
Physical Performance and Skeletal Muscle Transcriptional Adaptations Are Not Impacted by Exercise Training Frequency in Mice with Lower Extremity Peripheral Artery Disease
 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Material and Methods
2.1. Mouse Model of Lower Extremity Peripheral Artery Disease
2.2. Exercise Training Protocols
2.3. Treadmill Exhaustion Test
2.4. Gene Expression Analysis by Real-Time Polymerase Chain Reaction
2.5. Statistical Analysis
3. Results
3.1. Physical Performance
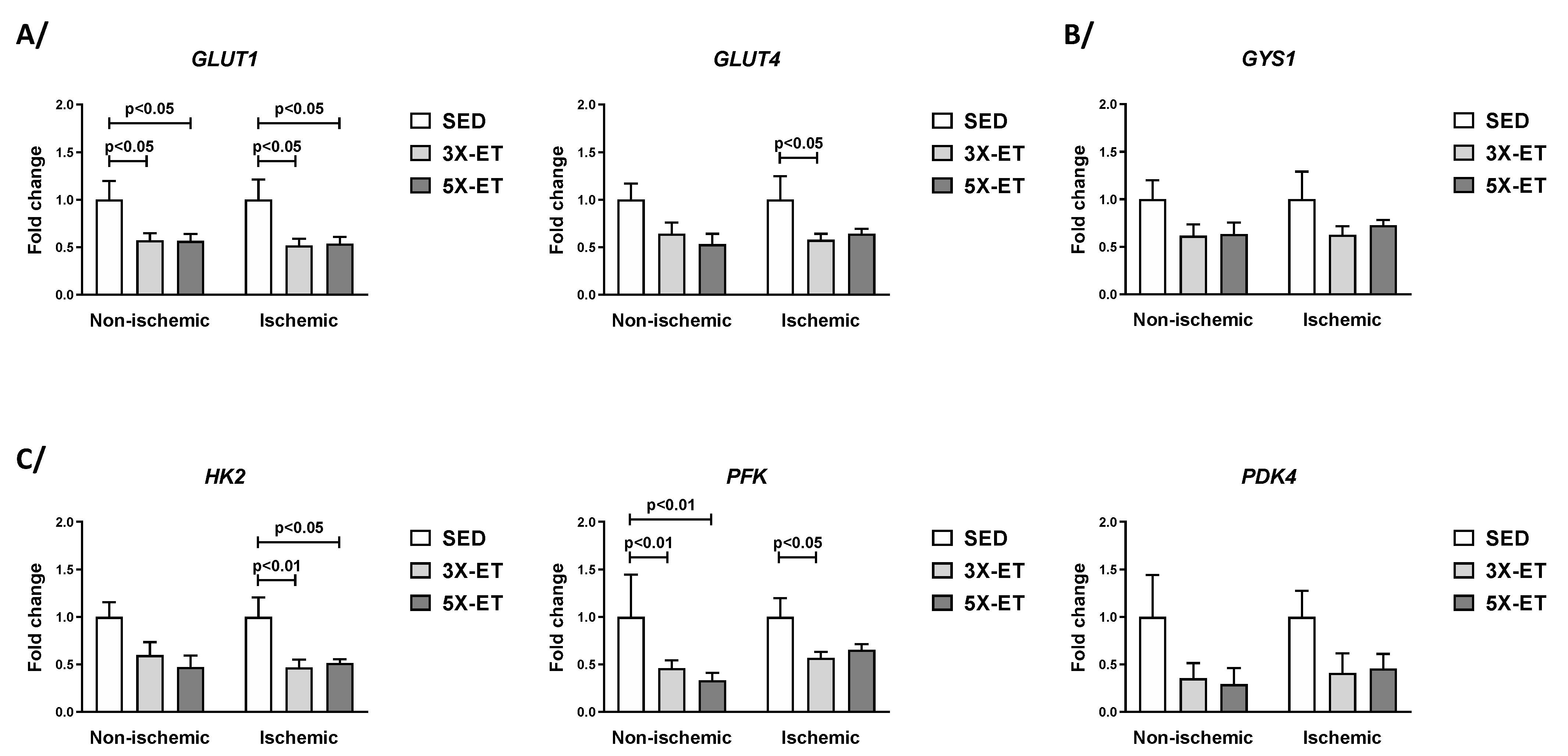
3.2. Gene Expression Related to Glucose and Lactate Metabolism
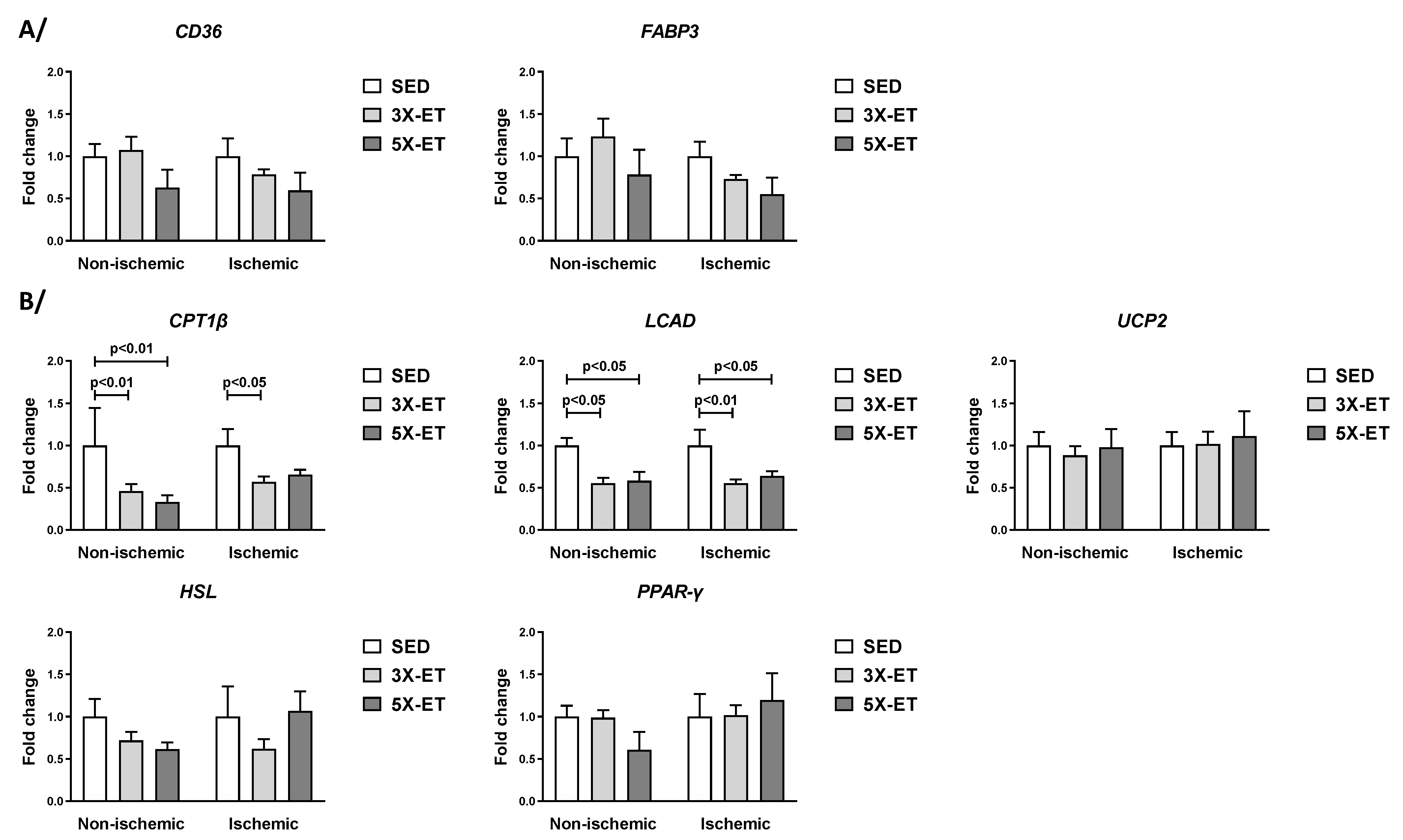
3.3. Gene Expression Related to Lipid Metabolism
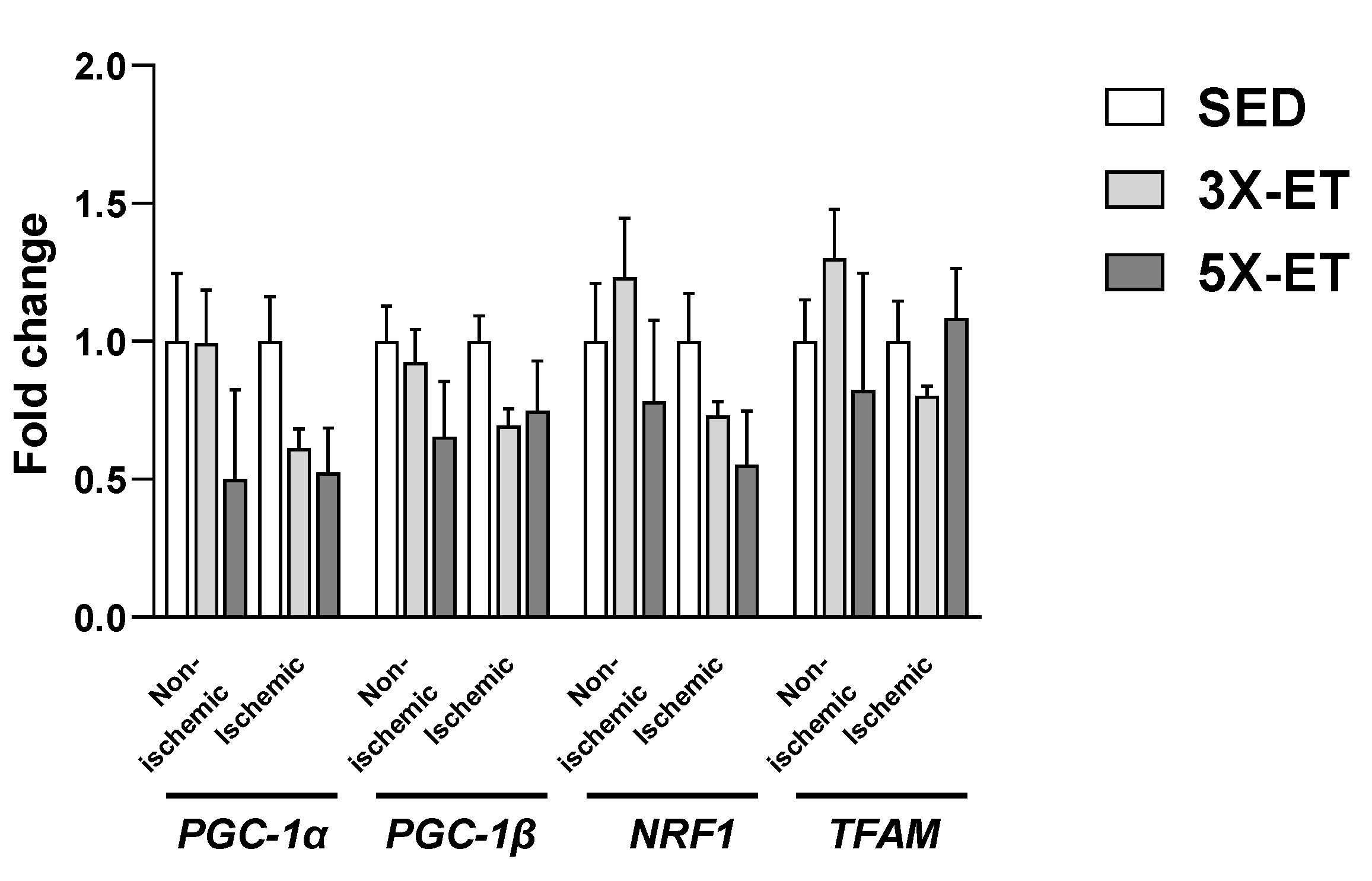
3.4. Gene Expression Related to Mitochondrial Biogenesis
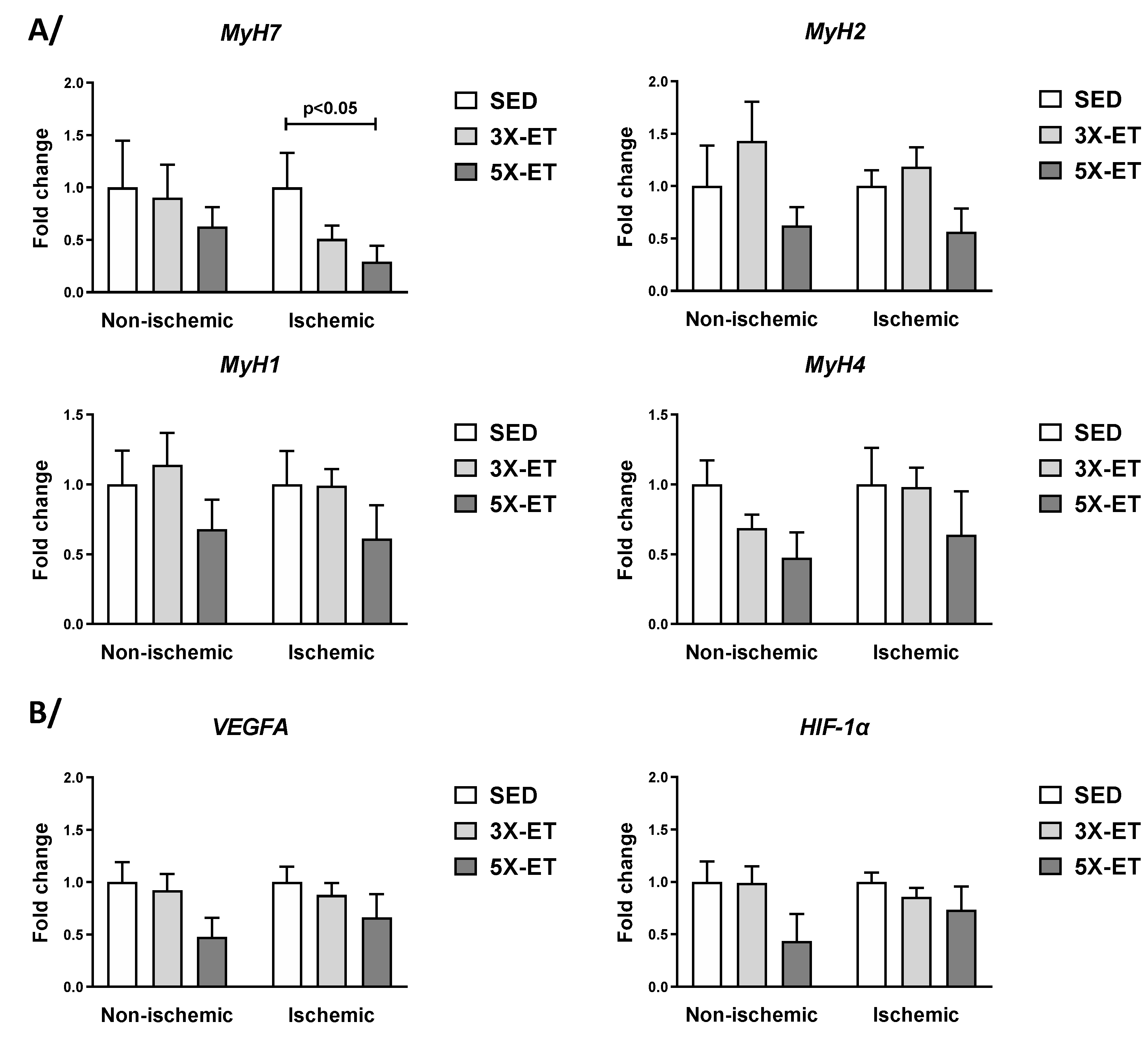
3.5. Gene Expression Related to Muscle Fiber-Type and Angiogenesis
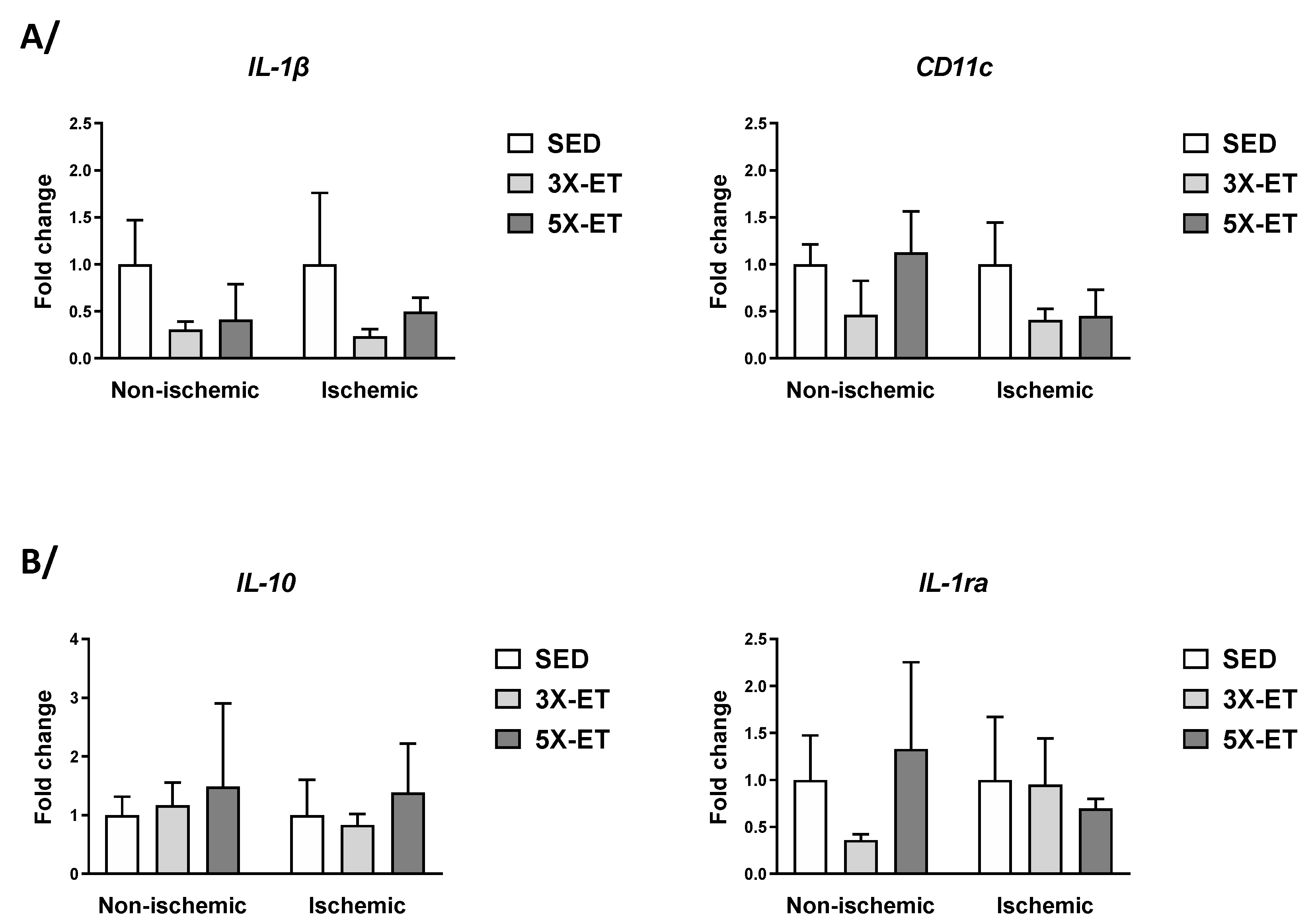
3.6. Gene Expression Related to Inflammation
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Song, P.; Rudan, D.; Zhu, Y.; Fowked, F.J.I.; Rahimi, K.; Fowkes, F.G.R.; Rudan, I. Global, regional, and national prevalence and risk factors for peripheral artery disease in 2015: An updated systematic review and analysis. Lancet Glob. Health 2019, 7, e1020–e1030. [Google Scholar] [CrossRef] [PubMed]
- Conte, S.M.; Vale, P.R. Peripheral Arterial Disease. Heart Lung Circ. 2018, 27, 427–432. [Google Scholar] [CrossRef]
- Aboyans, V.; Ricco, J.B.; Bartelink, M.E.L.; Bjorck, M.; Brodmann, M.; Cohnert, T.; Collet, J.P.; Czerny, M.; De Carlo, M.; Debus, S.; et al. 2017 ESC Guidelines on the Diagnosis and Treatment of Peripheral Arterial Diseases, in collaboration with the European Society for Vascular Surgery (ESVS): Document covering atherosclerotic disease of extracranial carotid and vertebral, mesenteric, renal, upper and lower extremity arteries Endorsed by: The European Stroke Organization (ESO)The Task Force for the Diagnosis and Treatment of Peripheral Arterial Diseases of the European Society of Cardiology (ESC) and of the European Society for Vascular Surgery (ESVS). Eur. Heart J. 2018, 39, 763–816. [Google Scholar] [PubMed]
- Gerhard-Herman, M.D.; Gornik, H.L.; Barrett, C.; Barshes, N.R.; Corriere, M.A.; Drachman, D.E.; Fleisher, L.A.; Fowkes, F.G.; Hamburg, N.M.; Kinlay, S.; et al. 2016 AHA/ACC Guideline on the Management of Patients with Lower Extremity Peripheral Artery Disease: A Report of the American College of Cardiology/American Heart Association Task Force on Clinical Practice Guidelines. Circulation 2017, 135, e726–e779. [Google Scholar] [PubMed]
- Fakhry, F.; van de Luijtgaarden, K.M.; Bax, L.; den Hoed, P.T.; Hunink, M.G.; Rouwet, E.V.; Spronk, S. Supervised walking therapy in patients with intermittent claudication. J. Vasc. Surg. 2012, 56, 1132–1142. [Google Scholar] [CrossRef] [PubMed]
- Gardner, A.W.; Poehlman, E.Z. Exercise rehabilitation programs for the treatment of claudication pain. A meta-analysis. JAMA 1995, 274, 975–980. [Google Scholar] [CrossRef] [PubMed]
- Lane, R.; Harwood, A.; Watson, L.; Leng, G.C. Exercise for intermittent claudication. Cochrane Database Syst. Rev. 2017, 12, CD000990. [Google Scholar] [CrossRef] [PubMed]
- Parmenter, B.J.; Dieberg, G.; Smart, N.A. Exercise training for management of peripheral arterial disease: A systematic review and meta-analysis. Sport. Med. 2015, 45, 231–244. [Google Scholar] [CrossRef] [PubMed]
- Patel, K.; Polonsky, T.S.; Kibbe, M.R.; Guralnik, J.M.; Tian, L.; Ferrucci, L.; Criqui, M.H.; Sufit, R.; Leeuwenburgh, C.; Zhang, D.; et al. Clinical characteristics and response to supervised exercise therapy of people with lower extremity peripheral artery disease. J. Vasc. Surg. 2021, 73, 608–625. [Google Scholar] [CrossRef]
- Calanca, L.; Lanzi, S.; Ney, B.; Berchtold, A.; Mazzolai, L. Multimodal supervised exercise significantly improves walking performances without changing hemodynamic parameters in patients with symptomatic lower extremity peripheral artery disease. Vasc. Endovasc. Surg. 2020, 54, 605–611. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, E.; Silva, I. Supervised exercise therapy in intermittent claudication: A systematic review of clinical impact and limitations. Int. Angiol. 2020, 39, 60–75. [Google Scholar] [CrossRef] [PubMed]
- Treat-Jacobson, D.; McDermott, M.M.; Bronas, U.G.; Campia, U.; Collins, T.C.; Criqui, M.H.; Gardner, A.W.; Hiatt, W.R.; Regensteiner, J.G.; Rich, K. Optimal Exercise Programs for Patients With Peripheral Artery Disease: A Scientific Statement from the American Heart Association. Circulation 2019, 139, e10–e33. [Google Scholar] [CrossRef]
- Pellegrin, M.; Bouzourene, K.; Aubert, J.F.; Bielmann, C.; Gruetter, R.; Rosenblatt-Velin, N.; Poitry-Yamate, C.; Mazzolai, L. Impact of aerobic exercise type on blood flow, muscle energy metabolism, and mitochondrial biogenesis in experimental lower extremity artery disease. Sci. Rep. 2020, 10, 14048. [Google Scholar] [CrossRef] [PubMed]
- Krishna, S.M.; Omer, S.M.; Li, J.; Morton, S.K.; Jose, R.J.; Golledge, J. Development of a two-stage limb ischemia model to better simulate human peripheral artery disease. Sci. Rep. 2020, 10, 3449. [Google Scholar] [CrossRef]
- Nagase, H.; Yao, S.; Ikeda, S. Acute and chronic effects of exercise on mRNA expression in the skeletal muscle of two mouse models of peripheral artery disease. PLoS ONE 2017, 12, e0182456. [Google Scholar] [CrossRef]
- Burch, N.; Arnold, A.S.; Item, F.; Summermatter, S.; Brochmann Santana Santos, G.; Christe, M.; Boutellier, U.; Toigo, M.; Handschin, C. Electric pulse stimulation of cultured murine muscle cells reproduces gene expression changes of trained mouse muscle. PLoS ONE 2010, 5, e10970. [Google Scholar] [CrossRef]
- Albadawi, H.; Tzika, A.A.; Rask-Madsen, C.; Crowley, L.M.; Koulopoulos, M.W.; Yoo, H.J.; Watkins, M.T. Revascularization and muscle adaptation to limb demand ischemia in diet-induced obese mice. J. Surg. Res. 2016, 205, 49–58. [Google Scholar] [CrossRef] [PubMed]
- Durigan, J.L.Q.; Peviani, S.M.; Russo, T.L.; Duarte, A.C.; Vieira, R.P.; Martins, M.A.; Carvalho, C.R.F.; Salvini, T.F. Physical training leads to remodeling of diaphragm muscle in asthma model. Int. J. Sports Med. 2009, 30, 430–434. [Google Scholar] [CrossRef]
- Fiuza-Luces, C.; Santos-Lozano, A.; Llavero, F.; Campo, R.; Nogales-Gadea, G.; Diez-Bermejo, J.; Baladrón, C.; Gónzalez-Murillo, Á.; Arenas, J.; Martín, M.A.; et al. Muscle molecular adaptations to endurance exercise training are conditioned by glycogen availability: A proteomics-based analysis in the McArdle mouse model. J. Physiol. 2018, 596, 1035–1061. [Google Scholar] [CrossRef]
- Vieira, R.P.; de Andrade, V.F.; Duarte, A.C.; Dos Santos, A.B.; Mauad, T.; Martins, M.A.; Dolhnikoff, A.; Carvalho, R.F. Aerobic conditioning and allergic pulmonary inflammation in mice. II. Effects on lung vascular and parenchymal inflammation and remodeling. Am. J. Physiol. Lung Cell. Mol. Physiol. 2008, 295, 670–679. [Google Scholar] [CrossRef]
- Lavier, J.; Beaumann, M.; Menetrey, S.; Mazzolai, L.; Peyter, A.C.; Pellegrin, M.; Millet, G.P. Supramaximal intensity hypoxic exercise and vascular function assessment in mice. J. Vis. Exp. 2019, 145, e58708. [Google Scholar]
- McDermott, M.M. Exercise Rehabilitation for Peripheral Artery Disease: A Review. J. Cardiopulm. Rehabil. Prev. 2018, 38, 63–69. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, Y.; Tanaka, K.; Yabushita, N.; Sakai, T.; Shigematsu, R. Effects of exercise frequency on functional fitness in older adult women. Arch. Gerontol. Geriatr. 2007, 44, 163–173. [Google Scholar] [CrossRef]
- Murlasits, Z.; Reed, J.; Wells, K. Effect of resistance training frequency on physiological adaptations in older adults. J. Exerc. Sci. Fit. 2012, 10, 28–32. [Google Scholar] [CrossRef]
- Nicolai, S.P.; Hendriks, E.J.; Prins, M.H.; Teijink, J.A.; EXITPAD Study Group. Optimizing supervised exercise therapy for patients with intermittent claudication. J. Vasc. Surg. 2010, 52, 1226–1233. [Google Scholar] [CrossRef] [PubMed]
- Le Bris, S.; Ledermann, B.; Topin, N.; Messner-Pellenc, P.; Le Gallais, D. High versus low training frequency in cardiac rehabilitation using a systems model of training. Eur. J. Appl. Physiol. 2006, 96, 217–224. [Google Scholar] [CrossRef]
- Harwood, A.E.; Smith, G.E.; Cayton, T.; Broadbent, E.; Chetter, I.C. A Systematic Review of the Uptake and Adherence Rates to Supervised Exercise Programs in Patients with Intermittent Claudication. Ann. Vasc. Surg. 2016, 34, 280–289. [Google Scholar] [CrossRef]
- Silva, L.A. Effects of different frequencies of physical training on electron transport chain and oxidative damage in healthy mice. Motriz J. Phys. Educ. 2018, 24, e101804. [Google Scholar] [CrossRef]
- Brooks, G.A. The Science and Translation of Lactate Shuttle Theory. Cell Metab. 2018, 27, 757–785. [Google Scholar] [CrossRef]
- Gill, J.F.; Santos, G.; Schnyder, S.; Handschin, C. PGC-1alpha affects aging-related changes in muscle and motor function by modulating specific exercise-mediated changes in old mice. Aging Cell 2018, 17, e12697. [Google Scholar] [CrossRef]
- Casanova-Vallve, N.; Duglan, D.; Vaughan, M.E.; Pariollaud, M.; Handzlik, M.K.; Fan, W.; Yu, R.T.; Liddle, C.; Downes, M.; Delezie, J.; et al. Daily running enhances molecular and physiological circadian rhythms in skeletal muscle. Mol. Metab. 2022, 61, 101504. [Google Scholar] [CrossRef]
- Liang, X.; Liu, L.; Fu, T.; Zhou, Q.; Zhou, D.; Xiao, L.; Liu, J.; Kong, Y.; Xie, H.; Yi, F.; et al. Exercise Inducible Lactate Dehydrogenase B Regulates Mitochondrial Function in Skeletal Muscle. J. Biol. Chem. 2016, 291, 25306–25318. [Google Scholar] [CrossRef]
- Halestrap, A.P.; Wilson, M.C. The monocarboxylate transporter family—Role and regulation. IUBMB Life 2012, 64, 109–119. [Google Scholar] [CrossRef] [PubMed]
- Ahmadi, A.; Sheikholeslami-Vatani, D.; Ghaeeni, S.; Baazm, M. The effects of different training modalities on monocarboxylate transporters MCT1 and MCT4, hypoxia inducible factor-1α (HIF-1α), and PGC-1α gene expression in rat skeletal muscles. Mol. Biol. Rep. 2021, 48, 2153–2161. [Google Scholar] [CrossRef] [PubMed]
- Martinez-Huenchullan, S.F.; Ban, L.A.; Olaya-Agudo, L.F.; Maharjan, B.R.; Williams, P.F.; Tam, C.S.; McLennan, S.V.; Twigg, S.M. Constant-Moderate and High-Intensity Interval Training Have Differential Benefits on Insulin Sensitive Tissues in High-Fat Fed Mice. Front. Physiol. 2019, 10, 459. [Google Scholar] [CrossRef] [PubMed]
- Schmitt, A.; Haug, A.L.; Schlegel, F.; Fragasso, A.; Munz, B. Effects of 10 weeks of regular running exercise with and without parallel PDTC treatment on expression of genes encoding sarcomere-associated proteins in murine skeletal muscle. Cell Stress Chaperones 2018, 35, 1041–1054. [Google Scholar] [CrossRef]
- Deloux, R.; Tannous, C.; Ferry, A.; Li, Z.; Mericskay, M. Aged Nicotinamide Riboside Kinase 2 Deficient Mice Present an Altered Response to Endurance Exercise Training. Front. Physiol. 2018, 9, 1290. [Google Scholar] [CrossRef]
- Kivela, R.; Silvennoinen, M.; Touvra, A.M.; Lehti, T.M.; Kainulainen, H.; Vihko, V. Effects of experimental type 1 diabetes and exercise training on angiogenic gene expression and capillarization in skeletal muscle. FASEB J. 2006, 20, 1570–1582. [Google Scholar] [CrossRef]
- Andrade-Lima, A.; Silva Junior, N.; Chehuen, M.; Miyasato, R.; Souza, R.W.A.; Leicht, A.S.; Brum, P.C.; de Oliveira, E.M.; Wolosker, N.; Forjaz, C.L.M. Walking Training Improves Systemic and Local Pathophysiological Processes in Intermittent Claudication. Eur. J. Vasc. Endovasc. Surg. 2021, 61, 954–963. [Google Scholar] [CrossRef]
- Toti, L.; Bartalucci, A.; Ferrucci, M.; Fulceri, F.; Lazzeri, G.; Lenzi, P.; Soldani, P.; Gobbi, P.; La Torre, A.; Gesi, M. High-intensity exercise training induces morphological and biochemical changes in skeletal muscles. Biol. Sport 2013, 30, 301–309. [Google Scholar] [CrossRef]
- Bartalucci, A.; Ferrucci, M.; Fulceri, F.; Lazzeri, G.; Lenzi, P.; Toti, L.; Serpiello, F.R.; La Torre, A.; Gesi, M. High-intensity exercise training produces morphological and biochemical changes in adrenal gland of mice. Histol. Histopatholy 2012, 27, 753–769. [Google Scholar]
- Zhou, X.; Cao, H.; Wang, M.; Zou, J.; Wu, W. Moderate-intensity treadmill running relieves motion-induced post-traumatic osteoarthritis mice by up-regulating the expression of lncRNA H19. Biomed. Eng. Online 2021, 20, 111. [Google Scholar] [CrossRef] [PubMed]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lavier, J.; Bouzourène, K.; Millet, G.P.; Mazzolai, L.; Pellegrin, M. Physical Performance and Skeletal Muscle Transcriptional Adaptations Are Not Impacted by Exercise Training Frequency in Mice with Lower Extremity Peripheral Artery Disease. Metabolites 2023, 13, 562. https://doi.org/10.3390/metabo13040562
Lavier J, Bouzourène K, Millet GP, Mazzolai L, Pellegrin M. Physical Performance and Skeletal Muscle Transcriptional Adaptations Are Not Impacted by Exercise Training Frequency in Mice with Lower Extremity Peripheral Artery Disease. Metabolites. 2023; 13(4):562. https://doi.org/10.3390/metabo13040562
Chicago/Turabian StyleLavier, Jessica, Karima Bouzourène, Grégoire P. Millet, Lucia Mazzolai, and Maxime Pellegrin. 2023. "Physical Performance and Skeletal Muscle Transcriptional Adaptations Are Not Impacted by Exercise Training Frequency in Mice with Lower Extremity Peripheral Artery Disease" Metabolites 13, no. 4: 562. https://doi.org/10.3390/metabo13040562
APA StyleLavier, J., Bouzourène, K., Millet, G. P., Mazzolai, L., & Pellegrin, M. (2023). Physical Performance and Skeletal Muscle Transcriptional Adaptations Are Not Impacted by Exercise Training Frequency in Mice with Lower Extremity Peripheral Artery Disease. Metabolites, 13(4), 562. https://doi.org/10.3390/metabo13040562