surface. | Changes in tear metabolic profile of DED can be modified by oral supplementation of antioxidants and EPUFAs. |
| Sjogren’s Syndrome | ||||
| Urbanski et al. [57] | Liquid chromatography—mass spectrometry (LC-MS) | Tears | 9 metabolites could be used to distinguish PSS and DED. | Metabolomic signature of tears could distinguish PSS from DED. |
| Contact Lens Wear | ||||
| Li et al. [66] | LC-MS | Lenticules extracted via SMILE from soft contact lens-wearing patients | Upregulation of short chain organic acids indicate decreased respiration of glucose and switch to anaerobic respiration process in patients who wore contact lenses. | In the corneal stroma, there is significant change in energy metabolism in the corneal stroma after wearing soft contact lenses. |
| Refractive Surgery | ||||
| Li et al. [13] | LC-MS | Lenticules extracted from SMILE | There was a higher concentration of inflammatory-related metabolites and lower anti-oxidants in older patients. This influenced rates of corneal wound healing. | Corneal wound healing and corneal nerve recovery after SMILE was significantly affected by age. |
| Keratoconus | ||||
| Karamichos et al. [75] | LC-MS | Human corneal keratocytes (HCKs), fibroblasts (HCFs), and keratoconus cells (HKCs) cultured in 2D and 3D in vitro systems | Lactate levels and lactate/malate and lactate/pyruvate ratios were elevated in HKCs, while arginine and GSH/GSSG ratios were reduced, indicative of oxidative stress. | Future studies may help to identify novel pathways that may lead to metabolic therapies for keratoconus. |
| Wojakowska et al. [77] | Gas chromatography–MS | Corneal buttons | Downregulation of unsaturated fatty acids such as linoleic acid indicate reduced ability for cornea repair due to their anti-inflammatory effects. Succinic acid, playing an important role in tricarboxylic acid cycle, was downregulated. | Metabolomic signatures indicate oxidative stress and inflammatory reactions are involved in the development of keratoconus. |
| Kryczka et al. [78] | NMR and high-performance liquid chromatography | Corneal buttons from cadavers and keratoconic corneas | No major differences in metabolic contents between older, normal corneas and keratoconic corneas. | Young keratoconic corneas are biochemically like older normal corneas, which may indicate accelerated aging of the cornea in keratoconus. |
| Mckay et al. [83] | LC-MS | HKCs in a 3D in vitro construct | Lower cytoplasmic arginine and spermidine levels in KC constructs compared with healthy controls. Arginine supplementation led to a robust increase in cytoplasmic arginine, ornithine, and spermidine levels in controls only, and a significant increase in type I collagen secretion in HKC constructs. | Arginine supplementation may support increased collagen type I secretion by HKCs. |
| Saglik et al. [86] | LC-MS | Tears | N-acetyl-L-aspartic acid, which is known to have anti-oxidant effects, showed the greatest percentage increase after CXL.3-OH butyric acid showed the greatest decrease, suggesting improvement in glucose metabolism after CXL. | Metabolomic studies of tears could derive new understanding of processes in the follow-up period after CXL. |
| Snytnikova et al. [79] | NMR and LC-MS | Corneal buttons and aqueous humor from KC patients and normal corneas from cadavers | Levels of glucose in KC cornea was almost tenfold lower than in aqueous humor, demonstrating glucose consumption in vivo. Enhanced levels of citrate and acetate in KC corneas relate to oxidative stress, and is supported by low values of GSH/GSSG ratio in KC corneas. | Oxidative stress may be involved in keratoconus pathology. |
| Diabetic Corneas | ||||
| Brunmair et al. [89] | LC-MS | Tears | Upregulation of amino acids in tears is thought to act via the same pathways as insulin such as mammalian target of rapamycin (mTOR) and its downstream targets which act via the same pathways as insulin.Taurine accumulation in tears could indicate a stressed state. | High number of identified molecule markers may support disease development prediction, preventative approaches, personalized patients’ treatments, and monitoring treatment efficacy. |
| Priyadarsini et al. [97] | LC-MS | Cadaver cornea buttons | T1DM showed upregulation of kynurenic acid, suggesting dysregulation of kynurenine metabolism. Kynurenic acid is known to have anti-oxidant, anti-inflammatory, and anti-proliferative properties. | The kynurenine pathway is a potential therapeutic target to prevent T1DM-related complications to the eye. |
| Whelchel et al. [103] | LC-MS | In vitro 3D non-innervated constructs vs innervated constructs | Addition of neuron-supported pathways such as pyrimidine metabolism, glycerol phosphate shuttle, electron transport chain, and glycolysis.Myoinositol dysregulation was related to neuronal dysfunction in diabetic corneas. | Corneal innervation has impact on the metabolism of diabetic corneal stroma. |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Teo, A.W.J.; Zhang, J.; Zhou, L.; Liu, Y.-C. Metabolomics in Corneal Diseases: A Narrative Review from Clinical Aspects. Metabolites 2023, 13, 380. https://doi.org/10.3390/metabo13030380
Teo AWJ, Zhang J, Zhou L, Liu Y-C. Metabolomics in Corneal Diseases: A Narrative Review from Clinical Aspects. Metabolites. 2023; 13(3):380. https://doi.org/10.3390/metabo13030380
Chicago/Turabian StyleTeo, Alvin Wei Jun, Jingwen Zhang, Lei Zhou, and Yu-Chi Liu. 2023. "Metabolomics in Corneal Diseases: A Narrative Review from Clinical Aspects" Metabolites 13, no. 3: 380. https://doi.org/10.3390/metabo13030380
APA StyleTeo, A. W. J., Zhang, J., Zhou, L., & Liu, Y. -C. (2023). Metabolomics in Corneal Diseases: A Narrative Review from Clinical Aspects. Metabolites, 13(3), 380. https://doi.org/10.3390/metabo13030380