
Is the Novel Slot Blot a Useful Method for Quantification of Intracellular Advanced Glycation End-Products?


Abstract
:1. Introduction
2. Traditional Slot Blot for Detection/Quantification of RNA, DNA, and Proteins
3. Methodology and Characteristics of the Novel Slot Blot
3.1. Equipment
3.2. Preparation and Application of Lysis Buffer
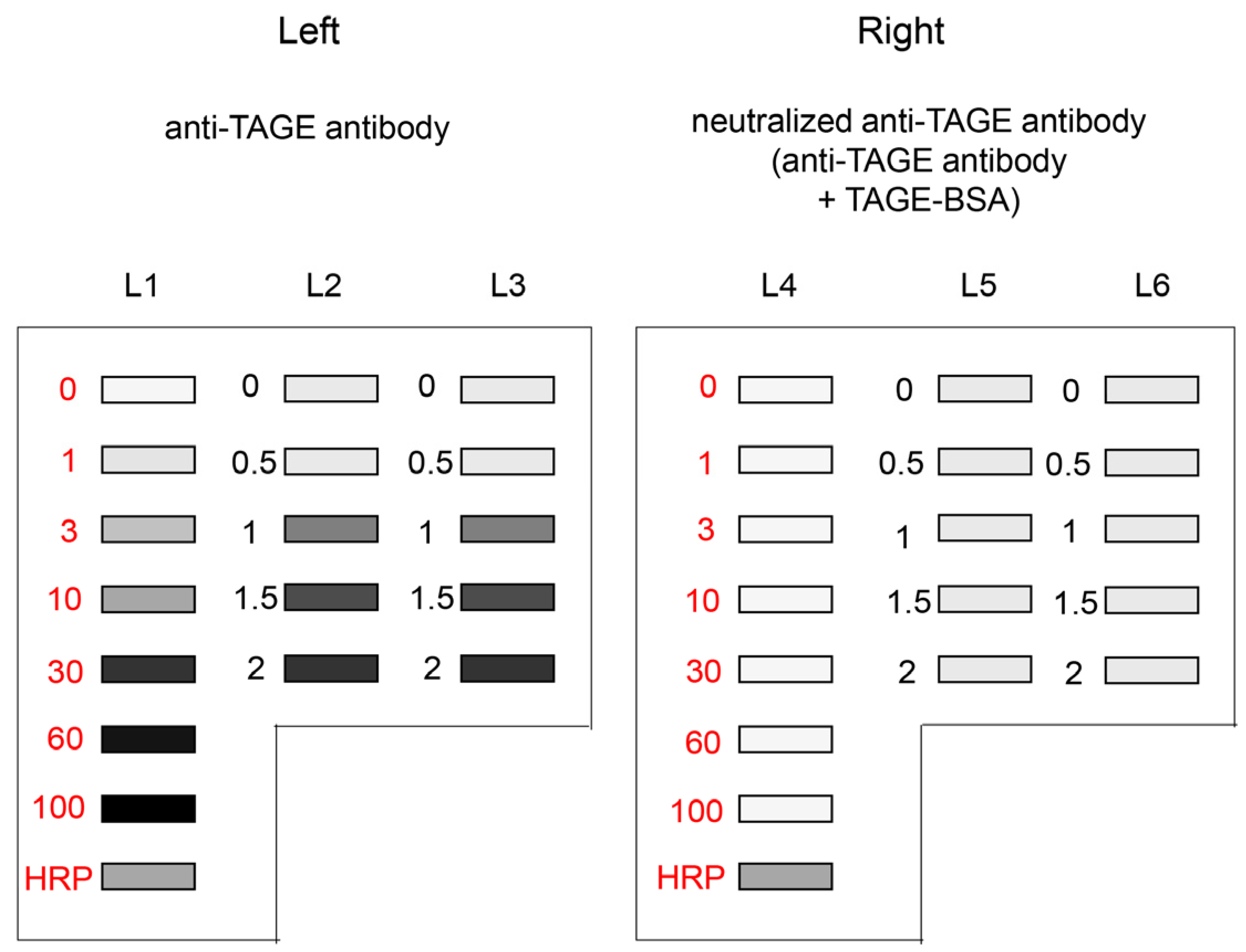
3.3. Application of Novel Slot Blot Assay for Standard AGE-Modified Protein Aliquots, Horseradish Peroxidase (HRP) Marker Solution, and Sample Solution
3.4. Chemiluminescene Imaging of Bands on Membranes and Evaluation of AGE Content
4. Comparison between the Novel Slot Blot Method and Other Methods for AGEs Quantification
4.1. Comparison with the Traditional Slot Blot or Dot Blot
4.2. Comparison with Western Blot
4.3. Comparison with Immunostaining
4.4. Comparison with ELISA
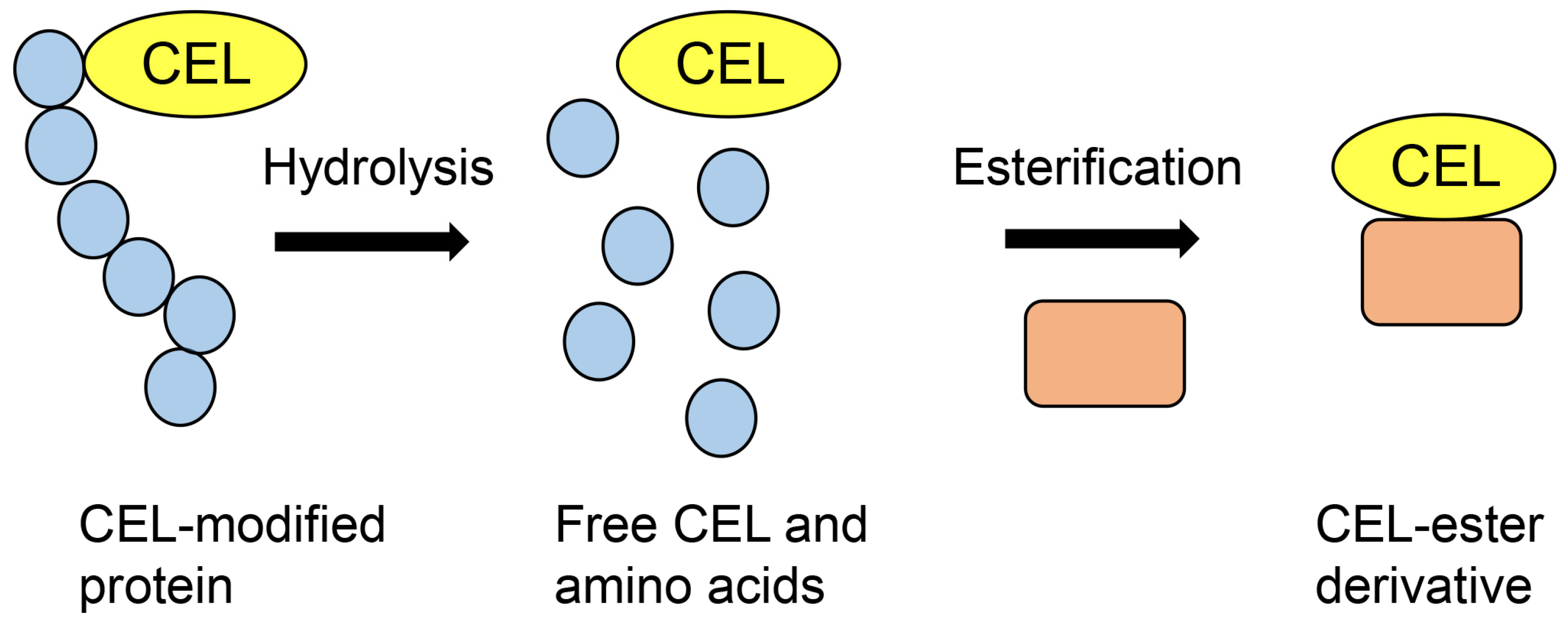
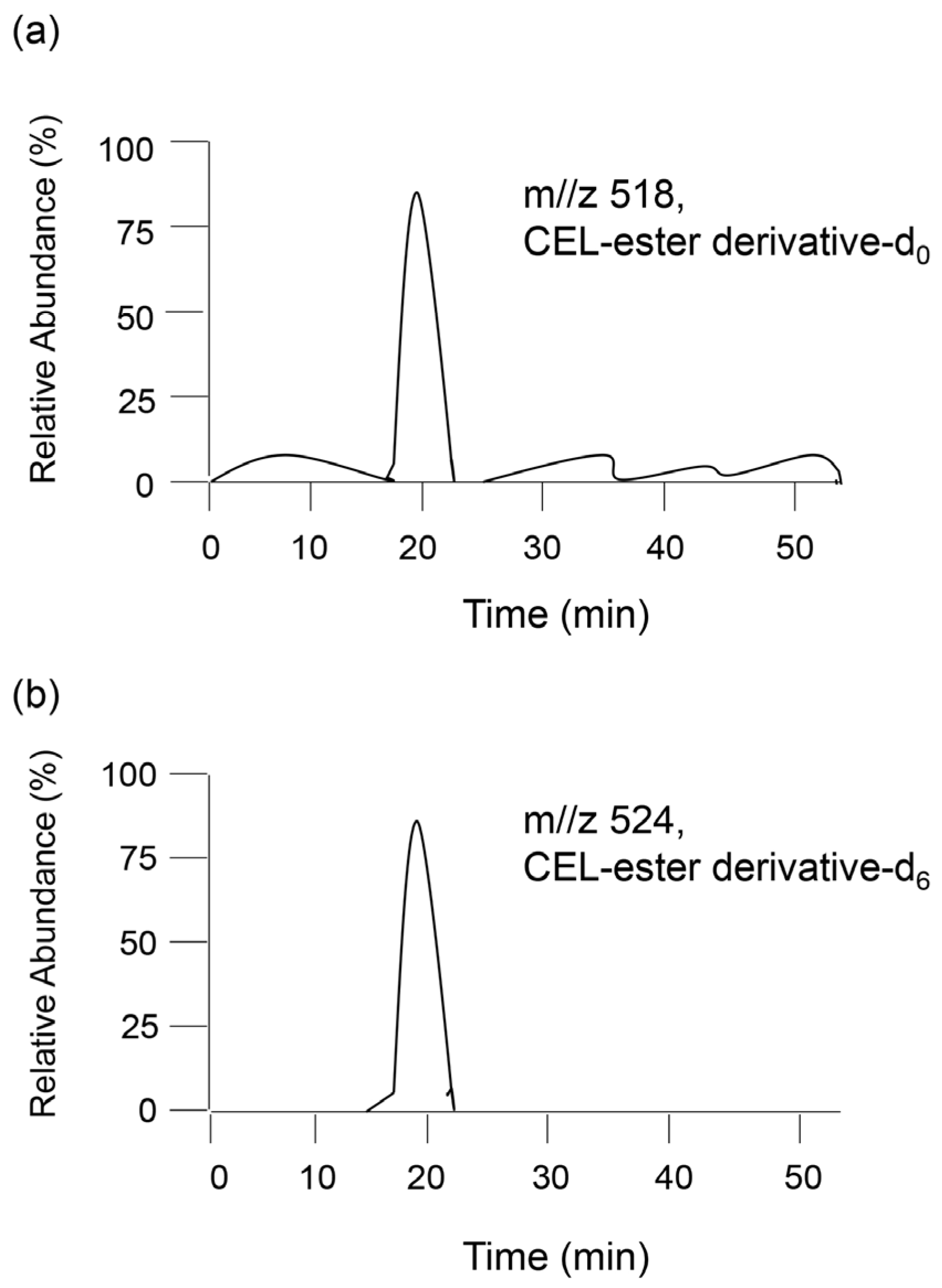
4.5. Comparison with GC–MS
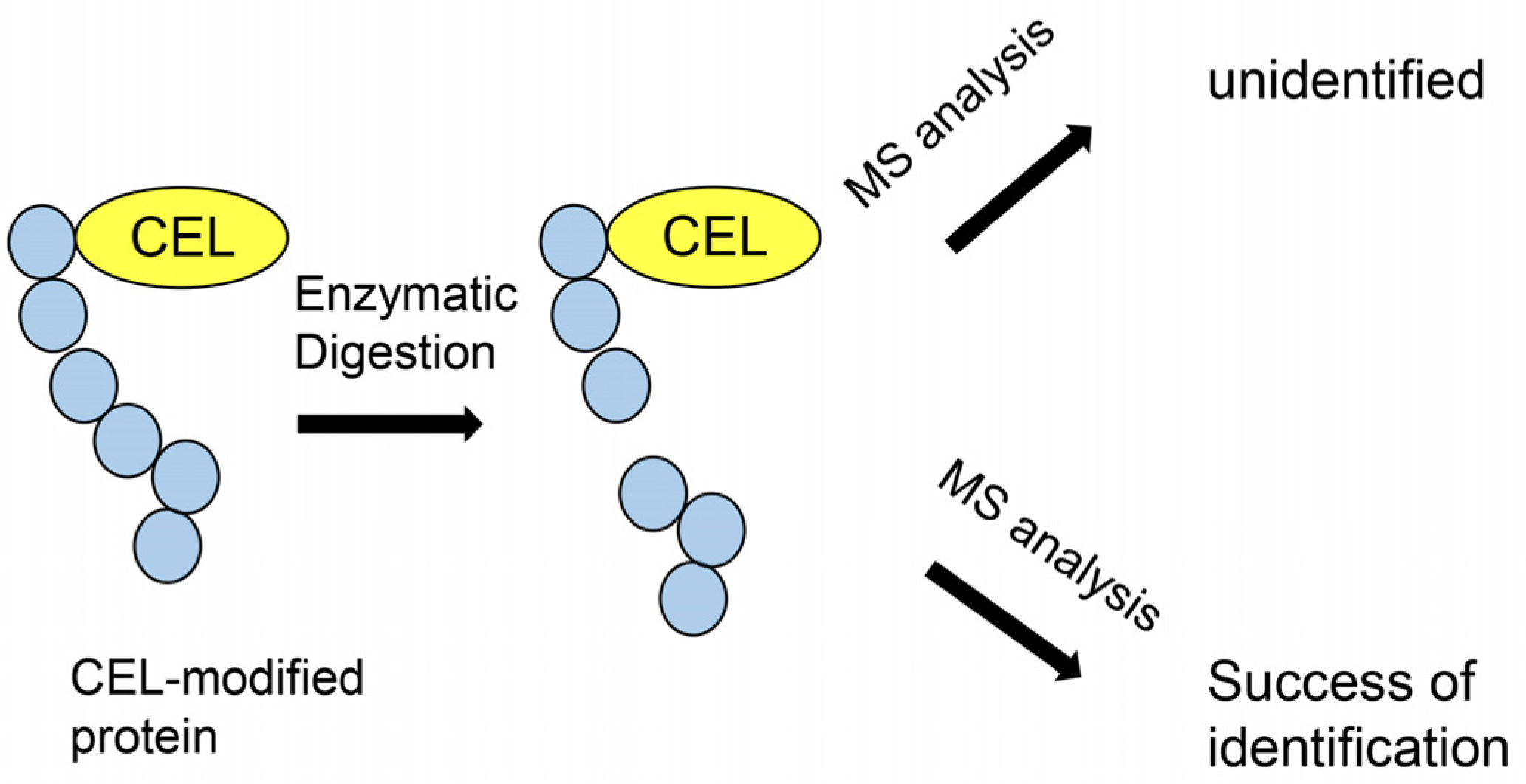
4.6. Comparison with MALDI–MS and LC–ESI–MS
5. Conclusions
Funding
Conflicts of Interest
Abbreviations
| AGEs | Advanced glycation end products |
| CHAPS | 3-[(3-cholamidopropyl)-dimethyl-ammonio]-1-propane sulfonate |
| ESI | Electrospray ionization |
| ESI-MS | ESI-mass spectrometry |
| GC | Gas chromatography |
| LC | Liquid chromatography |
| MS | Mass spectrometry |
| MALDI | Matrix-associated laser desorption/ionization |
| PVDF | Polyvinylidene difluoride |
| Tris | Tris-(hydroxymethyl)-aminomethane |
References
- Takauchi, M.; Sakasai-Sakai, A.; Takata, T.; Takino, J.; Koriyama, Y. Effects of Toxic AGEs (TAGE) on Human Health. Cells 2022, 11, 2178. [Google Scholar] [CrossRef]
- Takeuchi, M. Serum Levels of Toxic AGEs (TAGE) May Be a Promising Novel Biomarker for the Onset/Progression of Lifestyle-Related Diseases. Diagnostics 2016, 6, 23. [Google Scholar] [CrossRef] [PubMed]
- Mastrocola, R.; Collino, M.; Nigro, D.; Chiazza, F.; D’Antona, G.; Aragno, M.; Minetto, M. Accumulation of Advanced Glycation End-Products and Activation of the SCAP/SREBP Lipogenetic Pathway Occur in Diet-Induced Obese Mouse Skeletal Muscle. PLoS ONE 2015, 10, e0119587. [Google Scholar] [CrossRef]
- Chang, G.; Yeh, Y.; Chen, W.; Ko, Y.; Pang, J.S.; Lee, H. Inhibition of Advanved Glycation End-Products Formation Attenuates Cardiac Electrical and Mechanical Remodeling and Vulnerability to Tachyarrthmias in Diabetic Rats. J. Pharmacol. Exp. Ther. 2019, 368, 66–78. [Google Scholar] [CrossRef] [PubMed]
- Jung, W.K.; Park, S.; Kim, H.R.; Ryu, H.Y.; Kim, Y.H.; Kim, J. Advanced Glycation End Products Increase Salivary Gland Hypofunction in D-Galactose-Induced Aging Rats and Its Prevention by Physical Exercise. Curr. Issues Mol. Biol. 2021, 43, 2059–2067. [Google Scholar] [CrossRef]
- Kehm, R.; Rückrimen, J.; Weber, D.; Deubel, S.; Grnue, T.; Höhn, A. Eddogenous advanced glycation end products in pancreatic islets after short-term carbohydrate intervention in obese, diabetes-prone mice. Nutr. Diabetes 2019, 9, 9. [Google Scholar] [CrossRef] [PubMed]
- LeWinter, M.M.; Taatjes, D.; Ashikaga, T.; Palmer, B.; Bishop, N.; VanBuren, P.; Bell, S.; Donaldson, C.; Meyer, M.; Margulies, K.B.; et al. Abundance, localization, and functional correlates of the advanced glycation end-product carboxymethyl lysine in human myocardium. Physiol. Rep. 2017, 5, e13462. [Google Scholar] [CrossRef]
- Ruiz-Meana, M.; Minguet, M.; Bou-Teen, D.; Miro-Casas, E.; Castans, C.; Castellano, J.; Bonzon-Kulichenko, E.; Igual, A.; Rodriguez-Lecoq, R.; Vázquez, J.; et al. Ryanodine Receptor Glycation Favors Mitochondrial Damage in the Senescent Heart. Circulation 2019, 139, 949–964. [Google Scholar] [CrossRef] [PubMed]
- Suh, K.; Choi, E.; Jung, W.; Kim, Y.J.; Hong, S.M.; Park, S.Y.; Rhee, S.Y.; Chon, S. Deoxyactein protects pancreatic β-cells against methylglyoxal-induced oxidative cell damage by the upregulation of mitochondrial biogenesis. Int. J. Mol. Med. 2017, 40, 539. [Google Scholar] [CrossRef]
- Baskal, S.; Tsikas, D. Free L-Lysine and Its Methyl Ester React with Glyoxal and Methylglyoxyal in Phosphate Buffer (100 mM, pH 7.4) to Form Nε-Carboxymethel-Lysine, Nε-Carboxytheyl-Lysine and Nε-Hydroxymethyl-Lysine. Int. J. Mol. Sci. 2022, 23, 3446. [Google Scholar] [CrossRef]
- Iles, R.K.; Iles, J.K.; Lacey, J.; Gardiner, A.; Zmuidinaite, R. Direct Detection of Glycated Human Serum Albumin and Hyperglycosylated IgG3 in Serum, by MALDI-ToF-Mass Spectrometry, as predictor of COVID-19 severity. Diagnosis 2022, 12, 2521. [Google Scholar] [CrossRef] [PubMed]
- Ohno, R.; Ichimaru, K.; Tanaka, S.; Sugawa, H.; Katsuta, N.; Sakake, S.; Tominaga, Y.; Ban, I.; Shirakawa, J.; Yamaguchi, Y.; et al. Glucoselysine is derived from fructose and accumulates in the eye lens of diabetic rats. J. Biol. Chem. 2019, 294, 17326–17338. [Google Scholar] [CrossRef]
- Browicka-Szydełko, A.; Krzystek-Korpacka, M.; Kuzan, A.; Gostomaska-Pampuch, K.; Gacka, M.; Jakobsche-Policht, U.; Adamiec, R.; Gamian, A. Non-standard AGE4 epitopes that predict polyneuropathy independently of obesity can be detected by slot dot-blot immunoassay. Adv. Clin. Exp. Med. 2020, 29, 91–100. [Google Scholar] [CrossRef]
- Koriyama, Y.; Furukawa, A.; Muramatsu, M.; Takino, J.; Takeuchi, M. Glyceraldehyde caused Alzheimer’s disease-like alterations in diagnostic marker levels in SH-SY5Y human neuroblastoma cells. Sci. Rep. 2015, 5, 13313. [Google Scholar] [CrossRef] [PubMed]
- Takata, T.; Ueda, T.; Sakasa-Sakai, A.; Takeuchi, M. Generation of glyceraldehyde-derived advanced glycation end-products in pancreatic cancer cells and the potential of tumor promotion. World J. Gastroenterol. 2017, 23, 4910–4919. [Google Scholar] [CrossRef]
- Papadaki, M.; Holewinski, R.J.; Previs, S.B.; Martin, T.G.; Stachowski, M.J.; Li, A.; Blair, C.A.; Morave, C.S.; Van Eyk, J.E.; Campbell, K.S.; et al. Diabetes with heart failure increases methylglyoxal modifications in the sarcomere, which inhibit function. JCI Insight 2018, 3, e121264. [Google Scholar] [CrossRef] [PubMed]
- Gil-Agusti, M.T.; Campostrini, N.; Zolla, L.; Ciambella, C.; Invernizzi, C.; Righetti, G. Two-dimensional mapping as a tool for classification of green coffee bean species. Proteomics 2005, 5, 710–718. [Google Scholar] [CrossRef]
- Cui, Y.; Tian, M.; Zong, M.; Teng, M.; Chen, Y.; Lu, J.; Jiang, J.; Liu, X.; Han, J. Proteomics Analysis of Pancreatic Ductal Adenocarcinoma Compared with Normal Adjacent Pancreatic Tissue and Pancreatic Benign Cystadenoma. Pancreatology 2009, 9, 89–98. [Google Scholar] [CrossRef]
- Sakolvaree, Y.; Maneewatch, S.; Jiemsup, S.; Klaysing, B.; Tongtawe, P.; Srimanote, P.; Saengjaruk, P.; Banyen, S.; Tapchaisri, P.; Chonsa-nguan, M.; et al. Proteome and Immunome of Pathogenic Leptospira spp. Revealed by 2DE and 2DE-Immunoblotting with Immune Serum. Asian Pac. J. Allergy Immunol. 2007, 25, 53–73. [Google Scholar] [PubMed]
- Sakasai-Sakai, A.; Takata, T.; Takino, J.; Takeuchi, M. Impact of intracellular glyceraldehyde-derived advanced glycation end-products on human hepatocyte cell death. Sci. Rep. 2017, 7, 14282. [Google Scholar] [CrossRef] [PubMed]
- Sakasai-Sakai, A.; Takata, T.; Takeuchi, M. Intracellular Toxic Advanced Glycation End-Products Promote the Production of Reactive Oxygens Species in HepG2 Cells. Int. J. Mol. Sci. 2020, 21, 4861. [Google Scholar] [CrossRef]
- Kikuchi, C.; Sakasa-Sakai, A.; Okimura, R.; Tanaka, H.; Takata, T.; Takeuchi, M.; Matsunaga, T. Accumulation of Toxic Advanced Glycation End-Products Induces Cytotoxicity and Inflammation in Hepatocyte-Like Cells Differentiated from Human Induced Pluripotent Stem Cells. Biol. Pharm. Bull. 2021, 44, 1399–1402. [Google Scholar] [CrossRef] [PubMed]
- Takata, T.; Sakasa-Sakai, A.; Takino, J.; Takeuchi, M. Evidence for Toxic Advanced Glycation End-Poducts Generated in the Normal Rat Liver. Nutrients 2019, 11, 1612. [Google Scholar] [CrossRef] [PubMed]
- Takata, T.; Sakasa-Sakai, A.; Ueda, T.; Takeuchi, M. Intracellular toxic advanced glycation end-products in cardiomyocytes may cause cardiovascular disease. Sci. Rep. 2019, 9, 2121. [Google Scholar] [CrossRef]
- Takata, T.; Sakasa-Sakai, A.; Takeuchi, M. Intracellular Toxic Advanced Glycation End-Products May Induce Cell Death and Suppress Cardiac Fibroblasts. Metabolites 2022, 12, 615. [Google Scholar] [CrossRef]
- Takata, T.; Sakasai-Sakai, A.; Takeuchi, M. Intracellular Toxic Advanced Glycation End-Products in 1.4E7 Cell Line Induce Death with Reduction of Microtubule-Associated Protein 1 Light Chain 3 and p62. Nutrients 2022, 14, 332. [Google Scholar] [CrossRef]
- Takata, T.; Sakasai-Sakai, A.; Takeuchi, M. Impact of intracellular toxic advanced glycation end-products (TAGE) on murine myoblast cell death. Diabetol. Metab. Syndr. 2020, 12, 54. [Google Scholar] [CrossRef]
- Sakasai-Sakai, A.; Takata, T.; Takeuchi, M. The Association between Accumulation of Toxic Advanced Glycation End-Products and Cytotoxic Effect in MC3T3-E1 Cells. Nutrients 2022, 14, 990. [Google Scholar] [CrossRef]
- Inoue, S.; Takata, T.; Nakazawa, Y.; Nakamura, Y.; Guo, X.; Yamada, S.; Ishigaki, Y.; Takeuchi, M.; Miyazawa, K. Potential of an Interorgan Network Mediated by Toxic Advanced Glycation End-Products in a Rat Model. Nutrients 2020, 13, 80. [Google Scholar] [CrossRef] [PubMed]
- Sakasai-Sakai, A.; Takata, T.; Suzuki, H.; Maruyama, I.; Motomiya, Y.; Takeuchi, M. Immunological evidence for in vivo production of novel advanced glycation end-products from 1,5-anhydro-D-fructose, a glycogen metabolite. Sci. Rep. 2019, 9, 10194. [Google Scholar] [CrossRef]
- Williason, J.D.; Quantrano, R.S.; Cuming, A.C. Em polypeptide and its messenger RNA levels are modulated by ascisic acid during embryogenesis in wheat. Eur. J. Biochem. 1985, 152, 501–507. [Google Scholar] [CrossRef]
- Gubler, U.; Chua, A.O.; Young, D.; Fan, Z.-W.; Eng, J. Cholecystokinin mRNA in Porcine Cerebellum. J. Biol. Chem. 1987, 262, 15242–15425. [Google Scholar] [CrossRef] [PubMed]
- Nimmer, D.; Bergtrom, G.; Hirano, H.; Amran, D.L. Regulation of Plasma Fibrinectin Biosynthesis by Glucocorticoids in Chick Hepatocyte Cultures. J. Biol. Chem. 1987, 262, 10369–10375. [Google Scholar] [CrossRef] [PubMed]
- Nehls, P.; Adamkiwicz, J.; Rajewsky, M.F. Immuno-Slot-Blot: A Highly Sensitive Immunoassay for the Quantitation of Caracinogen-modified Nucleosides in DNA. J. Cancer Res. Clin. Oncol. 1984, 108, 23–29. [Google Scholar] [CrossRef] [PubMed]
- Vaughn, C.W.; Selby, C.P.; Yang, Y.; Hsu, D.S.; Sancar, A. Genome-wide single-nucleotide resolution of oxaliplatin-DNA adduct repair in drug-sensitive and -resistant colorectal cancer cell line. J. Biol. Chem. 2020, 295, 7584–7594. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.T.; Jagannath, S.; Francois, C.; Vanderstichele, H.; Stoops, E.; Lashuel, H.A. How specific are the conformation-specific α-synuclein antibodies? Characterization and validation of 16 α-synuclein conformation-specific antibodies using well-characterized preparations of α-synuclein monomers, fibris and oligomers with distinct structures and morphology. Neutobiol. Dis. 2020, 146, 10586. [Google Scholar]
- Ghiani, A.; Ania, R.; Asero, R.; Bellotto, E.; Citterio, S. Ragweed pollen collected along high-traffic roads shows a higher allergenicity than pollen sampled in vegetated areas. Allergy 2012, 67, 887–894. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Zheng, H.; Deng, B.; Mahajan, B.; Grabias, B.; Kozakai, Y.; Morin, M.J.; Locke, E.; Birkett, A.; Miura, K.; et al. A Slot Blot Immunoassay for Quantitative Detection of Plasmodium falciparum Circumsporozoite Protein in Mosquito Midgut Oocyst. PLoS ONE 2014, 9, e115807. [Google Scholar] [CrossRef] [PubMed]
- Grabias, B.; Verma, N.; Zheng, H.; Tripathi, A.K.; Mlambo, G.; Morin, M.J.; Locke, E.; Kumar, S. A no film slot blot for the detection of developing P. falciparum oocysts in mosquitoes. PLoS ONE 2017, 12, e0174229. [Google Scholar]
- Nimmo, J.T.; Verma, A.; Dodart, J.-C.; Wang, C.Y.; Savistchenko, J.; Melki, R.; Carare, R.O.; Nicoll, J.A.R. Novel antibodies detect additional α-synuclein pahaology in synucleinopathies: Potential development for immunotherapy. Alzheimer’s Res. Ther. 2020, 12, 159. [Google Scholar] [CrossRef]
- McCarthy, J.; Hopwood, F.; Oxley, D.; Laver, M.; Castagna, A.; Righetti, P.G.; Williams, K.; Herbert, B. Carbamylation of Protein in 2-D Electrophoresis-Myth or Reality? J. Proteome Res. 2003, 2, 239–242. [Google Scholar] [CrossRef] [PubMed]
- Herbert, B. Advances in protein solubilization for two-dimensional electrophoresis. Electrophoresis 1999, 20, 660–663. [Google Scholar] [CrossRef]
- Twine, S.M.; Mykytczuk, N.C.S.; Petit, M.; Tremblay, T.; Conlan, J.W.; Kelly, J.F. Francisella tularensis Proteome: Low Levels of ASB-14 Facilitate the Visualization of Membrane Proteins in Total Protein Extracts. J. Proteome Res. 2005, 4, 1848–1854. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Zhou, J.; Zhang, X.; Zheng, X.; Jiang, X.; Shi, L.; Yin, W.; Wang, J. Optimized samples preparation for two-dimensional gel electrophoresis of soluble proteins from chicken bursa of Fabricus. Proteome Sci. 2009, 7, 38. [Google Scholar] [CrossRef] [PubMed]
- Campos, A.; Puetro, M.; Prieto, A.; Cameán, A.; Almeida, A.M.; Coelho, A.; Vasconcelos, V.J. Protein extraction and two-dimensional gel electrophoresis of proteins in the marine mussel Mytilus galloprovincialis: An important tool for protein expression studies, food quality and safety assessment. Sci. Food Agric. 2013, 93, 1779–1787. [Google Scholar] [CrossRef]
- Pedroso, A.P.; Watanabe, R.L.H.; Albuquerque, K.T.; Telles, M.M.; Andrade, M.C.C.; Perez, J.D.; Sakata, M.M.; Lima, M.L.; Estadella, D.; Nascimento, C.M.O.; et al. Proteomic profiling of the rat hypothalamus. Proteome Sci. 2012, 10, 26. [Google Scholar] [CrossRef]
- Farinazzo, A.; Fasoli, E.; Kravchuk, A.V.; Candiano, G.; Aldini, G.; Regazzoni, L.; Righetti, P.G. En bloc elution of proteomes from combinatorial peptide ligand libraries. J. Proteom. 2009, 72, 725–730. [Google Scholar] [CrossRef] [PubMed]
- Tit-Oon, P.; Chokchaichamnankit, D.; Khongmanee, A.; Sawangreetrakul, P.; Svasti, J.; Srisomsap, C. Comparative secretome analysis of cholanagiocaricinoma cell line in three-dimensional culture. Int. J. Oncol. 2014, 45, 2108–2116. [Google Scholar] [CrossRef]
- Kawanami, T.; Kawanami-Iwao, H.; Takata, T.; Ishigaki, Y.; Tomosugi, N.; Takegami, T.; Yanagisawa, H.; Fujimoto, S.; Sakai, T.; Fujita, Y.; et al. Comprehensive analysis of protein-expression changes specific to immunoglobulin G4-related disease. Clin. Chim. Acta 2021, 523, 45–57. [Google Scholar] [CrossRef]
- Takata, T.; Ishigaki, Y.; Shimasaki, T.; Tsuchida, H.; Motoo, Y.; Hayashi, A.; Tomosugi, N. Characterization of proteins secreted by pancreatic cancer cells with anticancer drug treatment in vitro. Oncol. Rep. 2014, 28, 1968–1976. [Google Scholar] [CrossRef]
- Takeuchi, M.; Makita, Z.; Bucala, R.; Suzuki, T.; Koike, T.; Kameda, Y. Immunological Evidence that Non-carboxymethyllisine Advanced Glycation End-products Are Produced from Short Chain Sugars and Dicarbonyl Compounds in vivo. Mol. Med. 2000, 6, 114–125. [Google Scholar] [CrossRef] [PubMed]
- Barandalla, M.; Haucke, E.; Fischer, B.; Santos, A.N.; Colleoni, S.; Galli, C.; Santos, A.N.; Lazzari, G. Comparative Analysis of AGE and RAGE Levels in Human Somatic and Embryonic Stem Cell under H2O2-Induced Noncytotoxic Oxidative Stress Conditions. Oxidative Med. Cell. Longev. 2017, 2017, 4240136. [Google Scholar] [CrossRef] [PubMed]
- Nasu, R.; Furukawa, A.; Suzuki, K.; Takeuchi, M.; Koriyama, Y. The Effect of Glyceraldehyde-Derived Advanced Glycation End Products on β-Tubulin-Inhibited Neurite Outgrowth in SH-SY5Y Human Neuroblastoma Cells. Nutrients 2020, 12, 2958. [Google Scholar] [CrossRef] [PubMed]
- Takino, J.; Kobayashi, Y.; Takeuchi, M. The formation of intracellular glyceraldehyde-derived advanced glycation end-products and cytotoxicity. J. Gastroenterol. 2010, 45, 646–653. [Google Scholar] [CrossRef] [PubMed]
- Cui, Y.; Liu, P.; Yu, S.; He, J.; Afedo, S.Y.; Zou, S.; Zahang, Q.; Liu, J.; Song, L.; Xu, Y.; et al. Expression Analysis of Molecular Chaperones Hsp70 and Hsp90 on Development and Metabolism of Different Organs and Testis in Cattle (Cattle-yak and Yak). Metabolites 2022, 12, 1114. [Google Scholar] [CrossRef]
- Chung, Y.-H.; Hung, T.-H.; Yu, C.-F.; Tsai, C.-K.; Weng, C.-C.; Jhang, F.; Chen, F.-H.; Lin, G. Glycolytic Plasticity of Metastatic Lung Cnacer Captured by Noninvasive 18F-FDG PET/CT and Serum 1H-NMR Analysis: An Orthotopic Murine Model Study. Metabolites 2023, 13, 110. [Google Scholar] [CrossRef]
- Jia, J.; Duan, H.; Liu, B.; Ma, Y.; Ma, Y.; Cai, X. Alfalfa Xeno-miR168b Target CPT1A to Regulate Milk Fat Synthesis in Bovine Mammary Epithelial Cells. Metabolites 2023, 13, 76. [Google Scholar] [CrossRef]
- Wu, F.; Li, Y.; Liu, W.; Xiao, R.; Yao, B.; Gao, M.; Xu., D.; Wang, J. Comparative Investigation of Raw and Processed Radix Polygoni Multiflori on the Treatment of Vascular Dementia by Liquid Chromatograph-Mass Spectrometry Based Metabolomic Approach. Metabolites 2022, 12, 1297. [Google Scholar] [CrossRef]
- Aburto-Hernámdez, C.; Barrera, D.; Ortiz-Hernández, R.; Espinoza-Simón, E.; parra-Gámez, L.; González, J.; Escobar, M.L.; Vánquez-Nin, G.H.; Echeverrínez, O.; Torres-Ramírez, N. Impaired Carbohydrate Metabolism and Excess of Lipid Accumulation of Offspring of Hyperandrogenic Mice. Metabolites 2022, 12, 1182. [Google Scholar] [CrossRef]
- Kim, M.-J.; Kwak, H.-W.; Kin, S.-H.; Lee, H.-J.; Seo, J.-W.; Kim, J.-T.; Jang, S.-H.; Kim, M.-J.; Kim, Y.-M. Anti-Obesity Effect of Hot Water Extract of Bareley Sprout through the Inhibition of Adipocyte Differentiation and Growth. Metabolites 2021, 11, 610. [Google Scholar] [CrossRef]
- Jahrom, A.S.; Shojaei, M.; Ghobadifar, A. Insulin Resistance and Serum Levels of Interleukin-17 and Interleukin-18 in Normal Pregnancy. Immune New 2014, 14, 149–155. [Google Scholar] [CrossRef]
- Mi, L.; Zhang, Y.; Xu, Y.; Zheng, X.; Zhang, X.; Xue, M.; Jin, X. HMGB-1/RAGE pro-inflammatory axis promotes vascular endothelial cell apoptosis in limb ischemia/reperfusion injury. Biomed. Pharm. 2019, 116, 109005. [Google Scholar] [CrossRef] [PubMed]
- Indky, D.; Bronowicka-Szydełko, A.; Gamian, A.; Kuzan, A. Advanced glycation end prodcuts and their receptors in serum of patients with type 2 diabetes. Sci. Rep. 2021, 11, 13264. [Google Scholar] [CrossRef]
- Damasiewicz-Bodzek, A.; Łabuz-Rozak, B.; Kumaszka, B.; Tadeusiak, B.; Typień-Colder, K. The Assessment of Serum Concentrations of AGEs and Their Soluble Receptor (sRAGE) in Multiple Sclerosis Patients. Brain Sci. 2021, 11, 1021. [Google Scholar] [CrossRef]
- Matsui, T.; Joo, H.D.; Lee, J.M.; Ju, S.M.; Tao, W.H.; Higashimoto, Y.; Fukami, K.; Yamagishi, S. Development of a monoclonal antibody-based ELISA system for glyceraldehyde-derived advanced glycation end products. Immunol. Lett. 2015, 167, 141–146. [Google Scholar] [CrossRef]
- Hori, E.; Kikuchi, C.; Nagami, C.; Kajikuri, J.; Itoh, T.; Takeuchi, M.; Matsunaga, T. Role of Glyceraldehyde-Derived AGEs and Mitochondria in Superoxide Production in Femoral Artery of OLETF Rat and Effects of Pravastatin. Biol. Pharm. Bull. 2017, 40, 1903–1908. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, K.; Sakasai-Sakai, A.; Motomiya, Y.; Yoneda, T.; Takeuchi, M. Serum levels of 1.5-anhydroglucitol and 1.5-anhydrofructose-derived advanced glycation end products in patients undergoing hemodialysis. Diabetol. Metab. Syndr. 2021, 13, 85. [Google Scholar] [CrossRef]
- Pinto, R.S.; Ferreira, G.S.; Silvestre, C.R.; de Fátima M Santana, M.; Nunes, V.S.; Ledesma, L.; Pinto, P.R.; de Assis, S.I.; Machado, U.F.; da Silva, E.S.; et al. Plasma advanced glycation end products and soluble receptor for advanced glycation end products as indicators of sterol content in human carotid atherosclerotic plaques. Diab. Vasc. Dis. Res. 2022, 19, 14791641221085269. [Google Scholar] [CrossRef] [PubMed]
- Kashiwabara, S.; Hosoe, H.; Ohno, R.; Nagai, R.; Shiraki, M. Department and Evaluation of Novel ELISA for Determination of Urinary Pentosidine. J. Nutr. Sci. 2019, 65, 526–533. [Google Scholar]
- Ouyang, F.; Wang, Y.; Xu, L.; Li, D.; Li, F.; Sun-Waterhouse, D. Attenuation of Palmitic-Induced Interstinal Epithelial Barrier Dysfunction by 6-Shoganol in Caco-2 Cells: The Role of MiR-216a-5p/TLR4/NF-κB Axis. Metabolites 2022, 12, 1028. [Google Scholar] [CrossRef]
- Kinoshita, S.; Mera, K.; Ichikawa, H.; Shimasaki, S.; Nagai, M.; Taga, Y.; Iijima, K.; Hattori, S.; Fujiwara, Y.; Shirakawa, J.; et al. Nω-(Carboxymethyl)arginine Is One of the Dominant Advanced Glycation End Products in Glycated Collagens and Mouse Tissues. Oxidative Med. Cell. Longev. 2019, 2019, 9073451. [Google Scholar] [CrossRef]
- Tominaga, Y.; Sugawa, H.; Hirabayashi, K.; Ikeda, T.; Hoshi, Y.; Nagai, R. Drosera tokainensis extract containing multiple pehonolic compounds inhibits the formation of advanced glycation end-products. Arch. Biochem. Biophys. 2020, 693, 108586. [Google Scholar] [CrossRef] [PubMed]
- Ban, H.; Sugawa, H.; Nagai, R. Protein Modification with Ribose Generates Nδ-(5-hydro-5methyl-1,4-imidazolne-2-yl)-ornithine. Int. J. Mol. Sci. 2022, 23, 1224. [Google Scholar] [CrossRef] [PubMed]
- Baskal, S.; Kaiser, A.; Mels, C.; Kruger, R.; Tsikas, D. Specific and sensitive GC-MS analysis of hypusine, Nε-(4-amino-2-hydoroxybutyle)lysine, a biomarker of hypusinated eukaryotic initiation factor elF5A, and its application to the bi-ethnic ASOS study. Amino Acids 2022, 54, 1083–1099. [Google Scholar] [CrossRef]
- Cai, J.; Hurst, H.E. Identification and Quantitation of N-(Carboxymethyl)valine Adduct in Hemoglobin by Gas Chromatography/Mass Spectrometry. J. Mass Spectrom. 1999, 34, 537. [Google Scholar] [CrossRef]
- Requena, J.R.; Ahmed, M.U.; Fountain, W.F.; Degenhardt, T.P.; Reddy, S.; Perez, C.; Lyons, T.J.; Jenkins, A.J.; Baynes, J.W.; Thorpe, S.R. Carboxymethylethanolamine, a Biomarker of Phospholipid Modification during the Maillard Reaction in Vivo. J. Biol. Chem. 1997, 272, 17473–17479. [Google Scholar] [CrossRef]
- Baskal, S.; Bollenbach, A.; Mels, C.; Kruger, R.; Tskas, D. Development, validation of a GC-MS method for the simultaneous measurement of amino acids, their PTM metabolites and AGEs in human urine, and application to the bi-ethnic ASOS study with special emphasis to lysine. Amino Acids 2022, 54, 615–641. [Google Scholar] [CrossRef]
- Baskal, S.; Büttner, P.; Werner, S.; Besler, C.; Lurz, P.; Thiele, H.; Tsikas, D. Profile of urinary amino acids and their post-translational modifications (PTM) including advanced glycation end-products (AGE) of lysine, arginine and cysteine in lean and obese ZSF1 rats. Amino Acids 2022, 54, 643–652. [Google Scholar] [CrossRef]
- Fermández-Puente, P.; Mateos, J.; Blanco, F.J.; Ruiz-Romero, C. LC-MALDI-TOF-TOF for shotgun proteomics. Methods Mol. Biol. 2014, 1156, 27–38. [Google Scholar]
- Maus, A.; Mignon, B.; Basile, F. Enhanced protein identification using graphite-modified MALDI plates for offline LC-MALDI-MS/MS bottom-up proteomics. Anal. Biochem. 2018, 545, 31–37. [Google Scholar] [CrossRef]
- Niwa, T. Mass Spectrometry for Study of Protein Glycation in Disease. Mass Spectrom. Rev. 2006, 25, 713–723. [Google Scholar] [CrossRef]
- Lapolla, A.; Fedele, D.; Seraglia, R.; Traldi, P. The Role of Mass Spectrometry in the Study of Non-Enzymatic Protein Glycation in Diabetes: An update. Mass Spectrom. Rev. 2006, 25, 775–797. [Google Scholar] [CrossRef]
- Krause, R.; Kühn, J.; Knoll, P.K.; Henle, T. N-Terminal pyrazinones: A new class of peptide-bound advanced glycation end-products. Amino Acid 2004, 27, 9–18. [Google Scholar] [CrossRef]
- Fujimoto, S.; Murakami, Y.; Miyake, H.; Hayase, F.; Watanabe, H. Identification of a novel advanced glycation end product derived from lactaldehyde. Biosci. Biotechnol. Biochem. 2019, 83, 1136–1145. [Google Scholar] [CrossRef]
- Perween, S.; Abidi, M.; Faizy, A.F.; Moinuddin. Post-translational modifications on glycated plasma fibrinogen: A physicochemical insight. Int. J. Biol. Macromol. 2019, 126, 1201–1212. [Google Scholar] [CrossRef]
- Ghosh, P.; Kishore, N. Mechanistic physicochemical insights into glycation and drug binding by serum albumin: Implications in diabetic conditions. Biochimie 2022, 193, 16–37. [Google Scholar] [CrossRef]
- Zhang, Y.; Cocklin, R.R.; Bidasee, K.R.; Wang, M. Rapid Determination of Advanced Glycation End Products of Proteins Using MALDI-TOF-MS and PERL Script Peptide Searching Algorithm. J. Biomol. Tech. 2003, 14, 224–230. [Google Scholar] [PubMed]
- Heller, M.; Mattou, H.; Menzel, C.; Yao, X. Trypsin catalyzed 16O-to-18O exchange for comparative proteomics: Tandem mass spectrometry comparison using MALDI-TOF, ESI-QTOF, and ESI-ion trap mass spectrometers. J. Am. Soc. Mass Spectrom. 2003, 14, 704–718. [Google Scholar] [CrossRef]
- Gu, S.; Liu, Z.; Pan, S.; Jiang, Z.; Lu, H.; Amit, O.; Bradbury, E.M.; Hu, C.-A.A.; Chen, X. Global investigation of p53-induced apoptosis through quantitative proteomic profiling using comparative amino acid-coded tagging. Mol. Cell. Proteom. 2004, 3, 998–1008. [Google Scholar] [CrossRef] [PubMed]
- Kuang, L.; Jing, Z.; Wang, J.; Ma, L.; Liu, X.; Yang, J. Quantitative determination of ε-N-Carboxymethyl-L-lysine in human plasma by liquid chromatography-tandem mass spectrometry. J. Pharm. Biomed. Anal. 2004, 90, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Martocola, R.; Collino, M.; Rogazzo, C.; Medana, D.; Nigro, G.; Boccuzzi, M.; Argano, M. Advanced glycation end prodcuts promote hepatoseteatosis by interfering with SCAP-SREBP pathyway in fructose-dringking mice. Am. J. Physiol. Gastrointest. Liver Physiol. 2013, 305, G398–G407. [Google Scholar]
- Mastrocola, R.; Nigro, D.; Chiazza, F.; Medana, C.; Dal Bello, F.; Boccuzzi, G.; Collino, M.; Aragno, M. Fructose-derived advanced glycation end-products drive lipogenesis and skeletal muscle reprograming via SREBP-1c dysregulation in mice. Free Radic. Biol. Med. 2016, 91, 224–235. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, R.; Fujiwara, Y.; Saito, M.; Arakawa, S.; Shirakawa, J.; Yamanaka, M.; Komohara, Y.; Marumo, K.; Nagai, R. Intracellular Accumulation of Advanced Glycation End Products Induces Osteoblast Apoptosis Via Endoplasmic Reticulum Stress. J. Bone Miner. Res. 2020, 35, 1992–2003. [Google Scholar] [CrossRef]
- Kato, S.; Sugawa, H.; Tabe, K.; Ito, K.; Nakashima, H.; Nagai, R. Rapid pretretment for multi-sample analysis of advanced glycation end products and their role in nephropathy. J. Clin. Biochem. Nutr. 2022, 70, 256–261. [Google Scholar] [CrossRef] [PubMed]
- Katsuta, N.; Takahashi, H.; Nagai, M.; Sugawa, H.; Nagai, R. Changes in S-(2-succinyl)cysteine and advanced glycation end-products levels in mouse tissues associated with aging. Amino Acids 2022, 54, 653–661. [Google Scholar] [CrossRef] [PubMed]
- Senavirathna, L.; Ma, C.; Chen, R.; Pan, S. Proteomics Investigation of Glyceraldehyde-Derived Intracellular AGEs and Their Potential Influence on Pancreatic Ductal Cells. Cells 2021, 10, 1005. [Google Scholar] [CrossRef]
- Usui, T.; Hayase, F. Isolation and Identification of the 3-Hydroxy-5-hydromethyl-pyridinium Compound as a Novel Advanced Glycation End Product on Glyceraldehyde-derived Maillard Reaction. Biosci. Biotechnol. Biochem. 2003, 67, 930–932. [Google Scholar] [CrossRef]
- Usui, T.; Shimohira, K.; Watanabe, H.; Hayase, F. Detection and Determination of Glyceraldehyde-Derived Pyrimidinium-Type Advanced Glycation End Prodcut in Streptozotocin-Induced Diabetic Rats. Biomed. Biotechnol. Biochem. 2007, 71, 442–448. [Google Scholar] [CrossRef]
- Usui, T.; Shizuuchi, S.; Watanabe, H.; Hayase, F. Cytotoxicity and Oxidation Stress Induced by the Glyceraldehyde-related Maillard Reaction Products for HL-60 Cells. Biosci. Biotechnol. Biochem. 2004, 68, 333–340. [Google Scholar] [CrossRef]
- Usui, T.; Ohguchi, M.; Watanabe, H.; Hayase, F. The Formation of Argpyrimidine in Glyceraldehyde-Related Glycation. Biosci. Biotechmol. Biochem. 2008, 72, 568–571. [Google Scholar] [CrossRef]
- Usui, T.; Watanabe, H.; Hayase, F. Isolation and Identification of 5-Methyl-imidazolin-4-one Derivative as Glyceraldehyde-Derived Advanced Glycation End Product. Biosci. Biotechnol. Biochem. 2006, 70, 1496–1498. [Google Scholar] [CrossRef] [PubMed]
- Wilker, S.C.; Chellan, P.; Arnold, B.M.; Nagai, R.H. Chromatographic quantification of argpyrimidine, a methylglyoxal-dervied product in tissue proteins: Comparsion with pentosidine. Anal. Biochem. 2001, 290, 353–358. [Google Scholar] [CrossRef]
- Oya-Ito, T.; Naito, Y.; Takagi, T.; Handa, O.; Matsui, H.; Yamada, M.; Shima, K.; Yoshikawa, T. Heat-shock protein 27 (Hsp27) as a target of methylglyoxal in gastrointestinal cancer. Biochim. Biophys. Acta 2011, 1812, 769–781. [Google Scholar] [CrossRef] [PubMed]
- Sakamoto, H.; Mashima, T.; Yamamoto, K.; Tsuruo, T. Modulation of Heat-shock Protein 27 (Hsp27) Anti-apoptotic Activity by Methylglyoxal Modification. J. Biol. Chem. 2002, 277, 45770–45775. [Google Scholar] [CrossRef]
- Nokin, M.-J.; Duriex, F.; Peixoto, P.; Chiavarina, B.; Reulen, O.; Blomme, A.; Turtoi, A.; Costanza, B.; Smargisso, N.; Baiwir, D. Methylglyoxal, a glycolysis side-product, induces Hsp90 glycation and YAP-mediated tumor growth and metastasis. eLife 2016, 19, e19375. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, N.; Thornalley, P.J. Peptide Mapping of Human Serum Albumin Modified Minimally by Methylglyoxal in Vitro and in Vivo. Ann. N. Y. Acad. Sci. 2005, 1043, 260–266. [Google Scholar] [CrossRef]
- Karachallas, N.; Babaei-Jadaidi, R.; Kupich, C.; Ahmed, N.; Thornalley, P.J. High-Dose Thiamine Therapy Counters Dyslipidemia and Advanced Glycation of Plasma Protein in Streptozotoci-Induced Diabetic Rats. Ann. N. Y. Acad. Sci. 2005, 1043, 777–783. [Google Scholar] [CrossRef]
- Hayase, F.; Usui, T.; Nishiyama, K.; Sasaki, S.; Shirahashi, Y.; Tsuchiya, N.; Numata, N.; Watanabe, H. Ghemistry and Biological Effects of Melanoidins and Glyceraldehyde-Derived Pyridinium as Advanced Glycation End Products. Ann. N. Y. Acad. Sci. 2005, 1043, 104–110. [Google Scholar] [CrossRef]
- Kuzan, A. Toxicity of advanced glycation end products (Review). Biochem. Rep. 2021, 14, 46. [Google Scholar] [CrossRef]
- Tessier, F.J.; Monnier, V.M.; Sayre, L.M.; Kornfiled, J.A. Triosidines: Novel Maillard reaction products and cross-links from the reaction of triose sugars with lysine and arginine residues. Biochem. J. 2003, 369, 705–719. [Google Scholar] [CrossRef]
- Shigeta, T.; Sasamoto, K.; Yamamoto, T. Glyceraldehyde-derived advanced glycation end-products having pyrropyridinium-based crosslinks. Biochem. Biophys. Rep. 2021, 26, 100963. [Google Scholar] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Solution A | Solution B | Solution C | Solution D |
|---|---|---|---|
| 30 mM Tris | 1 protease inhibitor cocktail tablet/2 mL | 27 mM Tris | 30 mM Tris |
| 7 M urea | 6.3 M urea | 7 M urea | |
| 2 M thiourea | 1.8 M thiourea | 2 M thiourea | |
| 4% CHAPS | 3.6% CHAPS | 4% CHAPS | |
| 10% Solution B | 4% Solution B | ||
| (pH 8.5) | (pH 8.5) | (pH 8.5) |
| Solution | References |
|---|---|
| Solution C | [15,22,23,24,25,26,27,29] |
| Solution D | [20,21,28,30] |
| Cell Type | References |
|---|---|
| Tissue | [23,29] |
| Primary cells | [23,24] |
| Normal cells | [25] |
| Differentiated from iPS cells | [22] |
| Cell line | [15,20,21,26,27,28,30] |
| Organ | References |
|---|---|
| Liver | [20,21,22,23,29,30] |
| Heart | [24,25] |
| Pancreas | [15,26] |
| Skeletal muscle | [27] |
| Bone | [28] |
| Cell Type | Treatment | Cell Viability (%) | TAGE (μg/mg Protein) | Reference |
|---|---|---|---|---|
| Rat primary cardiomyocytes | 0 mM, 24 h | 100 | 0 | [24] |
| 2 mM, 24 h | 13 | 28.7 | ||
| 4 mM, 24 h | 0 | 38.5 | ||
| 4 mM, 0 h | 100 | 0 | ||
| 4 mM, 6 h | 39 | 12.0 | ||
| 4 mM, 12 h | 7 | 34.2 | ||
| Human normal cardiac fibroblasts | 0 mM, 24 h | 100 | 0.3 | [25] |
| 1 mM, 24 h | 51 | 2.8 | ||
| 1.5 mM, 24 h | 34 | 4.0 | ||
| 2 mM, 24 h | 14 | 8.1 | ||
| hiPSC-HLC | 0 mM, 24 h | 100 | 0.6 | [22] |
| 4 mM, 24 h | 20 | 5.4 | ||
| PANC-1 | 0 mM, 24 h | 100 | 0 | [15] |
| 2 mM, 24 h | 40 | 6.4 | ||
| 4 mM, 24 h | 6 | 21.2 | ||
| 1.4E7 | 0 mM, 24 h | 100 | 0.2 | [26] |
| 1.5 mM, 24 h | 61 | 3.0 | ||
| 2 mM, 24 h | 53 | 5.2 | ||
| 2.5 mM, 24 h | 33 | 6.3 | ||
| 3 mM, 24 h | 12 | 8.8 | ||
| C2C12 | 0 mM, 24 h | 100 | 0 | [27] |
| 1.5 mM, 24 h | 48 | 6.0 | ||
| 2 mM, 24 h | 5 | 15.9 | ||
| Hep3B | 0 mM, 24 h | 100 | 0 | [54] |
| 2 mM, 24 h | 46 | 0.2 | ||
| 4 mM, 0 h | 100 | 0 | ||
| 4 mM, 6 h | 65 | 0.2 | ||
| 4 mM, 12 h | 11 | 0.4 | ||
| 4 mM, 24 h | 8 | 0.9 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Takata, T. Is the Novel Slot Blot a Useful Method for Quantification of Intracellular Advanced Glycation End-Products? Metabolites 2023, 13, 564. https://doi.org/10.3390/metabo13040564
Takata T. Is the Novel Slot Blot a Useful Method for Quantification of Intracellular Advanced Glycation End-Products? Metabolites. 2023; 13(4):564. https://doi.org/10.3390/metabo13040564
Chicago/Turabian StyleTakata, Takanobu. 2023. "Is the Novel Slot Blot a Useful Method for Quantification of Intracellular Advanced Glycation End-Products?" Metabolites 13, no. 4: 564. https://doi.org/10.3390/metabo13040564
APA StyleTakata, T. (2023). Is the Novel Slot Blot a Useful Method for Quantification of Intracellular Advanced Glycation End-Products? Metabolites, 13(4), 564. https://doi.org/10.3390/metabo13040564