

Impact of Maternal Weight Gain on the Newborn Metabolome

 ,
,  , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Population: Clinical and Biochemical Measurements
- (A)
- From mothers, age, height, and weight measurements prior to gestation as recorded in their pregnancy booklet by the midwife, and weight at the end of gestation were collected. The GWG was calculated and its conformity with the 2009 recommendations of the Institute of Medicine (IOM) was verified [11]. Regarding the evolution of the pregnancy, the presence or absence of gestational diabetes, the degree of maternal adherence to the Mediterranean diet (AMD) using the validated questionnaire developed by Trichopoulos et al. [12] and smoking habits were recorded. Finally, the type of delivery was also recorded (vaginal or cesarean section).
- (B)
- From all newborns, the following information was recorded:
- -
- Somatometry (weight, length, head circumference and ponderal index and percentiles according to Intergrowth-21st), performed by trained nurses in the maternity ward. Weight was measured with ADE scale model M112600 (ADE GmbH & Co, Hamburg, Germany). Length was measured in the supine position using a neonatometer. Head circumference was measured with a tape measure at the maximum circumference. The ponderal index, also known as the corpulence index or Rohrer’s index, was calculated with the following formula PI = weight/length3 × 100. Newborns were classified as small for gestational age (SGA), appropriate for gestational age (AGA) or large for gestational age (LGA) [13].
- -
- Blood pressure (BP) was obtained by taking 3 measurements, using oscillometric method, with System 7100 Non-invasive Blood Pressure AMI (Advanced Medical Instruments Inc., Broken Arrow, OK, USA) and heart rate (HR).
- -
- Type of feeding (breastfeeding or infant formula feeding); as well as the need for supplementation with artificial formula during the first days of life in those who were breastfed.
- -
- Umbilical cord blood samples were obtained from the clamped umbilical cord immediately after delivery for metabolomic studies.
- (C)
- Blood sample were collected at 6 months of age to conduct metabolomic study.
- (D)
- In infants, at 12 months of life, weight, length, head circumference and BMI were recorded, and their percentiles were calculated according to the WHO 2006/2007 curves [14]. The type of feeding at 12 months was recorded. Finally, blood samples were collected to perform metabolite and biochemical studies.
2.2. NMR Metabolomics
2.3. Statistical Analysis
Demographics and Clinical Data Comparison
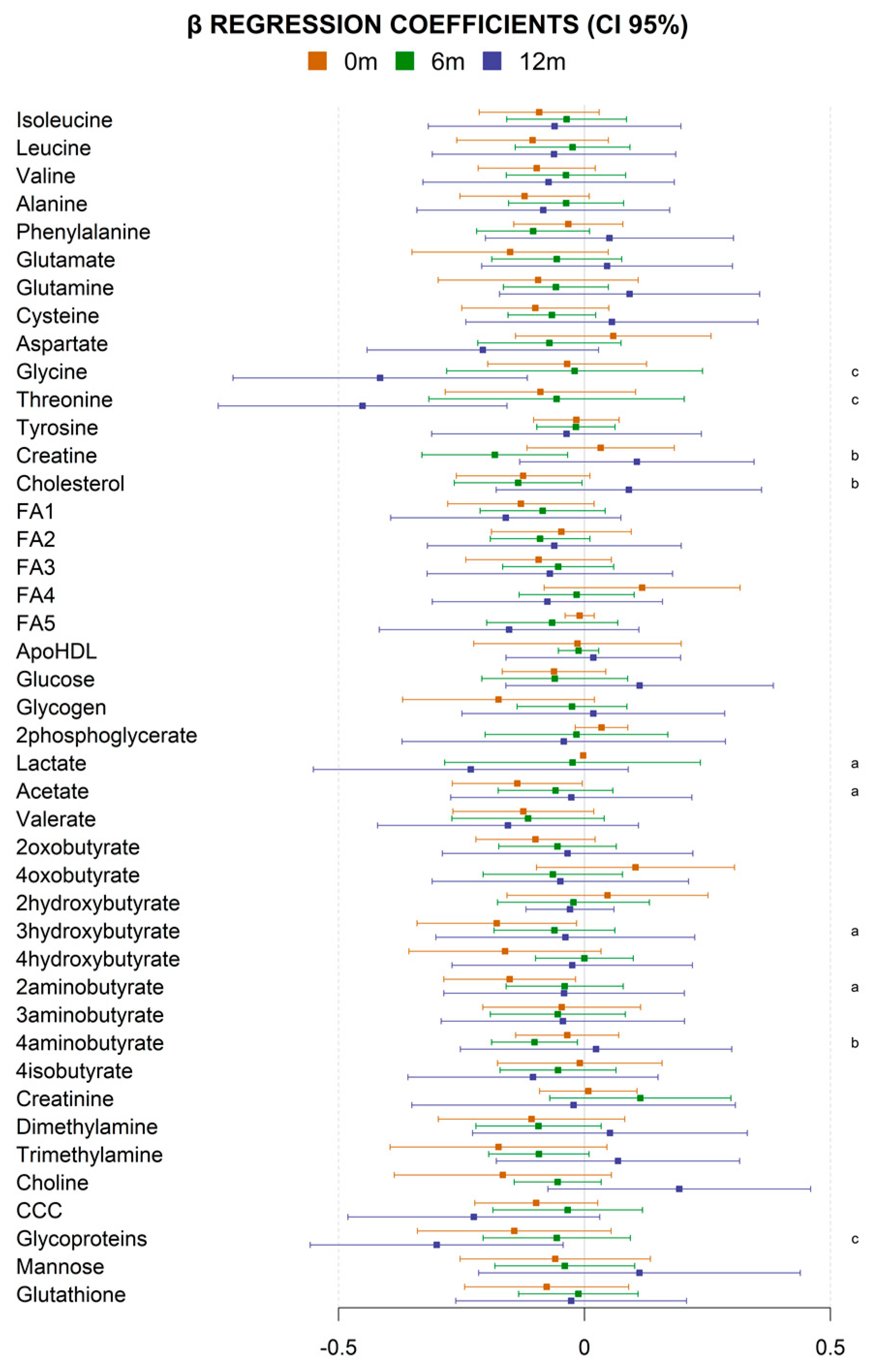
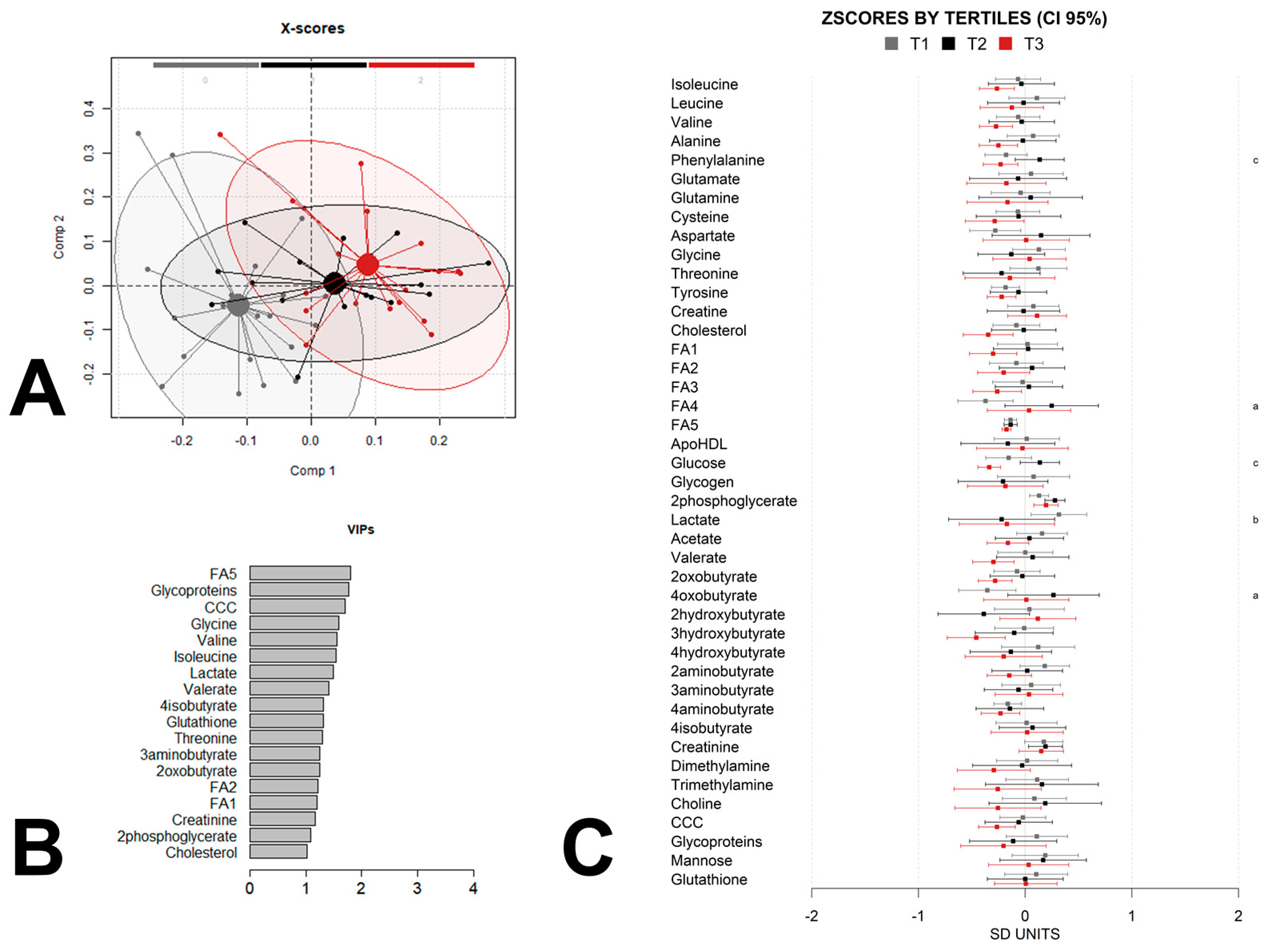
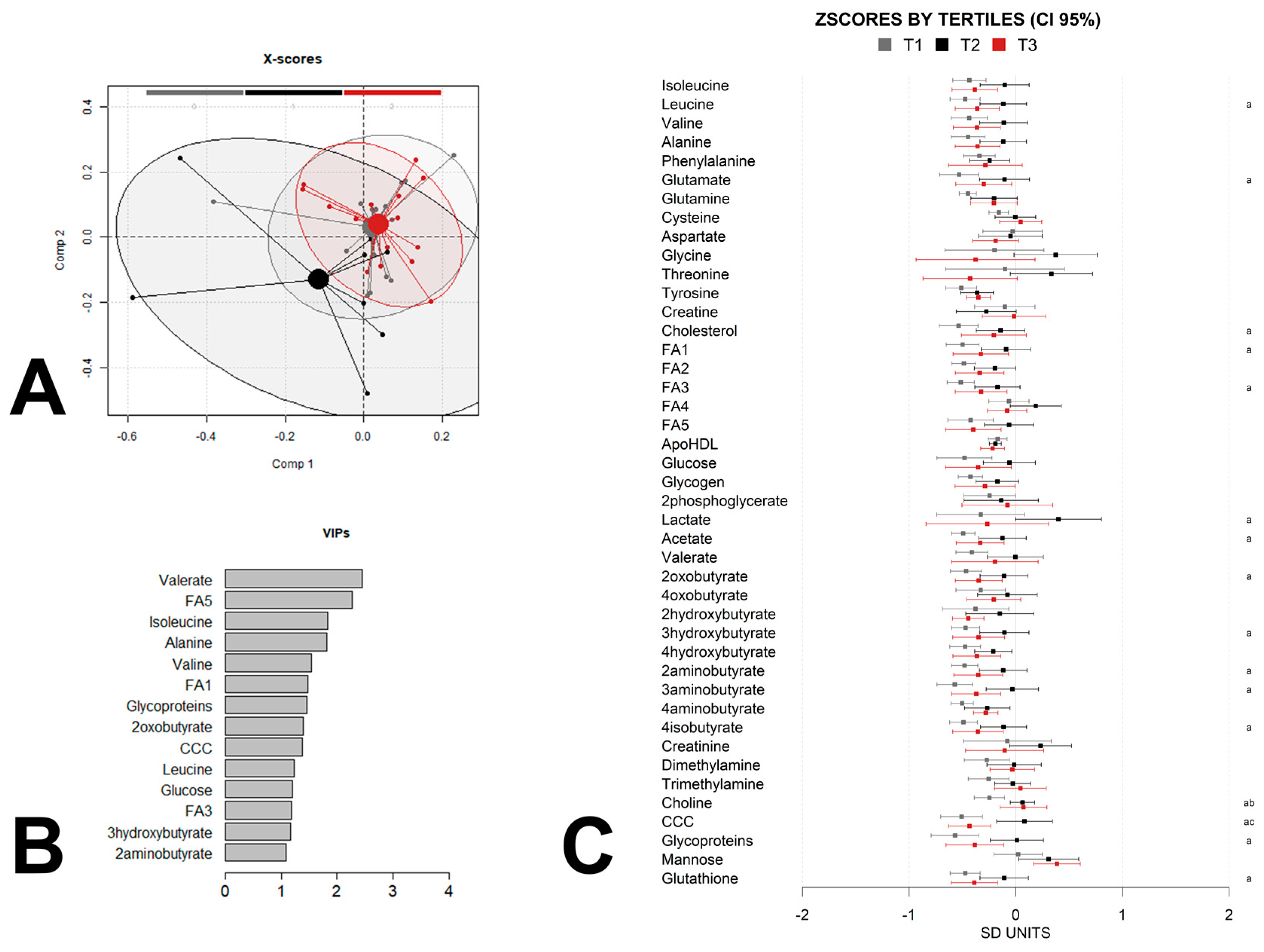
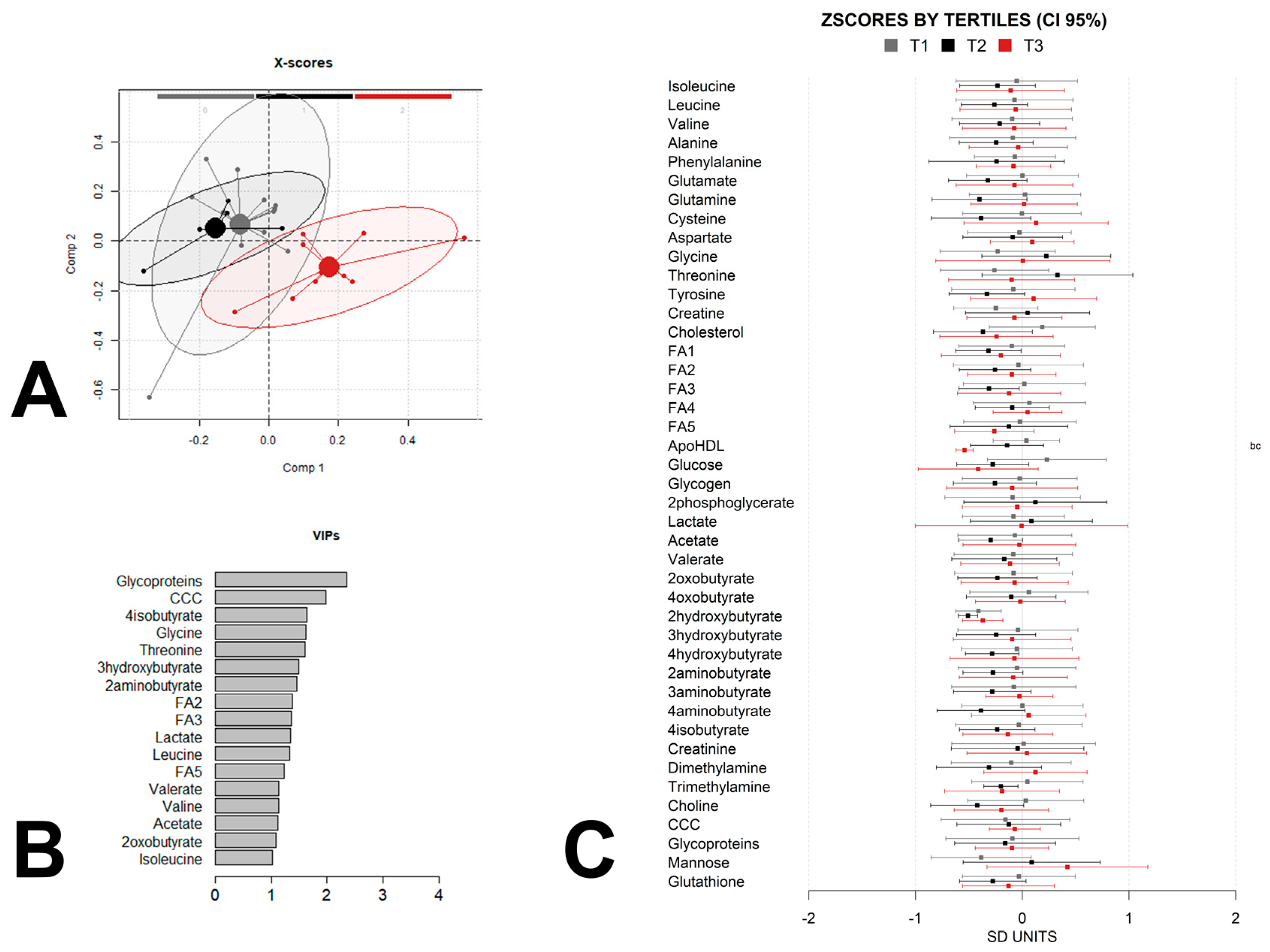
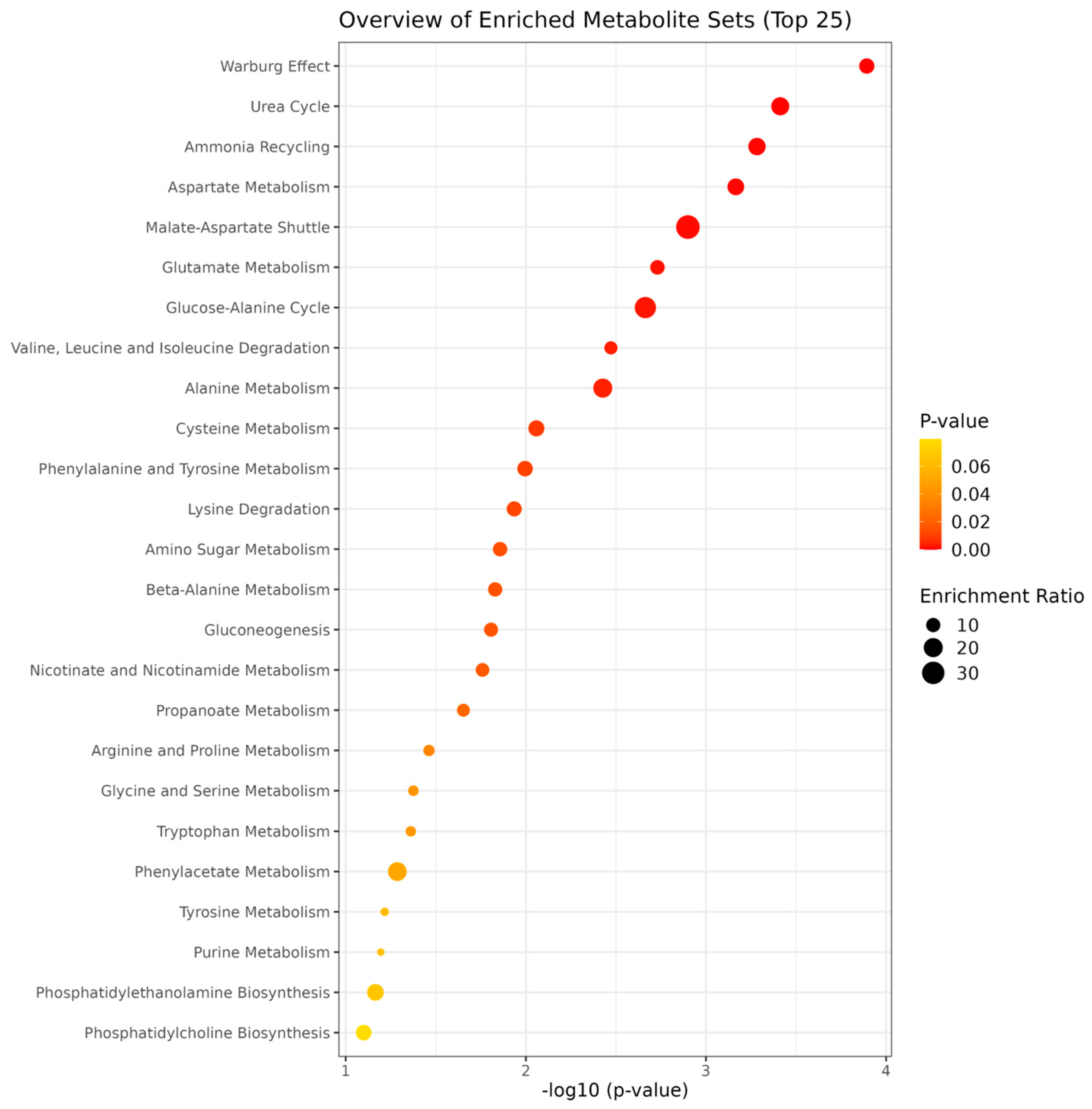
3. Results
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Barker, D.J.; Osmond, C.; Golding, G.; Kuh, D. Wadsworth ME Growth in utero, blood presure in childhood and adult life, and mortality from cardiovascular disease. BMJ 1989, 294, 564–567. [Google Scholar] [CrossRef] [PubMed]
- Kislal, S.; Shook, L.L.; Edlow, A.G. Perinatal exposure to maternal obesity: Lasting cardiometabolic impact on offspring. Prenat. Diagn. 2020, 40, 1109–1125. [Google Scholar] [CrossRef]
- Perng, W.; Oken, E.; Dabelea, D. Developmental overnutrition and obesity and type 2 diabetes in offspring. Diabetologia 2019, 62, 1779–1788. [Google Scholar] [CrossRef] [PubMed]
- Perng, W.; Gillman, M.W.; Fleisch, A.F.; Michalek, R.D.; Watkins, S.M.; Isganaitis, E.; Patti, M.; Oken, E. Metabolomic profiles and childhood obesity. Obesity (Silver Spring Md.) 2014, 22, 2570–2578. [Google Scholar] [CrossRef]
- Perng, W.; Rifas-Shiman, S.L.; McCulloch, S.; Chatzi, L.; Mantzoros, C.; Hivert, M.; Oken, E. Associations of cord blood metabolites with perinatal characteristics, newborn anthropometry, and cord blood hormones in project viva. Metab. Clin. Exp. 2017, 76, 11–22. [Google Scholar] [CrossRef]
- Kadakia, R.; Talbot, O.; Kuang, A.; Bain, J.R.; Muehlbauer, M.J.; Stevens, R.D.; Ilkayeva, O.R.; Lowe, L.P.; Metzger, B.E.; Newgard, C.B.; et al. Cord Blood Metabolomics: Association With Newborn Anthropometrics and C-Peptide Across Ancestries. J. Clin. Endocrinol. Metab. 2019, 104, 4459–4472. [Google Scholar] [CrossRef]
- Cao, T.; Zhao, J.; Hong, X.; Wang, G.; Hu, F.B.; Wang, X.; Liang, L. Cord Blood Metabolome and BMI Trajectory from Birth to Adolescence: A Prospective Birth Cohort Study on Early Life Biomarkers of Persistent Obesity. Metabolites 2021, 11, 739. [Google Scholar] [CrossRef] [PubMed]
- Lowe, J.; William, L.; Bain, J.R.; Nodzenski, M.; Reisetter, A.C.; Muehlbauer, M.J.; Stevens, R.D.; Ilkayeva, O.R.; Lowe, L.P.; Metzger, B.E.; et al. Erratum. Maternal BMI and Glycemia Impact the Fetal Metabolome. Diabetes Care 2017;40:902–910. Diabetes Care 2018, 41, 640. [Google Scholar] [CrossRef]
- Francis, E.C.; Kechris, K.; Cohen, C.C.; Michelotti, G.; Dabelea, D.; Perng, W. Metabolomic Profiles in Childhood and Adolescence Are Associated with Fetal Overnutrition. Metabolites 2022, 12, 265. [Google Scholar] [CrossRef]
- Ballard, J.L.; Novak, K.K.; Driver, M. A simplified score for assessment of fetal maturation of newly born infants. J. Pediatr. 1979, 95, 769–774. [Google Scholar] [CrossRef]
- Rasmussen, K.M.; Yaktine, A.L. Weight Gain During Pregnancy: Reexamining the Guidelines; National Academy of Sciences: Washington, DC, USA, 2009. [Google Scholar]
- Trichopoulou, A.; Costacou, T.; Bamia, C.; Trichopoulos, D. Adherence to a Mediterranean diet and survival in a Greek population. N. Engl. J. Med. 2003, 348, 2599–2608. [Google Scholar]
- Battaglia, F.C.; Lubchenco, L.O. A practical classification of newborn infants by weight and gestational age. J. Pediatr. 1967, 71, 159–163. [Google Scholar] [CrossRef] [PubMed]
- De Onis, M. WHO Child Growth Standards—Length/Height-for-Age, Weight-for-Age, Weight-for-Length, Weight-for-Height and Body Mass Index-for Age; World Health Organization: Geneva, Switzerland, 2006; Available online: https://www.who.int/publications/i/item/924154693X (accessed on 15 April 2022).
- Pichler, G.; Amigo, N.; Tellez-Plaza, M.; Pardo-Cea, M.A.; Dominguez-Lucas, A.; Marrachelli, V.G.; Monleon, D.; Martin-Escudero, J.C.; Ascaso, J.F.; Chaves, F.J.; et al. LDL particle size and composition and incident cardiovascular disease in a South-European population: The Hortega-Liposcale Follow-up Study. Int. J. Cardiol. 2018, 264, 172–178. [Google Scholar] [CrossRef] [PubMed]
- Galbis-Estrada, C.; Pinazo-Durán, M.D.; Martínez-Castillo, S.; Morales, J.M.; Monleón, D.; Zanon-Moreno, V. A metabolomic approach to dry eye disorders. The role of oral supplements with antioxidants and omega 3 fatty acids. Mol. Vis. 2015, 21, 555–567. [Google Scholar] [PubMed]
- García-Villaescusa, A.; Morales-Tatay, J.M.; Monleón-Salvadó, D.; González-Darder, J.M.; Bellot-Arcis, C.; Montiel-Company, J.M.; Almerich-Silla, J.M. Using NMR in saliva to identify possible biomarkers of glioblastoma and chronic periodontitis. PLoS ONE 2018, 13, e0188710. [Google Scholar] [CrossRef]
- Trygg, J.; Wold, S. Orthogonal projections to latent structures (O-PLS). J. Chemom. 2002, 16, 119–128. [Google Scholar] [CrossRef]
- Bhatia, J.; Gates, A. The Newborn Infant. In Clinical Biochemistry: Metabolic and Clinical Aspects; Elsevier: Amsterdam, The Netherlands, 2015; pp. 701–718. [Google Scholar]
- He, X.; Ji, Y.; Li, H. Maternal gestational weight gain and offspring’s amino acid metabolome in the first year of life: A prospective birth cohort study. JAMA Pediatr. 2020, 6, 570–578. [Google Scholar]
- Lu, Q.; Cheng, Y.; Chen, L.; Chen, X.; Wang, X.; Wang, S.; Yan, W. Associations of maternal gestational weight gain with newborn lipid levels and the risk of macrosomia: A prospective cohort study. BMC Pregnancy Childbirth 2021, 1, 1–9. [Google Scholar] [CrossRef]
- Newman, J.C.; Verdin, E. β-hydroxybutyrate: Much more than a metabolite. Diabetes Res. Clin. Pract. 2014, 106, 173–181. [Google Scholar] [CrossRef]
- Newman, J.C.; Verdin, E. Ketone bodies as signaling metabolites. Trends Endocrinol. Metab. 2014, 25, 42–52. [Google Scholar] [CrossRef]
- Harreiter, J.; Simmons, D.; Desoye, G.; Corcoy, R. Adverse pregancy outcomes and metabolic syndrome in women with gestational diabetes treated with metformin or insulin: A randomised trial. Diabetologia 2019, 4, 601–612. [Google Scholar] [CrossRef]
- Troisi, J.; Symes, S.J.K.; Lombardi, M.; Cavallo, P.; Colucci, A.; Scala, G.; Adair, D.C.; Guida, M.; Richards, S.M. Placental Metabolomics of Fetal Growth Restriction. Metabolites 2023, 13, 235. [Google Scholar] [CrossRef]
- Rising, C.L.; D’Alecy, L.G. Hypoxia-induced increases in hypoxic tolerance augmented by beta- hydroxybutyrate in mice. Stroke 1989, 20, 1219–1225. [Google Scholar] [CrossRef]
- Tremaroli, V.; Bäckhed, F. Functional interactions between the gut microbiota and host metabolism. Nature 2012, 489, 242–249. [Google Scholar] [CrossRef] [PubMed]
- Masuyama, H.; Hiramatsu, Y. Relationships of maternal weight gain during pregnancy with pacental and fetal weight, placental and fetal ratio and fetal lactate levels in normal and abnormal pregnancy. J. Obstet. Gynaecol. Res. 2010, 3, 457–465. [Google Scholar] [CrossRef]
- Goldin, K.R. Maternal and infant serum phenylalanine levels and pregnancy outcome in infants of nonphenylketonuric mothers. J. Natl. Med. Assoc. 1993, 85, 617–620. [Google Scholar]
- Zhang, Z.; Monleon, D.; Verhamme, P.; Staessen, J.A. Branched-Chain Amino Acids as Critical Switches in Health and Disease. Hypertension 2018, 72, 1012–1022. [Google Scholar] [CrossRef]
- Zeisel, S.H.; Da Costa, K. Choline: An essential nutrient for public health. Nutr. Rev. 2009, 67, 615–623. [Google Scholar] [CrossRef]
- Tang, W.H.W.; Wang, Z.; Levison, B.S.; Koeth, R.A.; Britt, E.B.; Fu, X.; Wu, Y.; Hazen, S.L. Intestinal Microbial Metabolism of Phosphatidylcholine and Cardiovascular Risk. N. Engl. J. Med. 2013, 368, 1575–1584. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Mothers’ Variables | 1st Tertile GWG (N = 28) | 2nd Tertile GWG (N = 27) | 3rd Tertile GWG (N = 28) | p Value |
|---|---|---|---|---|
| Age (years) ¥ | 31.5 (± 8.48) | 34.11 (± 5.17) | 33.18 (± 5.41) | 0.429 |
| Maternal obesity | Yes 75% No 25% | Yes 48.1% No 51.9% | Yes 57.1% No 42.9% | 0.116 |
| BMI prior to pregnancy (Kg/m²) ¥ | 33.61 (±7.31) | 28.34 (± 7.07) | 28.07 (± 6.03) | 0.004 ** |
| GWG (Kg) ǂ | 2.33 (± 4.31) | 10.11 (± 1.12) | 16.78 (± 3.91) | <0.001 *** |
| GWG adequacy according to IOM recommendations • | Yes 100% No 0% | Yes 66.7% No 33.3% | Yes 21.4% No 78.6% | <0.001 *** |
| AMD score ¥ | 6.96 (± 2) | 7.76 (± 1.7) | 7.46 (± 1.6) | 0.396 |
| Degree of AMD • | High 12.5% Medium 58.3% Low 29.2% | High 23.5% Medium 70.6% Low 5.9% | High 8.3% Medium 75% Low 16.7% | 0.266 |
| Diabetes during pregnancy | Yes 35.7% No 64.3% | Yes 11.1% No 88.9% | Yes 17.9% No 82.1% | 0.072 |
| Smoking habits • | Yes 14.3% No 85.7% | Yes 18.5% No 81.5% | Yes 21.4% No 78.6% | 0.78 |
| Type of delivery • | Vaginal 50% Cesarean 50% | Vaginal 44.4% Cesarean 55.6% | Vaginal 60.7% Cesarean 39.3% | 0.469 |
| Newborns’ Variables | 1st Tertile GWG (N = 28) | 2nd Tertile GWG (N = 27) | 3rd Tertile GWG (N = 28) | p Value |
|---|---|---|---|---|
| Birth weight (BW) (g) ǂ | 3363 (±552) | 3311 (±557) | 3454 (±449) | 0.593 |
| Weight percentile ǂ | 53.45 (±31.14) | 52.4 (±31.1) | 58 (±30.4) | 0.773 |
| BW Classification • | SGA 7.1% AGA 75% LGA 17.9% | SGA 11.1% AGA 70.4% LGA 18.5% | SGA 10.7% AGA 60.7% LGA 28.6% | 0.8 |
| Length (cm) ǂ | 49.37 (±2.08) | 48.6(±1.94) | 49.67 (±2.13) | 0.149 |
| Length percentile ǂ | 48.81 (±28.2) | 39.78 (±25.71) | 53.33 (±31.8) | 0.21 |
| Head circumference (HC) (cm) ¥ | 34.46 (±1.52) | 34.38 (±1.57) | 34.97 (±1.41) | 0.351 |
| HC percentile¥ | 62.8 (±27.01) | 64 (±28.79) | 71.68 (±27.67) | 0.323 |
| ponderal index ¥ | 2.77 (±0.267) | 2.85 (±0.24) | 2.81 (±0.31) | 0.406 |
| Systolic blood pressure (mmHg) ǂ | 78.63 (±87) | 81.5 (±12.5) | 78.89 (±12.66) | 0.605 |
| Diastolic blood pressure (mmHg) ǂ | 50.66 (±10) | 49.79 (±9.63) | 49.5 (±9.45) | 0.899 |
| Heart rate (bpm) ǂ | 126.81 (±14.69) | 129.65 (±12.65) | 132.69 (±16.93) | 0.366 |
| Type of feeding • | breast 67.9% formula 32.1% | breast 88.9% formula 11.1% | breast 75% formula 25% | 0.169 |
| Formula Supplementation • | Yes 31.6% No 68.4% | Yes 29.2% No 70.8% | Yes 19% No 81% | 0.62 |
| 1st Tertile GWG | 2nd Tertile GWG | 3rd Tertile GWG | p Value | |
|---|---|---|---|---|
| 12 Month Somatometry | (N = 27) | (N = 24) | (N = 25) | |
| Weight (g) ǂ | 10,509 (±1516) | 9736 (±1193) | 10,293 (±1242) | 0.114 |
| Weight percentile ¥ | 71.14 (±28.75) | 58.54 (± 29.43) | 61.92 (± 27.87) | 0.214 |
| Length (cm) ǂ | 76.74 (±3.4) | 74.83 (±2.64) | 76.64 (±2.4) | 0.039 * |
| Length percentile ¥ | 60.4 (±32) | 48.79 (±28.72) | 60.08 (±26.68) | 0.128 |
| Head circumference (HC) (cm) ¥ | 46.38 (±1.32) | 45.81 (±1.15) | 46.80 (±1.69) | 0.094 |
| HC percentile ¥ | 64.64 (±27.89) | 57.83 (±24.94) | 68.2 (±29.1) | 0.216 |
| BMI (Kg/m²) ǂ | 17.65 (±1.53) | 17.29 (±1.76) | 17.38 (±1.69) | 0.722 |
| BMI percentile ¥ | 67.86 (±29.172.63) | 6(±28.24) | 60.07(±29.59) | 0.516 |
| 12 Month Blood Test | (N = 12) | (N = 8) | (N = 12) | |
| Leptin (pg/ml) ¥ | 452 (±613) | 691 (±380) | 513 (±626) | 0.378 |
| Total cholesterol (mg/dl) ǂ | 160.6 (±32) | 158 (±38.2) | 152.75 (±25.64) | 0.827 |
| HDL cholesterol (mg/dl) ǂ | 40.5 (±9.4) | 44.75 (±10.91) | 43.16 (±10.66) | 0.64 |
| LDL cholesterol (mg/dl) ǂ | 95.75(±34.6) | 100.2 (±33.8) | 96.5 (±20.5) | 0.94 |
| Triglycerides (mg/dl) ǂ | 183 (±90) | 173 (±90.7) | 119 (±45) | 0.115 |
| HOMA index ¥ | 0.955 (±0.77) | 1.99 (±1.58) | 1.34 (±1.15) | 0.126 |
| Uric acid (mg/dl) ǂ | 3.15 (±0.64) | 3.22 (±0.49) | 3.05 (±0.669) | 0.822 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guixeres-Esteve, T.; Ponce-Zanón, F.; Morales, J.M.; Lurbe, E.; Alvarez-Pitti, J.; Monleón, D. Impact of Maternal Weight Gain on the Newborn Metabolome. Metabolites 2023, 13, 561. https://doi.org/10.3390/metabo13040561
Guixeres-Esteve T, Ponce-Zanón F, Morales JM, Lurbe E, Alvarez-Pitti J, Monleón D. Impact of Maternal Weight Gain on the Newborn Metabolome. Metabolites. 2023; 13(4):561. https://doi.org/10.3390/metabo13040561
Chicago/Turabian StyleGuixeres-Esteve, Teresa, Francisco Ponce-Zanón, José Manuel Morales, Empar Lurbe, Julio Alvarez-Pitti, and Daniel Monleón. 2023. "Impact of Maternal Weight Gain on the Newborn Metabolome" Metabolites 13, no. 4: 561. https://doi.org/10.3390/metabo13040561
APA StyleGuixeres-Esteve, T., Ponce-Zanón, F., Morales, J. M., Lurbe, E., Alvarez-Pitti, J., & Monleón, D. (2023). Impact of Maternal Weight Gain on the Newborn Metabolome. Metabolites, 13(4), 561. https://doi.org/10.3390/metabo13040561