
Metabolomic Profiling in Mouse Model of Menopause-Associated Asthma


Abstract
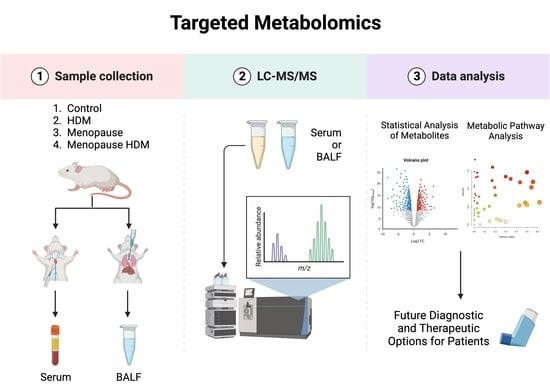
:
1. Introduction
2. Materials and Methods
3. Results
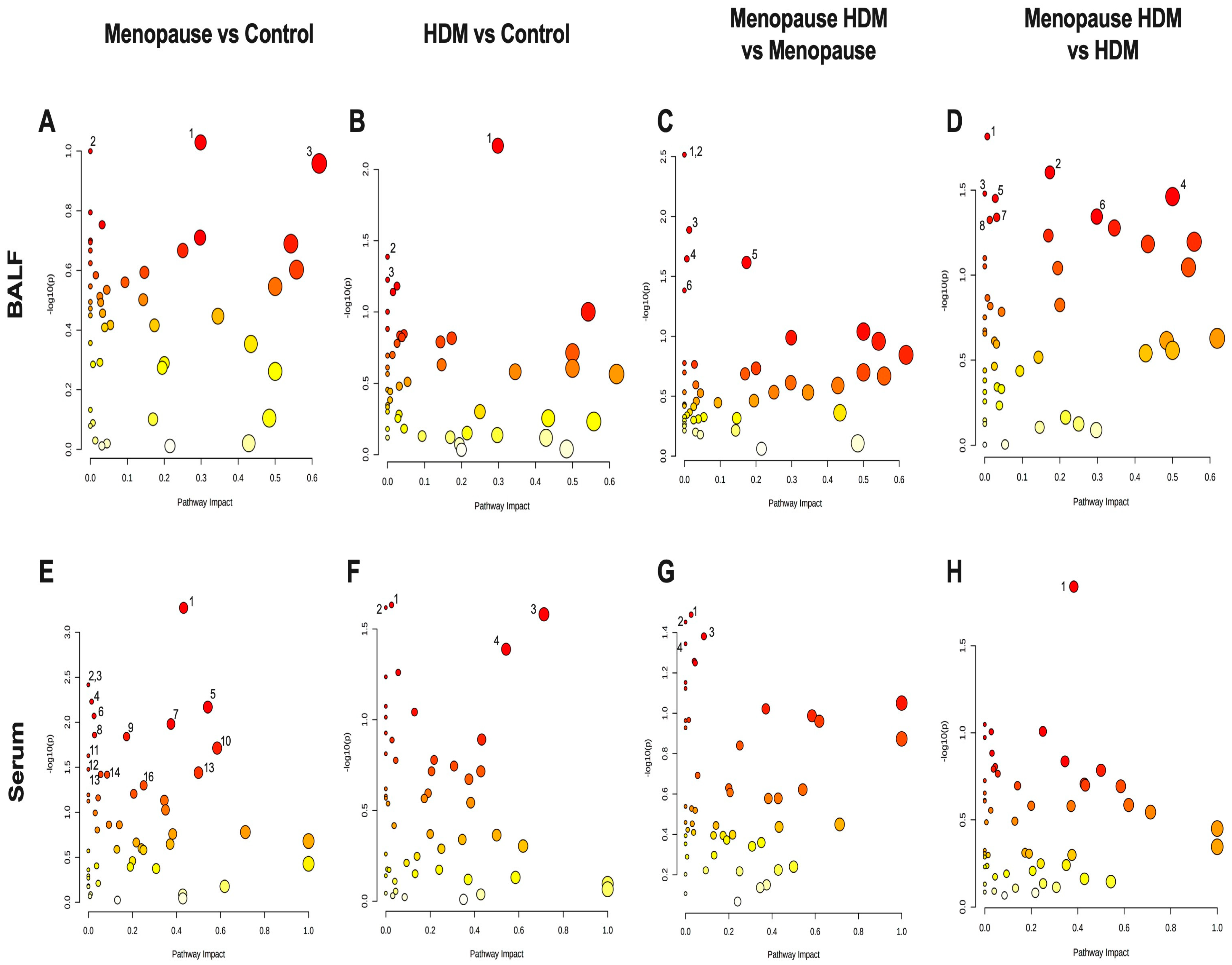
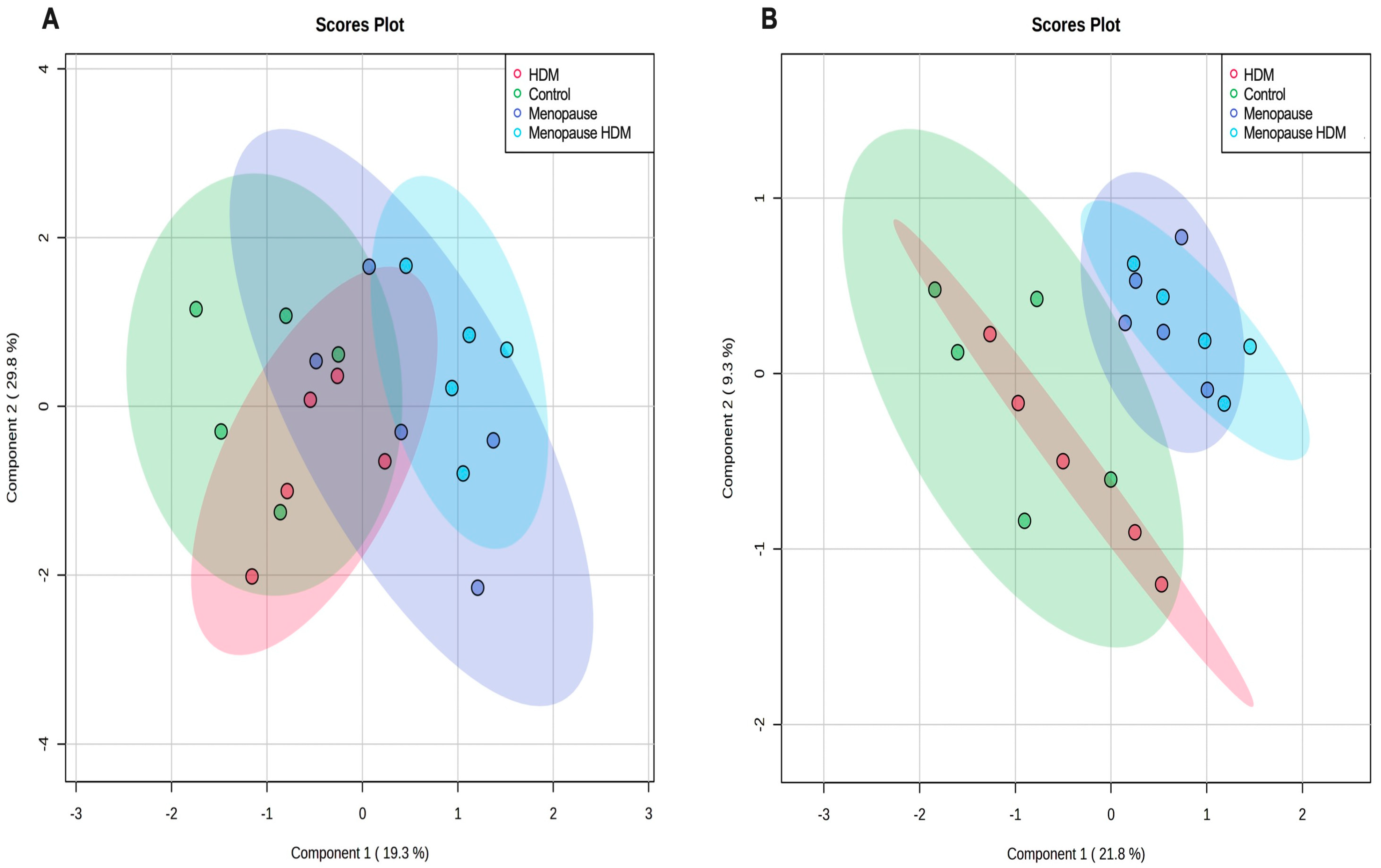
3.1. Impact of Menopause on Metabolomic Profile
3.2. Impact of HDM on Metabolomic Profile
3.3. Impact of HDM in Menopausal Mice on Metabolomic Profile
3.4. Impact of Menopause in HDM-Challenged Mice on Metabolomic Profile
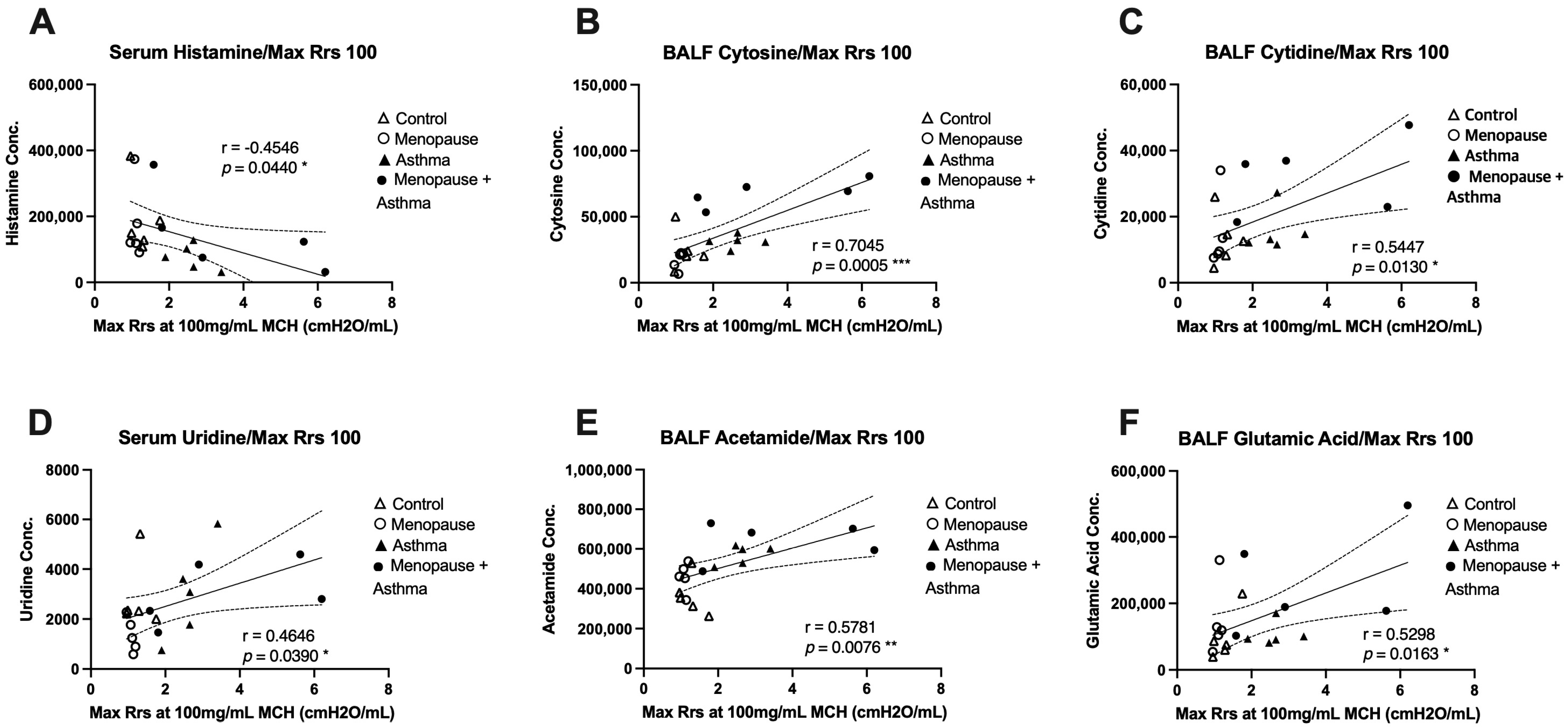
3.5. Associations between Metabolites and Airway Hyper-Responsiveness
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- National Current Asthma Prevalence. Centers for Disease Control and Prevention (CDC). 2019. Available online: https://www.cdc.gov/asthma/most_recent_national_asthma_data.htm (accessed on 1 January 2023).
- Castillo, J.R.; SPPeters Busse, W.W. Asthma Exacerbations: Pathogenesis, Prevention, and Treatment. J. Allergy Clin. Immunol. Pract. 2017, 5, 918–927. [Google Scholar] [CrossRef]
- Maslan, J.; Mims, J.W. What is asthma? Pathophysiology, demographics, and health care costs. Otolaryngol. Clin. N. Am. 2014, 47, 13–22. [Google Scholar] [CrossRef] [PubMed]
- Mims, J.W. Asthma: Definitions and pathophysiology. Int. Forum Allergy Rhinol. 2015, 5 (Suppl. 1), S2–S6. [Google Scholar] [CrossRef]
- Wenzel, S.E. Asthma phenotypes: The evolution from clinical to molecular approaches. Nat. Med. 2012, 18, 716–725. [Google Scholar] [CrossRef] [PubMed]
- Desai, M.; Oppenheimer, J. Elucidating asthma phenotypes and endotypes: Progress towards personalized medicine. Ann. Allergy Asthma Immunol. 2016, 116, 394–401. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Balzano, G.; Fuschillo, S.; De Angelis, E.; Gaudiosi, C.; Mancini, A.; Caputi, M. Persistent airway inflammation and high exacerbation rate in asthma that starts at menopause. Monaldi Arch. Chest Dis. 2007, 67, 135–141. [Google Scholar] [CrossRef]
- Pederson, W.P.; Ellerman, L.M.; Sandoval, E.C.; Boitano, S.; Frye, J.B.; Doyle, K.P.; Brooks, H.L.; Polverino, F.; Ledford, J.G. Development of a Novel Mouse Model of Menopause-associated Asthma. Am. J. Respir. Cell Mol. Biol. 2022, 67, 605–609. [Google Scholar] [CrossRef]
- Balzano, G.; Fuschillo, S.; Melillo, G.; Bonini, S. Asthma and sex hormones. Allergy 2001, 56, 13–20. [Google Scholar] [CrossRef]
- Luxon, B.A. Metabolomics in asthma. Adv. Exp. Med. Biol. 2014, 795, 207–220. [Google Scholar]
- Wang, C.; Jiang, S.; Zhang, S.; Ouyang, Z.; Wang, G.; Wang, F. Research Progress of Metabolomics in Asthma. Metabolites 2021, 11, 567. [Google Scholar] [CrossRef] [PubMed]
- Xu, S.; Panettieri, R.A., Jr.; Jude, J. Metabolomics in asthma: A platform for discovery. Mol. Asp. Med. 2022, 85, 100990. [Google Scholar] [CrossRef] [PubMed]
- Pite, H.; Morais-Almeida, M.; Rocha, S.M. Metabolomics in asthma: Where do we stand? Curr. Opin. Pulm. Med. 2018, 24, 94–103. [Google Scholar] [CrossRef] [PubMed]
- Carroll, P.A.; Diolaiti, D.; McFerrin, L.; Gu, H.; Djukovic, D.; Du, J.; Cheng, P.F.; Anderson, S.; Ulrich, M.; Hurley, J.B.; et al. Deregulated Myc requires MondoA/Mlx for metabolic reprogramming and tumorigenesis. Cancer Cell 2015, 27, 271–285. [Google Scholar] [CrossRef] [Green Version]
- Eghlimi, R.; Shi, X.; Hrovat, J.; Xi, B.; Gu, H. Triple Negative Breast Cancer Detection Using LC-MS/MS Lipidomic Profiling. J. Proteome Res. 2020, 19, 2367–2378. [Google Scholar] [CrossRef]
- Gu, H.; Carroll, P.A.; Du, J.; Zhu, J.; Neto, F.C.; Eisenman, R.N.; Raftery, D. Quantitative Method to Investigate the Balance between Metabolism and Proteome Biomass: Starting from Glycine. Angew. Chem. Int. Ed. Engl. 2016, 55, 15646–15650. [Google Scholar] [CrossRef] [PubMed]
- Gu, H.; Zhang, P.; Zhu, J.; Raftery, D. Globally Optimized Targeted Mass Spectrometry: Reliable Metabolomics Analysis with Broad Coverage. Anal. Chem. 2015, 87, 12355–12362. [Google Scholar] [CrossRef] [Green Version]
- Jasbi, P.; Mitchell, N.M.; Shi, X.; Grys, T.E.; Wei, Y.; Liu, L.; Lake, D.F.; Gu, H. Coccidioidomycosis Detection Using Targeted Plasma and Urine Metabolic Profiling. J. Proteome Res. 2019, 18, 2791–2802. [Google Scholar] [CrossRef]
- Jasbi, P.; Shi, X.; Chu, P.; Elliott, N.; Hudson, H.; Jones, D.; Serrano, G.; Chow, B.; Beach, T.G.; Liu, L.; et al. Metabolic Profiling of Neocortical Tissue Discriminates Alzheimer’s Disease from Mild Cognitive Impairment, High Pathology Controls, and Normal Controls. J. Proteome Res. 2021, 20, 4303–4317. [Google Scholar] [CrossRef]
- Shi, X.; Wang, S.; Jasbi, P.; Turner, C.; Hrovat, J.; Wei, Y.; Liu, J.; Gu, H. Database-Assisted Globally Optimized Targeted Mass Spectrometry (dGOT-MS): Broad and Reliable Metabolomics Analysis with Enhanced Identification. Anal. Chem. 2019, 91, 13737–13745. [Google Scholar] [CrossRef]
- Addison, K.J.; Morse, J.; Robichaud, A.; Daines, M.O.; Ledford, J.G. A Novel in vivo System to Test Bronchodilators. J. Infect. Pulm. Dis. 2017, 3, 10-16966. [Google Scholar]
- Kuruvilla, M.E.; Lee, F.E.; Lee, G.B. Understanding Asthma Phenotypes, Endotypes, and Mechanisms of Disease. Clin. Rev. Allergy Immunol. 2019, 56, 219–233. [Google Scholar] [CrossRef] [PubMed]
- Villasenor, A.; Eguiluz-Gracia, I.; Moreira, A.; Wheelock, C.E.; Escribese, M.M. Metabolomics in the Identification of Biomarkers of Asthma. Metabolites 2021, 11, 346. [Google Scholar] [CrossRef] [PubMed]
- Pal, M.M. Glutamate: The Master Neurotransmitter and Its Implications in Chronic Stress and Mood Disorders. Front. Hum. Neurosci. 2021, 15, 722323. [Google Scholar] [CrossRef] [PubMed]
- Petroff, O.A. GABA and glutamate in the human brain. Neuroscientist 2002, 8, 562–573. [Google Scholar] [CrossRef]
- Barrios, J.; Kho, A.T.; Aven, L.; Mitchel, J.A.; Park, J.A.; Randell, S.H.; Miller, L.A.; Tantisira, K.G.; Ai, X. Pulmonary Neuroendocrine Cells Secrete gamma-Aminobutyric Acid to Induce Goblet Cell Hyperplasia in Primate Models. Am. J. Respir. Cell Mol. Biol. 2019, 60, 687–694. [Google Scholar] [CrossRef]
- Reader, J.R.; Tepper, J.S.; Schelegle, E.S.; Aldrich, M.C.; Putney, L.F.; Pfeiffer, J.W.; Hyde, D.M. Pathogenesis of mucous cell metaplasia in a murine asthma model. Am. J. Pathol. 2003, 162, 2069–2078. [Google Scholar] [CrossRef] [Green Version]
- Xu, C.J.; Klunk, W.E.; Kanfer, J.N.; Xiong, Q.; Miller, G.; Pettegrew, J.W. Phosphocreatine-dependent glutamate uptake by synaptic vesicles. A comparison with atp-dependent glutamate uptake. J. Biol. Chem. 1996, 271, 13435–13440. [Google Scholar] [CrossRef] [Green Version]
- Camateros, P.; Kanagaratham, C.; Najdekr, L.; Holub, D.; Vrbkova, J.; Cote, L.; Fournier, J.; Gourdon, J.; Creery, D.; Olivenstein, R.; et al. Toll-Like Receptor 7/8 Ligand, S28463, Suppresses Ascaris suum-induced Allergic Asthma in Nonhuman Primates. Am. J. Respir. Cell Mol. Biol. 2018, 58, 55–65. [Google Scholar] [CrossRef]
- Burki, N.K.; Diamond, L. Serum creatine phosphokinase activity in asthma. Am. Rev. Respir. Dis. 1977, 116, 327–331. [Google Scholar]
- Al Heialy, S.; Ramakrishnan, R.K.; Hamid, Q. Recent advances in the immunopathogenesis of severe asthma. J. Allergy Clin. Immunol. 2022, 149, 455–465. [Google Scholar] [CrossRef]
- Boonpiyathad, T.; Sozener, Z.C.; Satitsuksanoa, P.; Akdis, C.A. Immunologic mechanisms in asthma. Semin. Immunol. 2019, 46, 101333. [Google Scholar] [CrossRef]
- Newcomb, D.C.; Peebles, R.S., Jr. Th17-mediated inflammation in asthma. Curr. Opin. Immunol. 2013, 25, 755–760. [Google Scholar] [CrossRef] [Green Version]
- Johnson, M.O.; Wolf, M.M.; Madden, M.Z.; Andrejeva, G.; Sugiura, A.; Contreras, D.C.; Maseda, D.; Liberti, M.V.; Paz, K.; Kishton, R.J.; et al. Distinct Regulation of Th17 and Th1 Cell Differentiation by Glutaminase-Dependent Metabolism. Cell 2018, 175, 1780–1795.e19. [Google Scholar] [CrossRef] [Green Version]
- Miao, Y.; Zheng, Y.; Geng, Y.; Yang, L.; Cao, N.; Dai, Y.; Wei, Z. The role of GLS1-mediated glutaminolysis/2-HG/H3K4me3 and GSH/ROS signals in Th17 responses counteracted by PPARgamma agonists. Theranostics 2021, 11, 4531–4548. [Google Scholar] [CrossRef]
- Schilling, S.; Wasternack, C.; Demuth, H.U. Glutaminyl cyclases from animals and plants: A case of functionally convergent protein evolution. Biol. Chem. 2008, 389, 983–991. [Google Scholar] [CrossRef]
- Zhou, Y.; Zhao, H.; Wang, T.; Zhao, X.; Wang, J.; Wang, Q. Anti-Inflammatory and Anti-asthmatic Effects of TMDCT Decoction in Eosinophilic Asthma Through Treg/Th17 Balance. Front. Pharmacol. 2022, 13, 819728. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Ma, P.; Wang, Y.; Hou, B.; Zhou, C.; Tian, H.; Li, B.; Shui, G.; Yang, X.; Qiang, G.; et al. Untargeted metabolomics and transcriptomics identified glutathione metabolism disturbance and PCS and TMAO as potential biomarkers for ER stress in lung. Sci. Rep. 2021, 11, 14680. [Google Scholar] [CrossRef]
- Chiu, C.Y.; Cheng, M.L.; Chiang, M.H.; Wang, C.J.; Tsai, M.H.; Lin, G. Metabolomic Analysis Reveals Distinct Profiles in the Plasma and Urine Associated with IgE Reactions in Childhood Asthma. J. Clin. Med. 2020, 9, 887. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Loureiro, C.C.; Duarte, I.F.; Gomes, J.; Carrola, J.; Barros, A.S.; Gil, A.M.; Bousquet, J.; Bom, A.T.; Rocha, S.M. Urinary metabolomic changes as a predictive biomarker of asthma exacerbation. J. Allergy Clin. Immunol. 2014, 133, 261–263. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Chen, G.; Shu, S.; Xu, Y.; Ma, X. Metabolomics analysis of baicalin on ovalbumin-sensitized allergic rhinitis rats. R. Soc. Open Sci. 2019, 6, 181081. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hashimoto, T.; Ohata, H.; Honda, K. Lysophosphatidic acid (LPA) induces plasma exudation and histamine release in mice via LPA receptors. J. Pharmacol. Sci. 2006, 100, 82–87. [Google Scholar] [CrossRef] [Green Version]
- Trevethick, M.A. Is uridine a treatment for asthma? Clin. Exp. Allergy 2010, 40, 1436–1438. [Google Scholar] [CrossRef]
- Roberts, I.; Wright Muelas, M.; Taylor, J.M.; Davison, A.S.; Xu, Y.; Grixti, J.M.; Gotts, N.; Sorokin, A.; Goodacre, R.; Kell, D.B. Untargeted metabolomics of COVID-19 patient serum reveals potential prognostic markers of both severity and outcome. Metabolomics 2021, 18, 6. [Google Scholar] [CrossRef] [PubMed]
- Blasco, H.; Bessy, C.; Plantier, L.; Lefevre, A.; Piver, E.; Bernard, L.; Marlet, J.; Stefic, K.; Benz-de Bretagne, I.; Cannet, P.; et al. The specific metabolome profiling of patients infected by SARS-COV-2 supports the key role of tryptophan-nicotinamide pathway and cytosine metabolism. Sci. Rep. 2020, 10, 16824. [Google Scholar] [CrossRef] [PubMed]
- Tounta, V.; Liu, Y.; Cheyne, A.; Larrouy-Maumus, G. Metabolomics in infectious diseases and drug discovery. Mol. Omics 2021, 17, 376–393. [Google Scholar] [CrossRef] [PubMed]
- Takhar, P.; Corrigan, C.J.; Smurthwaite, L.; O’Connor, B.J.; Durham, S.R.; Lee, T.H.; Gould, H.J. Class switch recombination to IgE in the bronchial mucosa of atopic and nonatopic patients with asthma. J. Allergy Clin. Immunol. 2007, 119, 213–218. [Google Scholar] [CrossRef]
- Tian, M.; Chen, M.; Bao, Y.L.; Xu, C.D.; Qin, Q.Z.; Zhang, W.X.; He, Y.T.; Shao, Q. Sputum metabolomic profiling of bronchial asthma based on quadruple time-of-flight mass spectrometry. Int. J. Clin. Exp. Pathol. 2017, 10, 10363–10373. [Google Scholar]
- Berry, D.E.; Chan, J.A.; MacKenzie, L.; Hecht, S.M. 9-Octadecynoic acid: A novel DNA binding agent. Chem. Res. Toxicol. 1991, 4, 195–198. [Google Scholar] [CrossRef]
- Meyer, F.; Bloch, K. Metabolism of Stearolic Acid in Yeast. J. Biol. Chem. 1963, 238, 2654–2659. [Google Scholar] [CrossRef]
- Thomasson, H.J. Stearolic acid, an essential fatty acid? Nature 1954, 173, 452. [Google Scholar] [CrossRef]
- Bahado-Singh, R.O.; Akolekar, R.; Mandal, R.; Dong, E.; Xia, J.; Kruger, M.; Wishart, D.S.; Nicolaides, K. Metabolomics and first-trimester prediction of early-onset preeclampsia. J. Matern. Fetal Neonatal Med. 2012, 25, 1840–1847. [Google Scholar] [CrossRef] [PubMed]
- Bahado-Singh, R.O.; Akolekar, R.; Mandal, R.; Dong, E.; Xia, J.; Kruger, M.; Wishart, D.S.; Nicolaides, K. First-trimester metabolomic detection of late-onset preeclampsia. Am. J. Obstet. Gynecol. 2013, 208, 58.e1–58.e7. [Google Scholar] [CrossRef] [PubMed]
- Bahado-Singh, R.O.; Akolekar, R.; Mandal, R.; Dong, E.; Xia, J.; Kruger, M.; Wishart, D.S.; Nicolaides, K. Metabolomic analysis for first-trimester Down syndrome prediction. Am. J. Obstet. Gynecol. 2013, 208, 371.e1–371.e8. [Google Scholar] [CrossRef] [PubMed]
- Bahado-Singh, R.O.; Ertl, R.; Mandal, R.; Bjorndahl, T.C.; Syngelaki, A.; Han, B.; Dong, E.; Liu, P.B.; Alpay-Savasan, Z.; Wishart, D.S.; et al. Metabolomic prediction of fetal congenital heart defect in the first trimester. Am. J. Obstet. Gynecol. 2014, 211, 240.e1–240.e14. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Comparison | Change | Metabolite | p Value | Fold Change |
|---|---|---|---|---|
| Menopause vs. Control | Downregulated | TMAO | 0.007 | 0.11 |
| 2-HydroxybenzoicAcid | 0.012 | 0.30 | ||
| Threonine | 0.014 | 0.59 | ||
| Homoserine | 0.016 | 0.60 | ||
| Hippuric Acid | 0.038 | 0.46 | ||
| Hydroxyproline | 0.046 | 0.63 | ||
| 5-Aminolevulinic Acid | 0.049 | 0.67 | ||
| Upregulated | 4-Aminobutyric Acid | 0.080 | 5.42 | |
| HDM vs. Control | Downregulated | Fructose | 0.041 | 0.60 |
| Ribose | 0.045 | 0.61 | ||
| TMAO | 0.048 | 0.25 | ||
| Upregulated | 9-Octadecynoic Acid | 0.001 | 3.31 | |
| Acetamide | 0.005 | 1.55 | ||
| Glycocyamine | 0.012 | 2.15 | ||
| Menopause HDM vs. Menopause | Upregulated | Cytosine | 0.0003 | 3.96 |
| GlycoCyamine | 0.005 | 1.92 | ||
| 9-Octadecynoic Acid | 0.011 | 2.24 | ||
| D-Ribose 5-phosphate | 0.013 | 2.98 | ||
| Cytidine | 0.023 | 2.21 | ||
| Phenylpyruvic Acid | 0.040 | 1.86 | ||
| Menopause HDM vs. HDM | Upregulated | Cytosine | 0.00005 | 2.17 |
| Cytidine | 0.015 | 2.05 | ||
| Glutamic Acid | 0.033 | 2.44 | ||
| D-Ribose 5 Phosphate | 0.047 | 1.96 |
| Comparison | Change | Metabolite | p Value | Fold Change |
|---|---|---|---|---|
| Menopause vs. Control | Downregulated | 2-Hydroxybenzoic Acid | 0.007 | 0.12 |
| Oxidized Glutathione | 0.007 | 0.61 | ||
| Mucic Acid | 0.008 | 0.38 | ||
| 5-aminolevulinic acid | 0.008 | 0.67 | ||
| Acetylcholine | 0.008 | 0.67 | ||
| Ribose | 0.011 | 0.22 | ||
| Methyl Guanidine | 0.011 | 0.34 | ||
| Xylose | 0.011 | 0.19 | ||
| Hydroxyproline | 0.012 | 0.65 | ||
| Carnosine | 0.032 | 0.59 | ||
| Uridine | 0.031 | 0.47 | ||
| GA3P | 0.035 | 0.33 | ||
| Upregulated | Pyroglutamic Acid | 0.0003 | 2.00 | |
| Glucosamine | 0.006 | 1.63 | ||
| 3-hydroxyisovaleric Acid | 0.007 | 1.63 | ||
| Myristic Acid | 0.015 | 1.66 | ||
| Nicotinuric Acid | 0.020 | 1.83 | ||
| 4-Pyridoxic Acid | 0.024 | 1.73 | ||
| Oxoglutaric Acid | 0.032 | 1.82 | ||
| 2-Methylglutaric Acid | 0.037 | 1.77 | ||
| Indole 3-acetic Acid | 0.042 | 2.33 | ||
| HDM vs. Control | Downregulated | 3-Methyladipic Acid | 0.005 | 0.66 |
| Methyl-D-Mannopyranoside | 0.023 | 0.61 | ||
| Histamine | 0.026 | 0.40 | ||
| Carnosine | 0.045 | 0.64 | ||
| Menopause HDM vs. Menopause | Upregulated | Uridine | 0.028 | 2.27 |
| Acetylcholine | 0.034 | 1.66 | ||
| Menopause HDM vs. HDM | Upregulated | Phosphocreatine | 0.014 | 1.86 |
| Comparison | Pathway Name | Match Status | p Value | Impact |
|---|---|---|---|---|
| Menopause vs. Control | 1. Arginine & Proline Metabolism | 9/38 | 0.094 | 0.30 |
| 2. Amino Sugar & Nucleotide Sugar Metabolism | 1/37 | 0.100 | 0.0 | |
| 3. Phenylalanine Metabolism | 3/12 | 0.110 | 0.62 | |
| HDM vs. Control | 1. Arginine & Proline Metabolism | 9/38 | 0.007 | 0.30 |
| 2. Amino Sugar & Nucleotide Sugar Metabolism | 1/37 | 0.041 | 0.0 | |
| 3. Glycolysis/Gluconeogenesis | 1/26 | 0.059 | 0.0 | |
| Menopause HDM vs. Menopause | 1. Fatty Acid Elongation | 1/39 | 0.003 | 0.0 |
| 2. Fatty Acid Degradation | 1/39 | 0.003 | 0.0 | |
| 3. Purine Metabolism | 1/66 | 0.013 | 0.01 | |
| 4. Pyrimidine Metabolism | 1/39 | 0.023 | 0.01 | |
| 5. Pentose Phosphate Pathway | 3/22 | 0.024 | 0.17 | |
| 6. Biosynthesis of Unsaturated Fatty Acids | 2/36 | 0.041 | 0.0 | |
| Menopause HDM vs. HDM | 1. Pyrimidine Metabolism | 1/39 | 0.015 | 0.01 |
| 2. Pentose Phosphate Pathway | 3/22 | 0.025 | 0.17 | |
| 3. Nitrogen Metabolism | 1/6 | 0.033 | 0.0 | |
| 4. D-Glutamine & D-Glutamate metabolism | 2/6 | 0.035 | 0.5 | |
| 5. Porphyrin and Chlorophyll Metabolism | 3/30 | 0.035 | 0.03 | |
| 6. Arginine and Proline Metabolism | 9/38 | 0.045 | 0.30 | |
| 7. Butanoate Metabolism | 5/15 | 0.045 | 0.03 | |
| 8. Purine Metabolism | 1/66 | 0.047 | 0.01 |
| Comparison | Pathway Name | Match Status | p Value | Impact |
|---|---|---|---|---|
| Menopause vs. Control | 1. Glutathione Metabolism | 7/28 | 0.0005 | 0.43 |
| 2. Fatty Acid Elongation | 1/39 | 0.004 | 0.0 | |
| 3. Fatty Acid Degradation | 1/39 | 0.004 | 0.0 | |
| 4. Fatty Acid Biosynthesis | 5/47 | 0.006 | 0.02 | |
| 5. Glycine, serine, & Threonine Metabolism | 11/34 | 0.007 | 0.54 | |
| 6. Glycerophospholipid Metabolism | 2/36 | 0.009 | 0.03 | |
| 7. Pentose & Glucuronate Interconversions | 5/18 | 0.011 | 0.38 | |
| 8. Porphyrin and Chlorophyll Metabolism | 3/30 | 0.014 | 0.03 | |
| 9. Pentose Phosphate Pathway | 3/22 | 0.014 | 0.17 | |
| 10. Alanine, aspartate, & Glutamate Metabolism | 9/28 | 0.019 | 0.59 | |
| 11. Vitamin B6 Metabolism | 1/9 | 0.024 | 0.0 | |
| 12. Butanoate Metabolism | 5/15 | 0.033 | 0.0 | |
| 13. D-Glutamine & D-Glutamate metabolism | 3/6 | 0.036 | 0.5 | |
| 14. Beta-Alanine Metabolism | 3/21 | 0.038 | 0.06 | |
| 15. Pyrimidine Metabolism | 5/39 | 0.038 | 0.09 | |
| 16. Tryptophan Metabolism | 4/41 | 0.050 | 0.25 | |
| HDM vs. Control | 1. Glycerophospholipid Metabolism | 2/36 | 0.023 | 0.03 |
| 2. Selenocompound Metabolism | 1/20 | 0.024 | 0.0 | |
| 3. Histidine Metabolism | 9/16 | 0.026 | 0.71 | |
| 4. Glycine, Serine, & Threonine Metabolism | 11/34 | 0.041 | 0.54 | |
| Menopause HDM vs. Menopause | 1. Glycerophospholipid metabolism | 2/36 | 0.033 | 0.03 |
| 2. Arachidonic Acid Metabolism | 1/36 | 0.035 | 0.0 | |
| 3. Pyrimidine Metabolism | 5/39 | 0.042 | 0.09 | |
| 4. Selenocompound Metabolism | 1/20 | 0.045 | 0.0 | |
| Menopause HDM vs. HDM | 1. Arginine and Proline Metabolism | 8/38 | 0.015 | 0.38 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pederson, W.P.; Ellerman, L.M.; Jin, Y.; Gu, H.; Ledford, J.G. Metabolomic Profiling in Mouse Model of Menopause-Associated Asthma. Metabolites 2023, 13, 546. https://doi.org/10.3390/metabo13040546
Pederson WP, Ellerman LM, Jin Y, Gu H, Ledford JG. Metabolomic Profiling in Mouse Model of Menopause-Associated Asthma. Metabolites. 2023; 13(4):546. https://doi.org/10.3390/metabo13040546
Chicago/Turabian StylePederson, William P., Laurie M. Ellerman, Yan Jin, Haiwei Gu, and Julie G. Ledford. 2023. "Metabolomic Profiling in Mouse Model of Menopause-Associated Asthma" Metabolites 13, no. 4: 546. https://doi.org/10.3390/metabo13040546
APA StylePederson, W. P., Ellerman, L. M., Jin, Y., Gu, H., & Ledford, J. G. (2023). Metabolomic Profiling in Mouse Model of Menopause-Associated Asthma. Metabolites, 13(4), 546. https://doi.org/10.3390/metabo13040546