
Role of Elevated Ozone on Development and Metabolite Contents of Lemongrass [Cymbopogon flexuosus (Steud.) (Wats.)]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Site
2.2. Plant Material
2.3. Experimental Design
2.4. Ozone Monitoring
2.5. Plant Sampling and Assessment of Plant Parameters
2.5.1. Growth and Biomass Parameters
2.5.2. Biochemical Assays
Hydrogen Peroxide (H2O2), Superoxide Radicals (O2o−) and MDA Content
Antioxidants and Metabolites
ROS Scavenging Activity
2.5.3. Measurement of Gas Exchange and Photochemical Efficiency
2.5.4. Photosynthetic Pigments
2.5.5. Isolation of Essential Oil
2.5.6. Extraction, Analysis, and Identification of Different Metabolites of Leaf
2.6. GC–MS Conditions and Specifications
2.7. Identification of Different Metabolites of Leaf Extract and Essential Oil
2.8. Statistical Analysis
3. Result
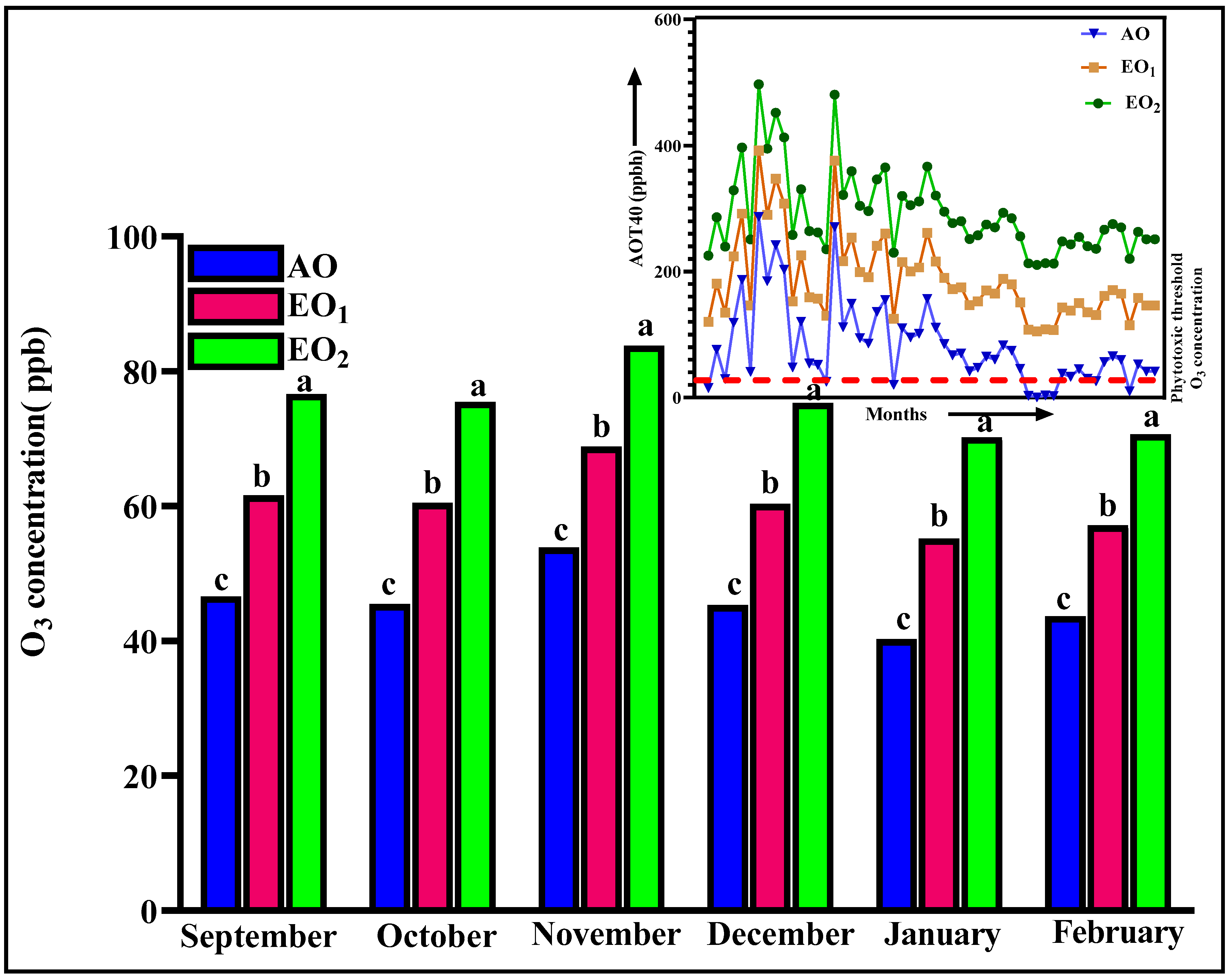
3.1. Ozone Monitoring and AOT 40
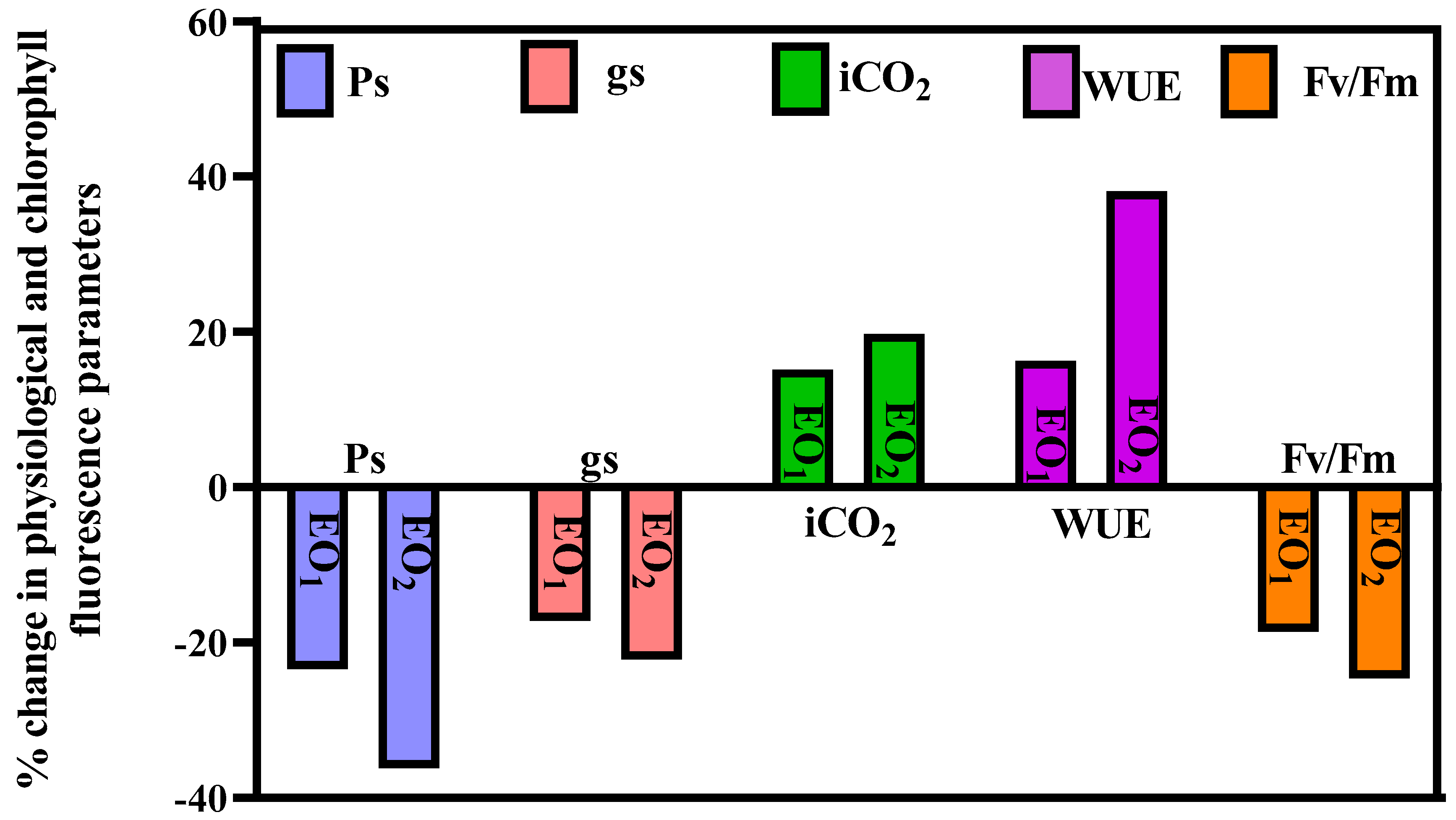
3.2. Gas Exchange and Chlorophyll Fluorescence
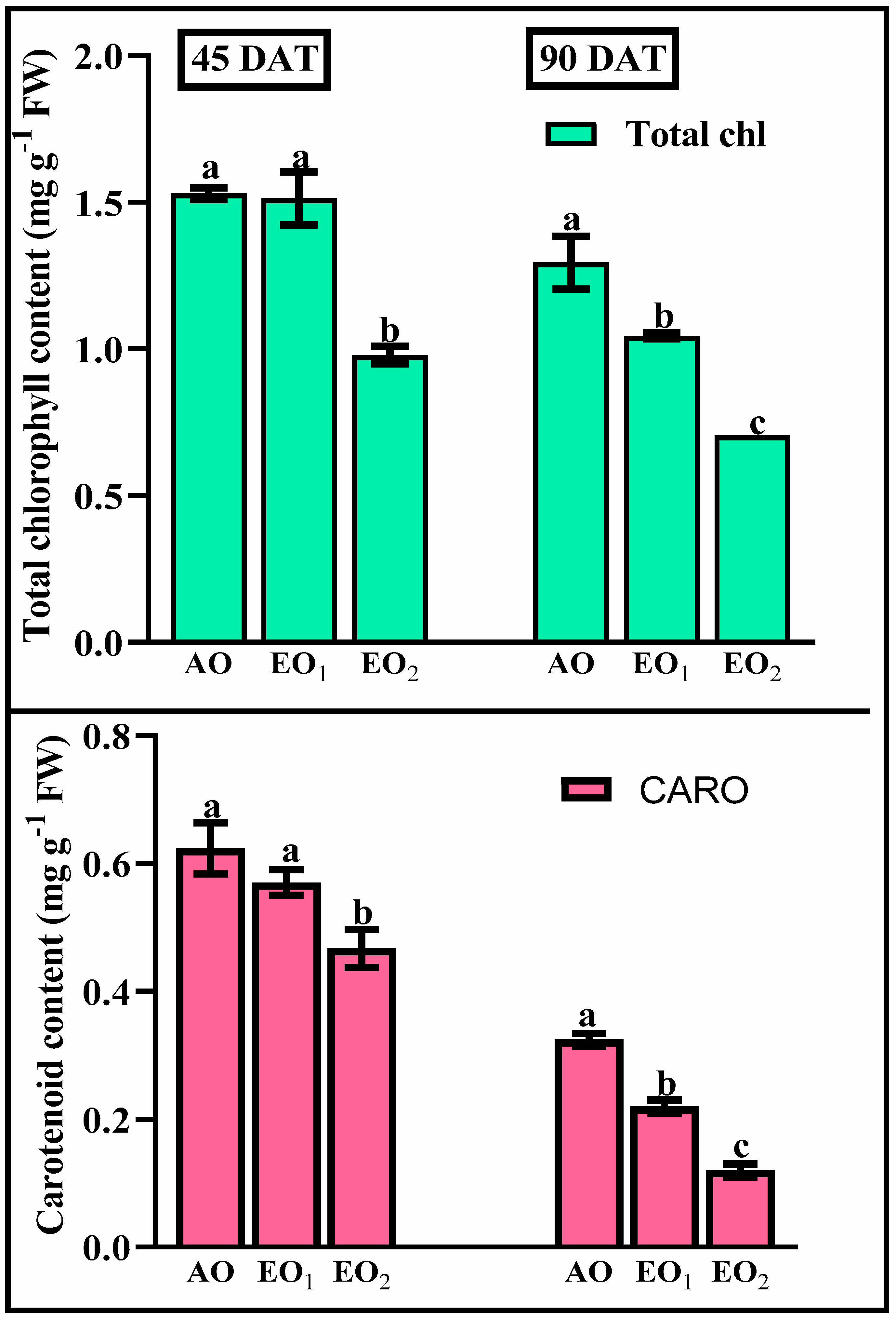
3.3. Photosynthetic Pigments
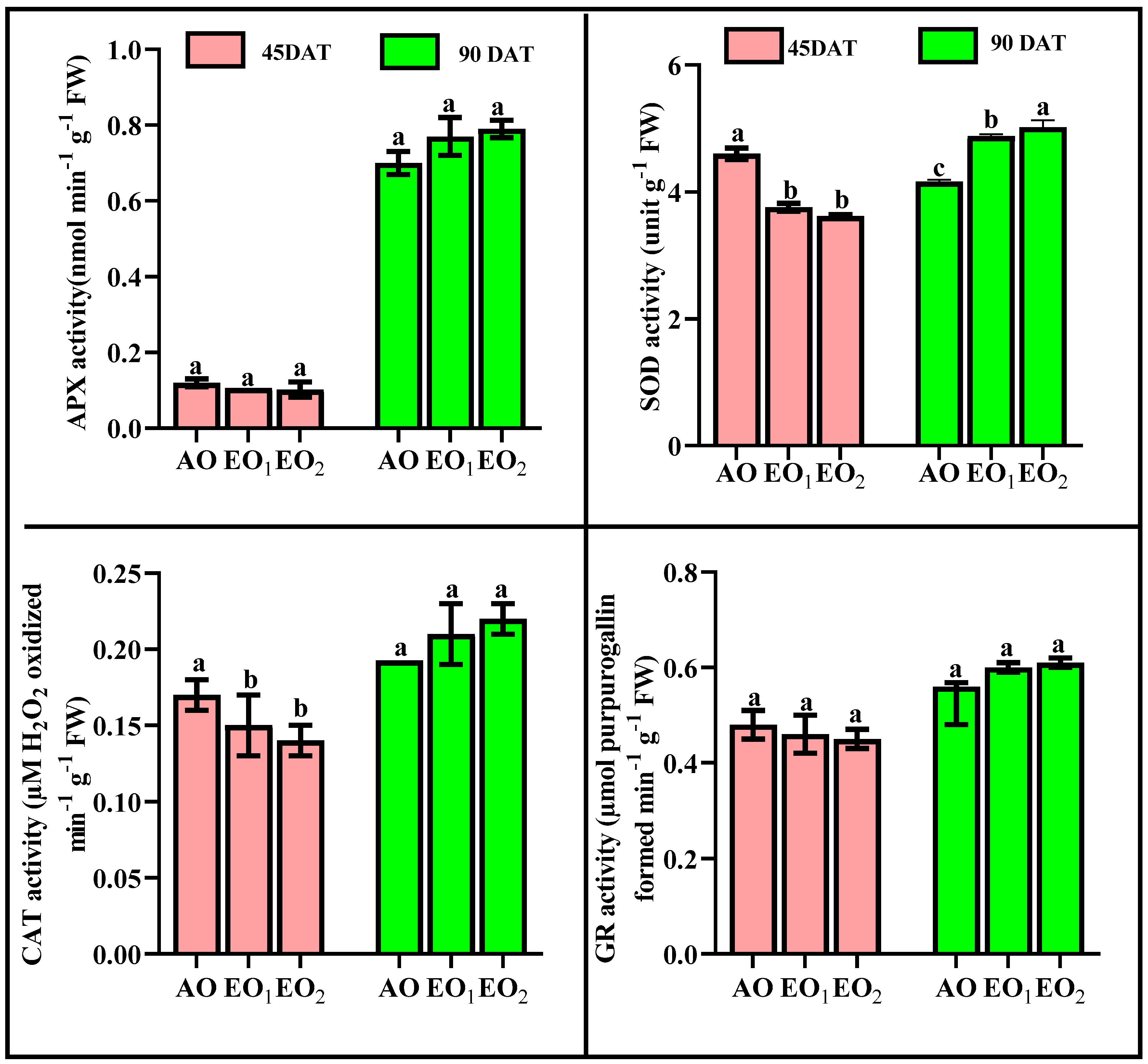
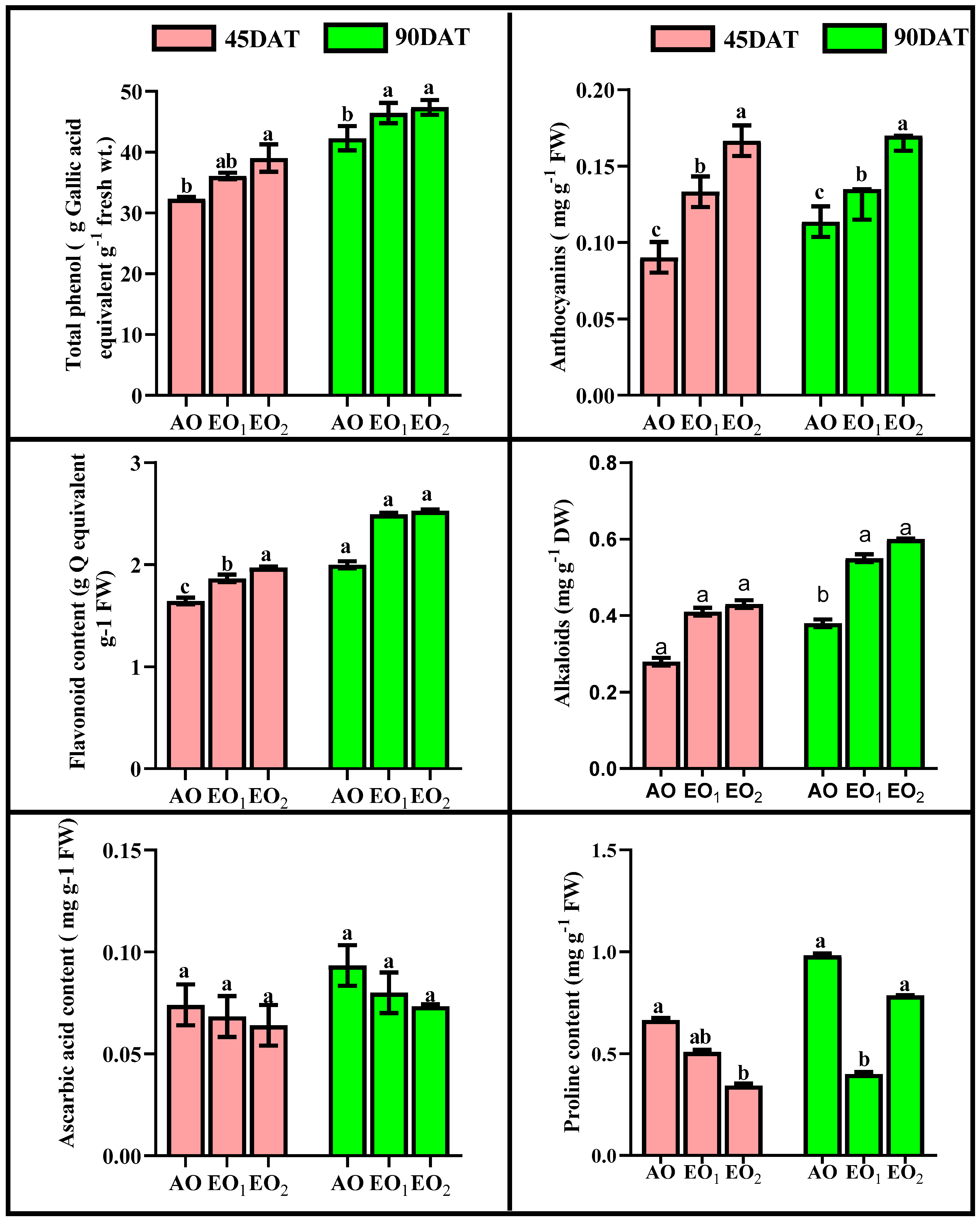
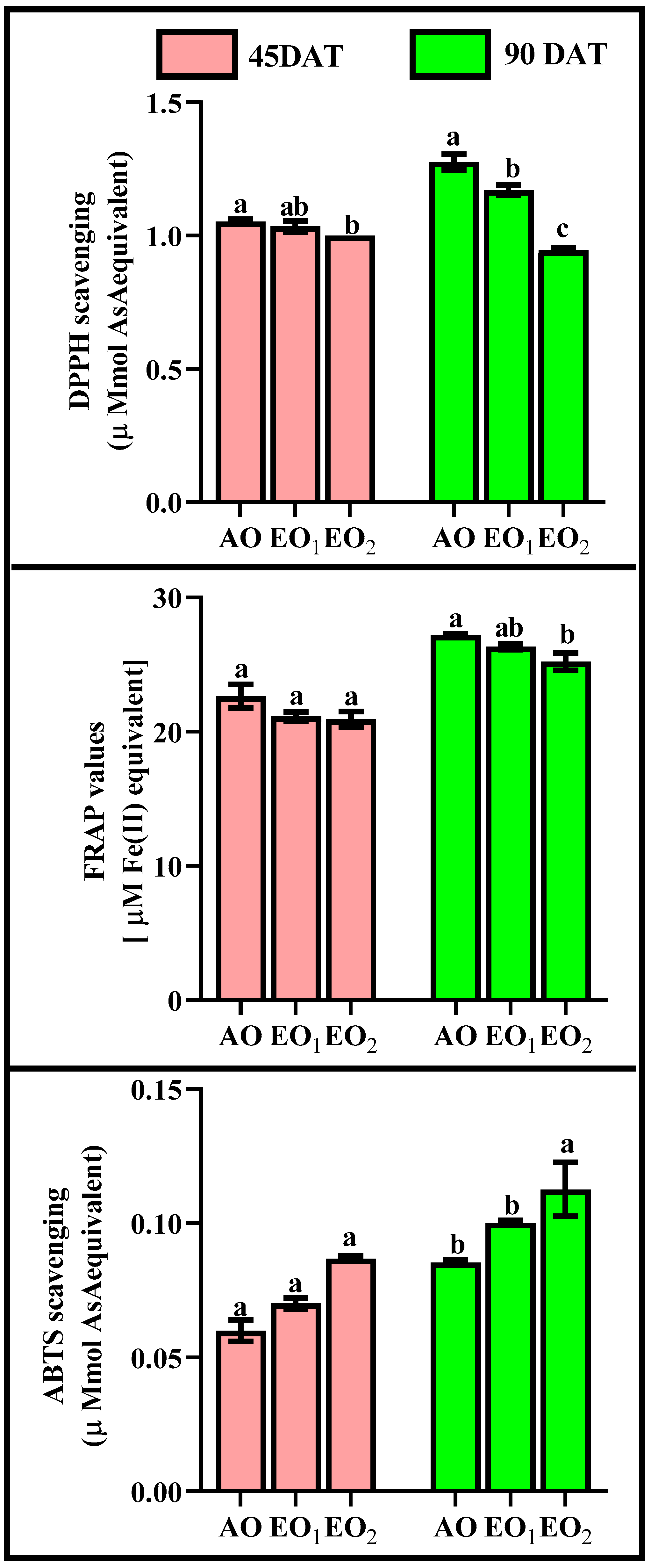
3.4. Biochemical Assays
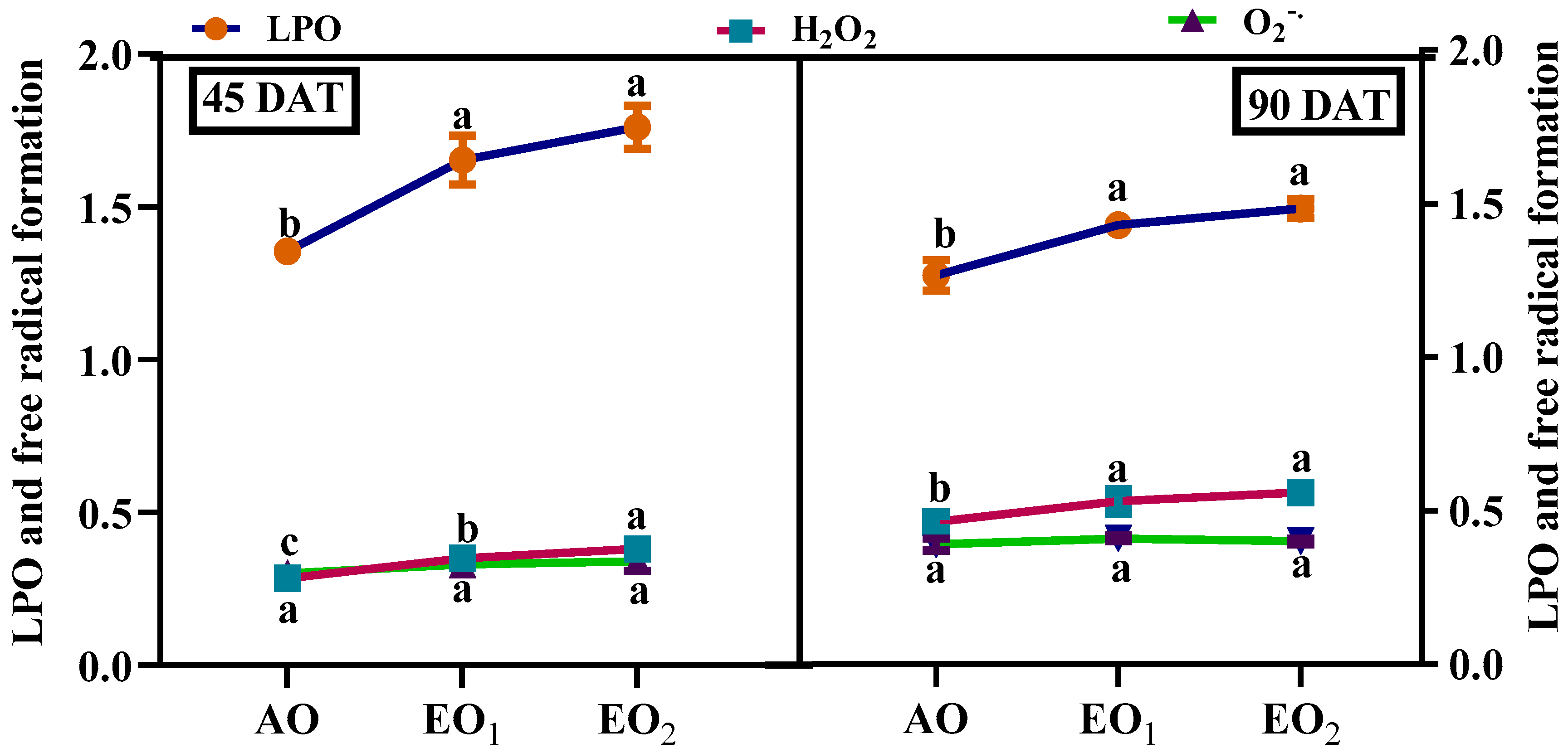
3.4.1. Hydrogen Peroxide (H2O2), Superoxide Radicals (O2o−) and MDA Contents
3.4.2. Antioxidants and Metabolites
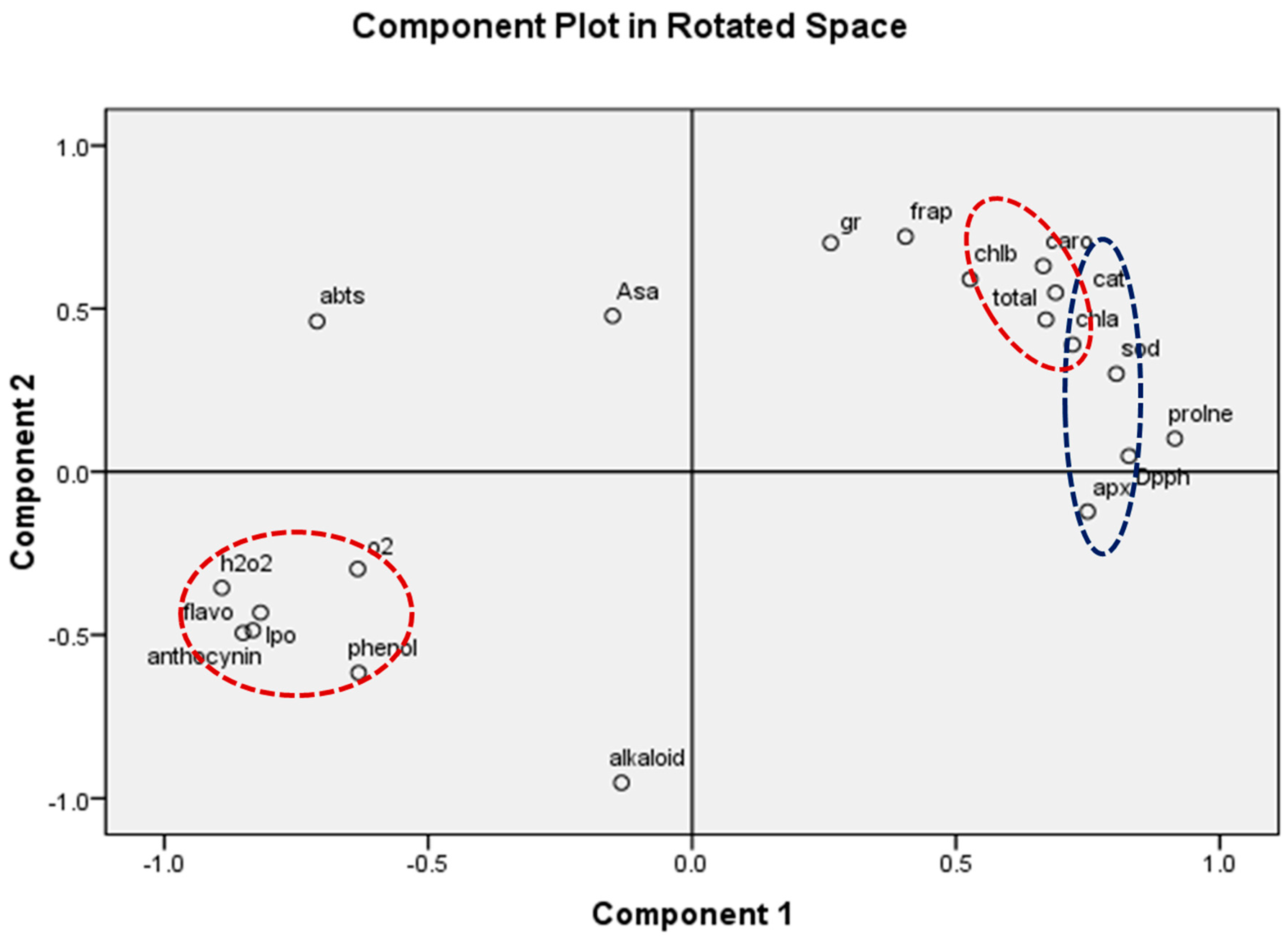
3.4.3. ROS Scavenging Potential
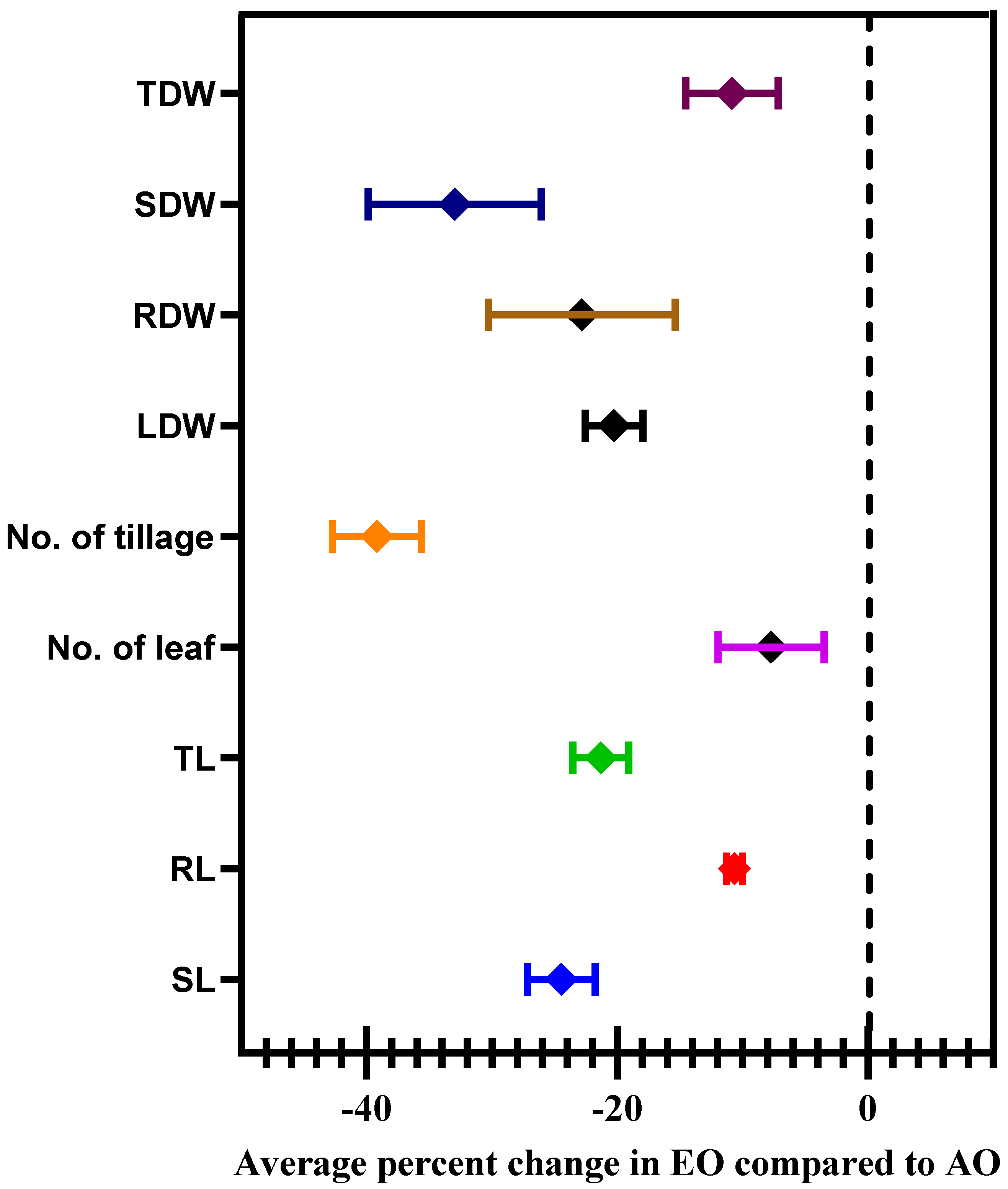
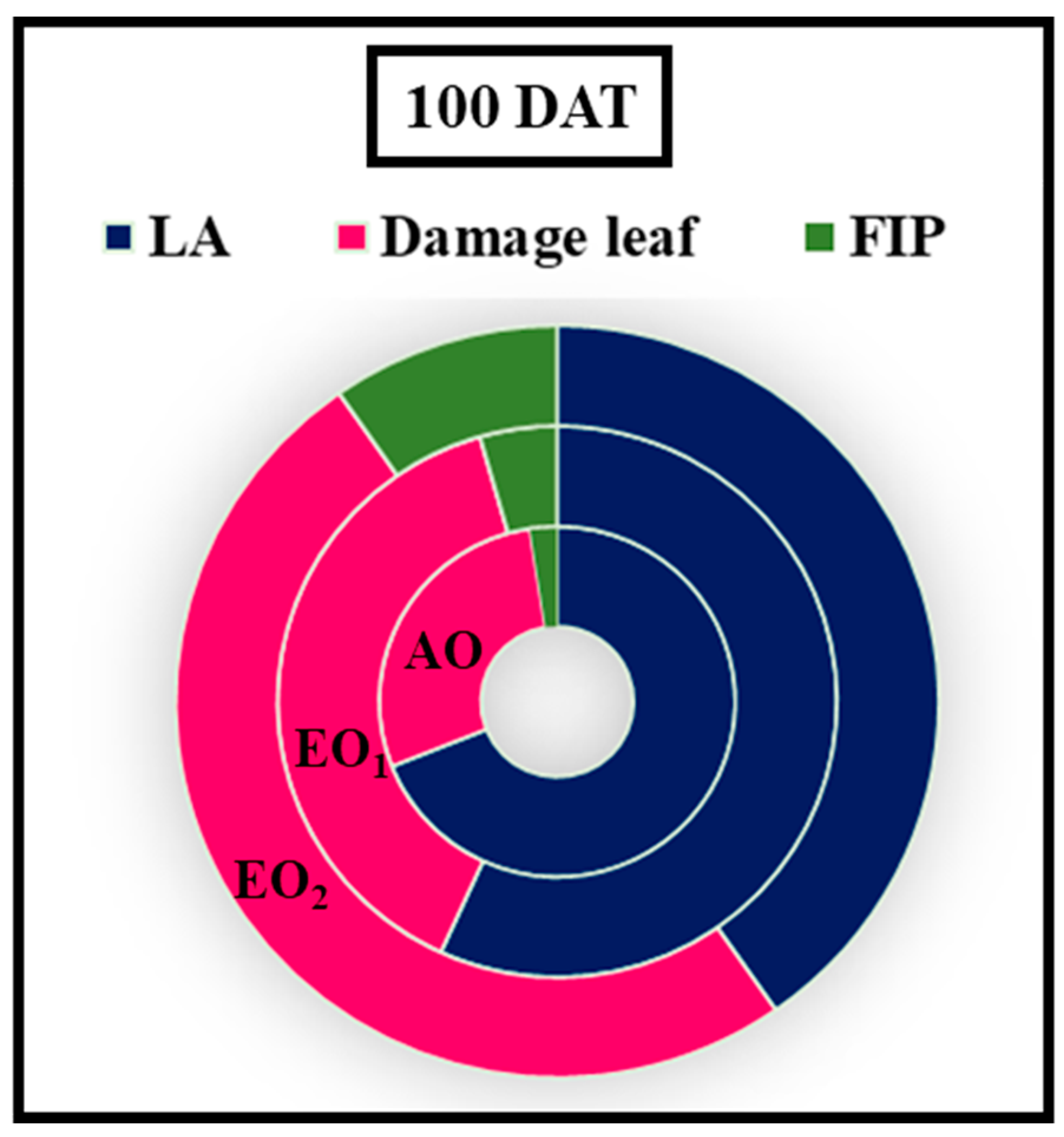
3.5. Morphological Parameters
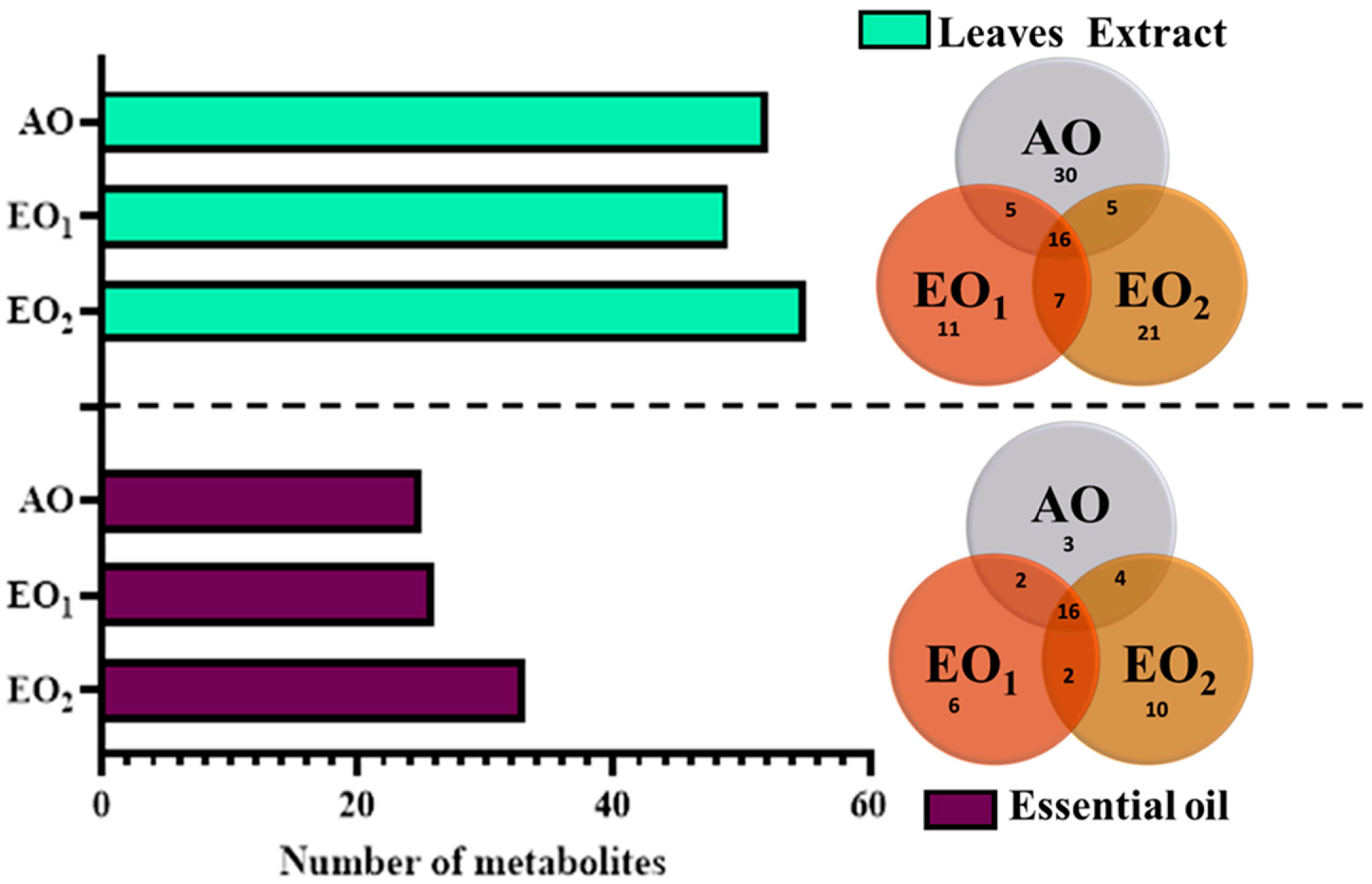
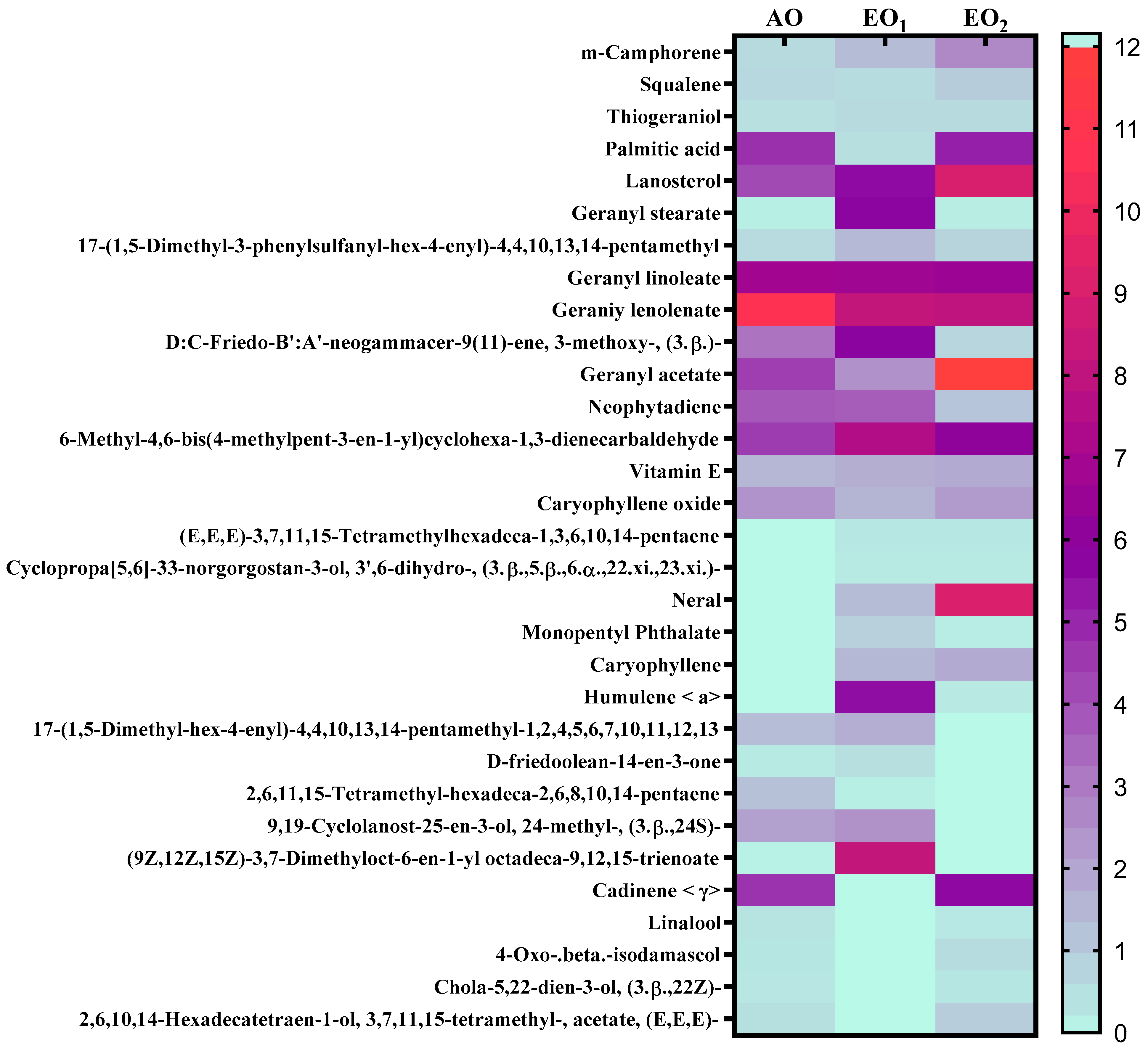
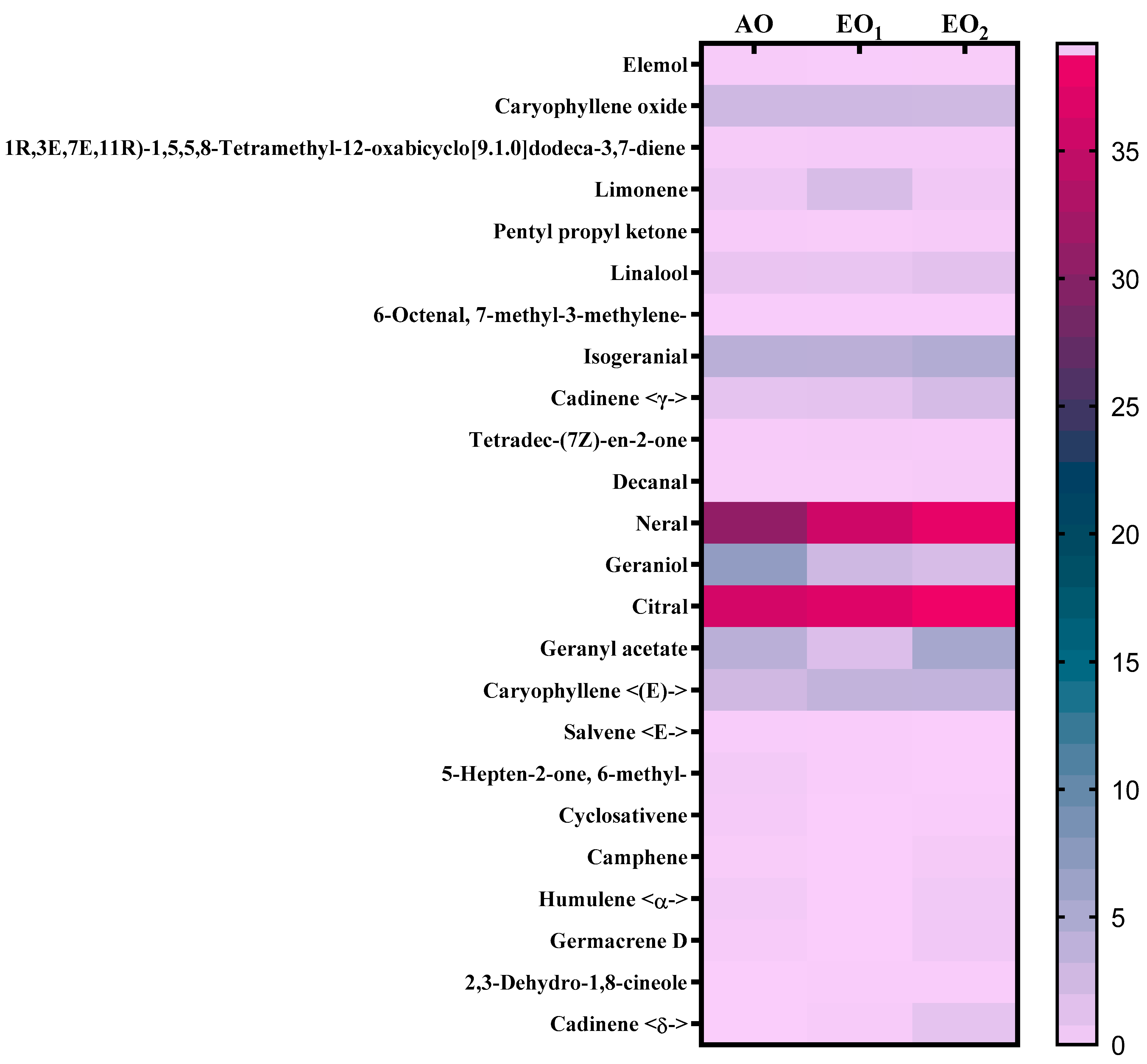
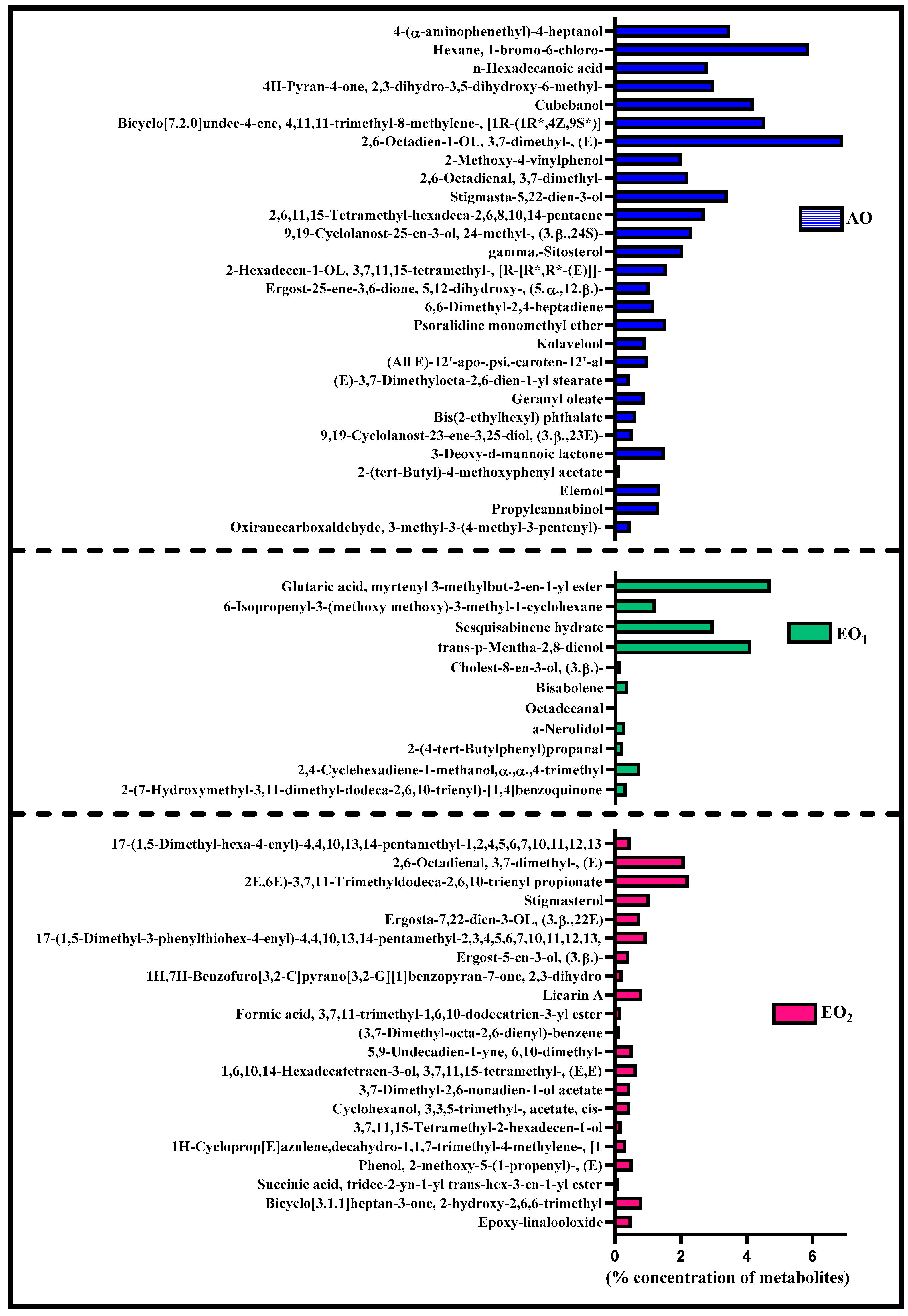
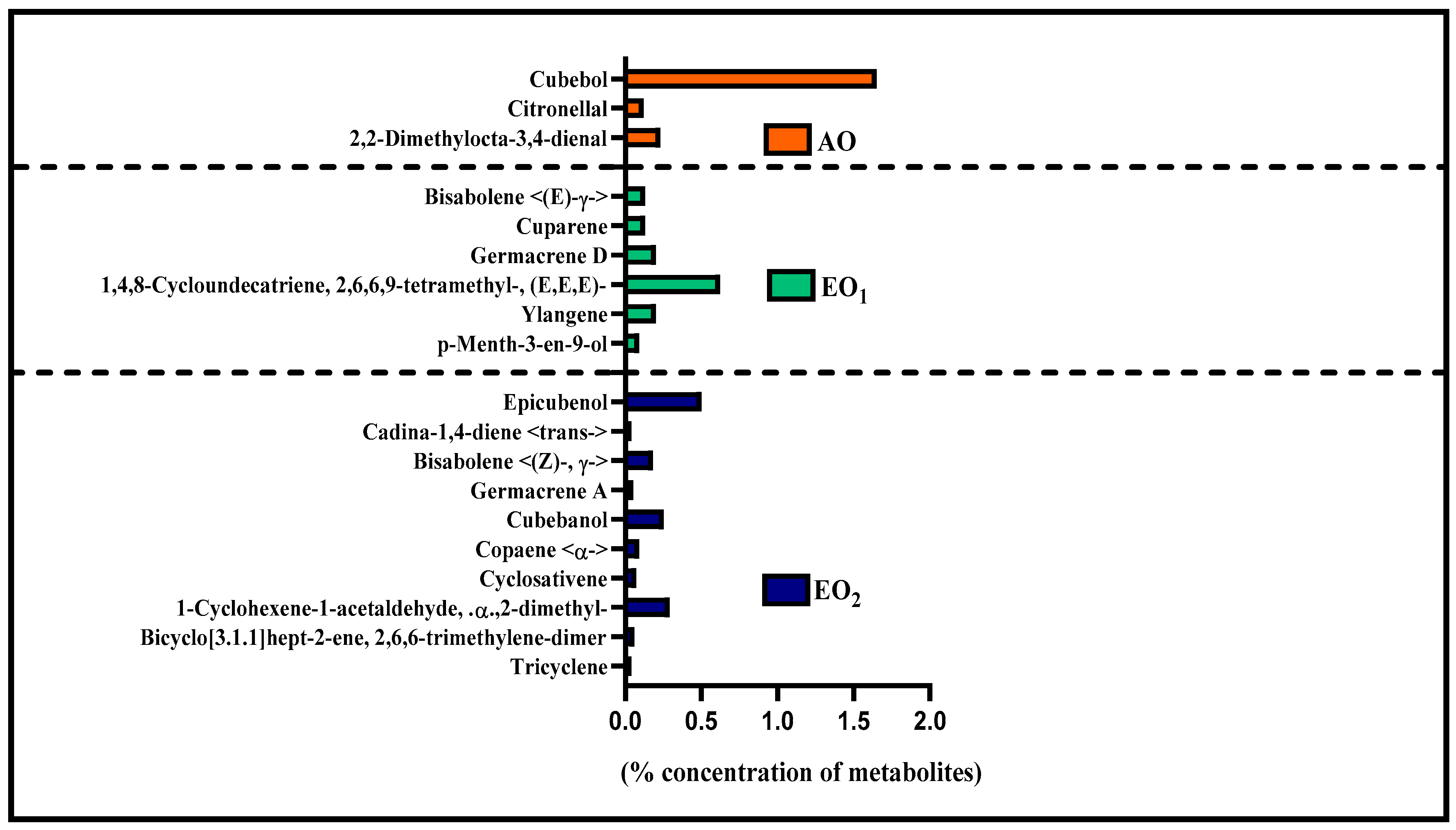
3.6. Effects on Metabolite Constituents in Leaves Extract and Essential Oil
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Tiwari, S.; Rai, R.; Agrawal, M. Annual and seasonal variations in tropospheric ozone concentrations around Varanasi. Int. J. Remote Sens. 2008, 29, 4499–4514. [Google Scholar] [CrossRef]
- Harmens, H.; Hayes, F.; Sharps, K.; Radbourne, A.; Mills, G. Can reduced irrigation mitigate ozone impacts on an ozone-sensitive African wheat variety. Plants 2019, 8, 220. [Google Scholar] [CrossRef] [PubMed]
- IPCC. Climate change 2007: The physical science basis. In Contribution of Working Group I to the Fourth Assessment Report of the Intergovernmental Panel on Climate Change; Solomon, S., Qin, D., Manning, M., Eds.; Cambridge University Press: Cambridge, UK, 2007. [Google Scholar]
- Ansari, N.; Yadav, D.S.; Agrawal, M.; Agrawal, S.B. The impact of elevated ozone on growth, secondary metabolites, production of reactive oxygen species and antioxidant response in an anti-diabetic plant Costus pictus. Funct. Plant Biol. 2021, 48, 597–610. [Google Scholar] [CrossRef] [PubMed]
- Manisalidis, I.; Stavropoulou, E.; Stavropoulos, A.; Bezirtzoglou, E. Environmental and health impacts of air pollution: A review. Front. Public Health 2020, 8, 14. [Google Scholar] [CrossRef] [PubMed]
- Madheshiya, P.; Gupta, G.S.; Sahoo, A.; Tiwari, S. Biomonitoring tools and bioprogramming: An overview. In New Paradigms in Environmental Biomonitoring Using Plants; Elsevier: Amsterdam, The Netherlands, 2022; pp. 341–366. [Google Scholar] [CrossRef]
- Sicard, P.; Anav, A.; de Marco, A.; Paoletti, E. Projected global ground-level ozone impacts on vegetation under different emission and climate scenarios. Atmos. Chem. Phys. 2017, 17, 12177–12196. [Google Scholar] [CrossRef]
- Kumari, S.; Lakhani, A.; Kumari, K.M. First observation-based study on surface O3 trend in Indo-Gangetic Plain: Assessment of its impact on crop yield. Chemosphere 2020, 255, 126972. [Google Scholar] [CrossRef]
- Kumar, A.; Jnanesha, A.C.; Lal, R.K. Coppicing impact on the essential oil yield and its chemical composition of lemongrass cultivars of the genus Cymbopogon under the semi-arid region of South India. Acta Ecol. Sin. 2023, 43, 20–26. [Google Scholar] [CrossRef]
- Ojha, N.; Pozzer, A.; Rauthe-Schöch, A.; Baker, A.K.; Yoon, J.; Brenninkmeijer, C.A.; Lelieveld, J. Ozone and carbon monoxide over India during the summer monsoon: Regional emissions and transport. Atmos. Chem. Phys. 2016, 16, 3013–3032. [Google Scholar] [CrossRef]
- Mukherjee, A.; Yadav, D.S.; Agrawal, S.B.; Agrawal, M. Ozone a persistent challenge to food security in India: Current status and policy implications. Curr. Opin. Environ. Sci. Health 2021, 19, 100220. [Google Scholar] [CrossRef]
- Boleti, E.; Hueglin, C.; Grange, S.K.; Prévôt, A.S.; Takahama, S. Temporal and spatial analysis of ozone concentrations in Europe based on timescale decomposition and a multi-clustering approach. Atmos. Chem. Phys. 2020, 20, 9051–9066. [Google Scholar] [CrossRef]
- Mitchell, M.; Wiacek, A.; Ashpole, I. Surface ozone in the North American pollution outflow region of Nova Scotia: Long-term analysis of surface concentrations, precursor emissions and long-range transport influence. Atmos. Environ. 2021, 261, 118536. [Google Scholar] [CrossRef]
- Cooper, O.R.; Parrish, D.D.; Ziemke, J.; Balashov, N.V.; Cupeiro, M.; Galbally, I.E.; Gilge, S.; Horowitz, L.; Jensen, N.R.; Lamarque, J.F.; et al. Global distribution and trends of tropospheric ozone: An observation-based review Global distribution and trends of tropospheric ozone. Elem. Sci. Anthr. 2014, 2, 000029. [Google Scholar] [CrossRef]
- Tiwari, S.; Agrawal, M. Tropospheric Ozone and Its Impacts on Crop Plants; Springer International Publishing AG: Berlin, Germany, 2018; Volume 1007. [Google Scholar]
- Zhan, Y.; Luo, Y.; Deng, X.; Grieneisen, M.L.; Zhang, M.; Di, B. Spatiotemporal prediction of daily ambient ozone levels across China using random forest for human exposure assessment. Environ. Pollut. 2018, 233, 464–473. [Google Scholar] [CrossRef] [PubMed]
- Fatima, A.; Singh, A.A.; Mukherjee, A.; Agrawal, M.; Agrawal, S.B. Variability in defence mechanism operating in three wheat cultivars having different levels of sensitivity against elevated ozone. Environ. Exp. Bot. 2018, 155, 66–78. [Google Scholar] [CrossRef]
- Gupta, G.S.; Tiwari, S. Role of antioxidant pool in management of ozone stress through soil nitrogen amendments in two cultivars of a tropical legume. Funct. Plant Biol. 2020, 48, 371–385. [Google Scholar] [CrossRef]
- Hassan, A. Effects of mineral nutrients on physiological and biochemical processes related to secondary metabolites production in medicinal herbs. Med Arom Plant Sci Biotechnol. 2012, 6, 105. [Google Scholar]
- Elshafie, H.S.; Camele, I.; Mohamed, A.A. A Comprehensive Review on the Biological, Agricultural and Pharmaceutical Properties of Secondary Metabolites Based-Plant Origin. Int. J. Mol. Sci. 2023, 24, 3266. [Google Scholar] [CrossRef]
- George, B.P.; Chandran, R.; Abrahamse, H. Role of phytochemicals in cancer chemoprevention: Insights. Antioxidants 2021, 10, 1455. [Google Scholar] [CrossRef]
- Sharma, A. Gene expression analysis in medicinal plants under abiotic stress conditions. In Plant Metabolites and Regulation under Environmental Stress; Academic Press: Cambridge, MA, USA, 2018; pp. 407–414. [Google Scholar] [CrossRef]
- Bortolin, R.C.; Caregnato, F.F.; Junior, A.M.; Zanotto-Filho, A.; Moresco, K.S.; de Oliveira Rios, A.; de Oliveira Salvi, A.; Ortmann, C.F.; de Carvalho, P.; Reginatto, F.H.; et al. Chronic ozone exposure alters the secondary metabolite profile, antioxidant potential, anti-inflammatory property, and quality of red pepper fruit from Capsicum baccatum. Ecotoxicol. Environ. Saf. 2016, 129, 16–24. [Google Scholar] [CrossRef]
- Pant, P.; Pandey, S.; Dall’Acqua, S. The influence of environmental conditions on secondary metabolites in medicinal plants: A literature review. Chem. Biodivers. 2021, 18, e2100345. [Google Scholar] [CrossRef]
- Ansari, N.; Agrawal, M.; Agrawal, S.B. An assessment of growth, floral morphology, and metabolites of a medicinal plant Sida cordifolia L. under the influence of elevated ozone. Environ. Sci. Pollut. Res. 2021, 28, 832–845. [Google Scholar] [CrossRef] [PubMed]
- Kumar, I.; Sharma, R.K. Production of secondary metabolites in plants under abiotic stress: An overview. Significances Bioeng. Biosci. 2018, 2, 196–200. [Google Scholar] [CrossRef]
- Ainsworth, E.A. Understanding and improving global crop response to ozone pollution. Plant J. 2017, 90, 886–897. [Google Scholar] [CrossRef] [PubMed]
- Singh, S.; Singh, P.; Agrawal, S.B.; Agrawal, M. Use of Ethylenediurea (EDU) in identifying indicator cultivars of Indian clover against ambient ozone. Ecotoxicol. Environ. Saf. 2018, 147, 1046–1055. [Google Scholar] [CrossRef]
- Dumont, J.; Keski-Saari, S.; Keinänen, M.; Cohen, D.; Ningre, N.; Kontunen-Soppela, S.; Baldet, P.; Gibon, Y.; Dizengremel, P.; Vaultier, M.N.; et al. Ozone affects ascorbate and glutathione biosynthesis as well as amino acid contents in three Euramerican poplar genotypes. Tree Physiol. 2014, 34, 253–266. [Google Scholar] [CrossRef] [PubMed]
- Ashraf, M.A.; Iqbal, M.; Rasheed, R.; Hussain, I.; Riaz, M.; Arif, M.S. Environmental stress and secondary metabolites in plants: An overview. In Plant Metabolites and Regulation under Environmental Stress; Academic Press: Cambridge, MA, USA, 2018; pp. 153–167. [Google Scholar] [CrossRef]
- Ali, M.; Khan, T.; Fatima, K.; Ali, Q.U.; Ovais, M.; Khalil, A.T.; Ullah, I.; Raza, A.; Shinwari, Z.K.; Idrees, M. Selected hepatoprotective herbal medicines: Evidence from ethnomedicinal applications, animal models, and possible mechanism of actions. Phytother. Res. 2018, 32, 199–215. [Google Scholar] [CrossRef]
- Haque, A.N.; Remadevi, R.; Naebe, M. Lemongrass (Cymbopogon): A review on its structure, properties, applications and recent developments. Cellulose 2018, 25, 5455–5477. [Google Scholar] [CrossRef]
- Mukarram, M.; Choudhary, S.; Khan, M.A.; Poltronieri, P.; Khan, M.M.; Ali, J.; Kurjak, D.; Shahid, M. Lemongrass essential oil components with antimicrobial and anticancer activities. Antioxidants 2021, 11, 20. [Google Scholar] [CrossRef]
- Choudhary, S.; Zehra, A.; Mukarram, M.; Wani, K.I.; Naeem, M.; Khan, M.M.; Aftab, T. Salicylic acid-mediated alleviation of soil boron toxicity in Mentha arvensis and Cymbopogon flexuosus: Growth, antioxidant responses, essential oil contents and components. Chemosphere 2021, 276, 130153. [Google Scholar] [CrossRef]
- Joy, P.P.; Skaria, B.P.; Mathew, S.; Mathew, G.; Joseph, A.; Sreevidya, P.P. Lemongrass. Ind. J. Arecanut Spices Medicin. Plants 2006, 2, 55–64. [Google Scholar]
- Bahl, J.R.; Singh, A.K.; Lal, R.K.; Gupta, A.K. High-yielding improved varieties of medicinal and aromatic crops for enhanced income. In New Age Herbals: Resource, Quality and Pharmacognosy; Springer: Berlin/Heidelberg, Germany, 2018; pp. 247–265. [Google Scholar]
- Lal, R.K. Stability for oil yield and variety recommendations’ using AMMI (additive main effects and multiplicative interactions) model in Lemongrass (Cymbopogon species). Ind. Crops Prod. 2012, 40, 296–301. [Google Scholar] [CrossRef]
- Tiwari, S.; Agrawal, M.; Marshall, F.M. Evaluation of ambient air pollution impact on carrot plants at a sub urban site using open top chambers. Environ. Monit. Assess. 2006, 119, 15–30. [Google Scholar] [CrossRef] [PubMed]
- Mauzerall, D.L.; Wang, X. Protecting agricultural crops from the effects of tropospheric ozone exposure: Reconciling science and standard setting in the United States, Europe, and Asia. Annu. Rev. Energy Environ. 2001, 26, 237–268. [Google Scholar] [CrossRef]
- Mishra, A.K.; Rai, R.; Agrawal, S.B. Differential response of dwarf and tall tropical wheat cultivars to elevated ozone with and without carbon dioxide enrichment: Growth, yield and grain quality. Field Crops Res. 2013, 145, 21–32. [Google Scholar] [CrossRef]
- Alexieva, V.; Sergiev, I.; Mapelli, S.; Karanov, E. The effect of drought and ultraviolet radiation on growth and stress markers in pea and wheat. Plant Cell Environ. 2001, 24, 1337–1344. [Google Scholar] [CrossRef]
- Elstner, E.F.; Heupel, A. Inhibition of nitrite formation from hydroxyl ammonium chloride: A simple assay for superoxide dismutase. Anal. Biochem. 1976, 70, 616–620. [Google Scholar] [CrossRef]
- Heath, R.L.; Packer, L. Photoperoxidation in isolated chloroplasts: I. Kinetics and stoichiometry of fatty acid peroxidation. Arch. Biochem. Biophys. 1968, 125, 189–198. [Google Scholar] [CrossRef]
- Fridovich, I. Superoxide dismutases. Adv. Enzymol. Relat. Areas Mol. Biol. 1974, 41, 35–97. [Google Scholar]
- Anderson, M.E. Glutathione. In Free Radicals: A practical Approach; Punchard, N.A., Kelly, F.J., Eds.; Oxford University Press: Oxford, UK, 1996; pp. 213–226. [Google Scholar]
- Nakano, Y.; Asada, K. Hydrogen peroxide is scavenged by ascorbate-specific peroxidase in spinach chloroplasts. Plant Cell Physiol. 1981, 22, 867–880. [Google Scholar] [CrossRef]
- Aebi, H. [13] Catalase in vitro. In Methods in Enzymology; Academic Press: Cambridge, MA, USA, 1984; Volume 105, pp. 121–126. [Google Scholar] [CrossRef]
- Bray, H.G.; Thorpe, W. Analysis of phenolic compounds of interest in metabolism. In Methods of Biochemical Analysis; Interscience Publishers, Inc.: Newy York, NY, USA, 1954; pp. 27–52. [Google Scholar] [CrossRef]
- Flint, S.D.; Jordan, P.W.; Caldwell, M.M. Plant protective response to enhanced UV-B radiation under field conditions: Leaf optical properties and photosynthesis. Photochem. Photobiol. 1985, 41, 95–99. [Google Scholar] [CrossRef]
- Gitelson, A.A.; Merzlyak, M.N.; Chivkunova, O.B. Optical properties and non-destructive estimation of anthocyanin content in plant leaves. Photochem. Photobiol. 2001, 74, 38–45. [Google Scholar] [CrossRef] [PubMed]
- Jamil, M.; Bashir, S.A.; Anwar, S.; Bibi, S.; Bangash, A.; Ullah, F.; Rha, E.S. Effect of salinity on physiological and biochemical characteristics of different varieties of rice. Pak. J. Bot. 2012, 44, 7–13. [Google Scholar]
- Takshak, S.; Agrawal, S.Á. Secondary metabolites and phenylpropanoid pathway enzymes as influenced under supplemental ultraviolet-B radiation in Withania somnifera Dunal, an indigenous medicinal plant. J. Photochem. Photobiol. B Biol. 2014, 140, 332–343. [Google Scholar] [CrossRef] [PubMed]
- Keller, T.; Schwager, H. Air pollution and ascorbic acid. Eur. J. For. Pathol. 1977, 7, 338–350. [Google Scholar] [CrossRef]
- Noreen, H.; Semmar, N.; Farman, M.; McCullagh, J.S. Measurement of total phenolic content and antioxidant activity of aerial parts of medicinal plant Coronopus didymus. Asian Pac. J. Trop. Med. 2017, 10, 792–801. [Google Scholar] [CrossRef]
- Khatua, S.; Acharya, K. Functional ingredients and medicinal prospects of ethanol extract from Macrocybe lobayensis. Pharmacogn. J. 2018, 10. [Google Scholar] [CrossRef]
- Clarke, G.; Ting, K.N.; Wiart, C.; Fry, J. High correlation of 2, 2-diphenyl-1-picrylhydrazyl (DPPH) radical scavenging, ferric reducing activity potential and total phenolics content indicates redundancy in use of all three assays to screen for antioxidant activity of extracts of plants from the Malaysian rainforest. Antioxidants 2013, 2, 1–10. [Google Scholar] [CrossRef]
- Maxwell, K.; Johnson, G.N. Chlorophyll fluorescence—A practical guide. J. Exp. Bot. 2000, 51, 659–668. [Google Scholar] [CrossRef]
- Takshak, S.; Agrawal, S.B. Interactive effects of supplemental ultraviolet-B radiation and indole-3-acetic acid on Coleus forskohlii Briq.: Alterations in morphological-, physiological-, and biochemical characteristics and essential oil content. Ecotoxicol. Environ. Saf. 2018, 147, 313–326. [Google Scholar] [CrossRef]
- Sebastian, A.; Kumari, R.; Kiran, B.R.; Prasad, M.N. Ultraviolet B induced bioactive changes of enzymatic and non-enzymatic antioxidants and lipids in L. (Fenugreek). Euro Biotech J. 2018, 2, 64–71. [Google Scholar] [CrossRef]
- Takshak, S.; Agrawal, S.B. Defence strategies adopted by the medicinal plant Coleus forskohlii against supplemental ultraviolet-B radiation: Augmentation of secondary metabolites and antioxidants. Plant Physiol. Biochem. 2015, 97, 124–138. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, A.; Agrawal, M.; Agrawal, S.B. Effect of water deficit stress on an Indian wheat cultivar (Triticum aestivum L. HD 2967) under ambient and elevated level of ozone. Sci. Total Environ. 2020, 714, 136837. [Google Scholar] [CrossRef] [PubMed]
- Yadav, D.S.; Rai, R.; Mishra, A.K.; Chaudhary, N.; Mukherjee, A.; Agrawal, S.B.; Agrawal, M. ROS production and its detoxification in early and late sown cultivars of wheat under future O3 concentration. Sci. Total Environ. 2019, 659, 200–210. [Google Scholar] [CrossRef] [PubMed]
- Sarkar, A.; Singh, A.A.; Agrawal, S.B.; Ahmad, A.; Rai, S.P. Cultivar specific variations in antioxidative defense system, genome and proteome of two tropical rice cultivars against ambient and elevated ozone. Ecotoxicol. Environ. Saf. 2015, 115, 101–111. [Google Scholar] [CrossRef] [PubMed]
- Ansari, N.; Yadav, D.S.; Singh, P.; Agrawal, M.; Agrawal, S.B. Ozone exposure response on physiological and biochemical parameters vis-a-vis secondary metabolites in a traditional medicinal plant Sida cordifolia L. Ind. Crops Prod. 2023, 194, 116267. [Google Scholar] [CrossRef]
- Sahoo, A.; Madheshiya, P.; Mishra, A.K.; Tiwari, S. Combating ozone stress through N fertilization: A case study of Indian bean (Dolichos lablab L.). Front. Plant Sci. 2023, 14, 548. [Google Scholar] [CrossRef]
- Rathore, D.; Chaudhary, I.J. Ozone risk assessment of castor (Ricinus communis L.) cultivars using open top chamber and ethylene di urea (EDU). Environ. Pollut. 2019, 244, 257–269. [Google Scholar] [CrossRef]
- Singh, A.A.; Agrawal, S.B.; Shahi, J.P.; Agrawal, M. Assessment of growth and yield losses in two Zea mays L. cultivars (quality protein maize and nonquality protein maize) under projected levels of ozone. Environ. Sci. Pollut. Res. 2014, 21, 2628–2641. [Google Scholar] [CrossRef]
- Wang, Y.; Yang, L.; Höller, M.; Zaisheng, S.; Pariasca-Tanaka, J.; Wissuwa, M.; Frei, M. Pyramiding of ozone tolerance QTLs OzT8 and OzT9 confers improved tolerance to season-long ozone exposure in rice. Environ. Exp. Bot. 2014, 104, 26–33. [Google Scholar] [CrossRef]
- Tiwari, S.; Agrawal, M. Assessment of the variability in response of radish and brinjal at biochemical and physiological levels under similar ozone exposure conditions. Environ. Monit. Assess. 2011, 175, 443–454. [Google Scholar] [CrossRef]
- Chaudhary, I.J.; Rathore, D. Assessment of dose-response relationship between ozone dose and groundnut (Arachis hypogaea L) cultivars using Open Top Chamber (OTC) and Ethylenediurea (EDU). Environ. Technol. Innov. 2021, 22, 101494. [Google Scholar] [CrossRef]
- Singh, R.; Rathore, D. Oxidative stress defence responses of wheat (Triticum aestivum L.) and chilli (Capsicum annum L.) cultivars grown under textile effluent fertilization. Plant Physiol. Biochem. 2018, 123, 342–358. [Google Scholar] [CrossRef] [PubMed]
- You, J.; Chan, Z. ROS regulation during abiotic stress responses in crop plants. Front. Plant Sci. 2015, 6, 1092. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Zong, M.H.; Fu, S.Y.; Li, N. Correlation between physicochemical properties and enzymatic digestibility of rice straw pretreated with cholinium ionic liquids. ACS Sustain. Chem. Eng. 2016, 4, 4340–4345. [Google Scholar] [CrossRef]
- Kumari, R.; Agrawal, S.B. Supplemental UV-B induced changes in leaf morphology, physiology and secondary metabolites of an Indian aromatic plant Cymbopogon citratus (DC) Staph under natural field conditions. Int. J. Environ. Stud. 2010, 67, 655–675. [Google Scholar] [CrossRef]
- Yadav, D.S.; Jaiswal, B.; Agrawal, S.B.; Agrawal, M. Diurnal variations in physiological characteristics, photo assimilates, and total ascorbate in early and late sown Indian wheat cultivars under exposure to elevated ozone. Atmosphere 2021, 12, 1568. [Google Scholar] [CrossRef]
- Singh, E.; Tiwari, S.; Agrawal, M. Effects of elevated ozone on photosynthesis and stomatal conductance of two soybean varieties: A case study to assess impacts of one component of predicted global climate change. Plant Biol. 2009, 11, 101–108. [Google Scholar] [CrossRef]
- Nishiyama, Y.; Allakhverdiev, S.I.; Murata, N. Protein synthesis is the primary target of reactive oxygen species in the photoinhibition of photosystem, I.I. Physiol. Plant. 2011, 142, 35–46. [Google Scholar] [CrossRef]
- Tiwari, S.; Agrawal, M.; Marshall, F.M. Seasonal variations in adaptational strategies of Beta vulgaris L. plants in response to ambient air pollution: Biomass allocation, yield and nutritional quality. Trop. Ecol. 2010, 51, 353–363. [Google Scholar]
- Qaderi, M.M.; Martel, A.B.; Strugnell, C.A. Environmental Factors Regulate Plant Secondary Metabolites. Plants 2023, 12, 447. [Google Scholar] [CrossRef]
- Jan, R.; Asaf, S.; Numan, M.; Kim, K.M. Plant secondary metabolite biosynthesis and transcriptional regulation in response to biotic and abiotic stress conditions. Agronomy 2021, 11, 968. [Google Scholar] [CrossRef]
- Lin, D.; Xiao, M.; Zhao, J.; Li, Z.; Xing, B.; Li, X.; Kong, M.; Li, L.; Zhang, Q.; Liu, Y.; et al. An overview of plant phenolic compounds and their importance in human nutrition and management of type 2 diabetes. Molecules 2016, 21, 1374. [Google Scholar] [CrossRef]
- Feng, Z.; De Marco, A.; Anav, A.; Gualtieri, M.; Sicard, P.; Tian, H.; Fornasier, F.; Tao, F.; Guo, A.; Paoletti, E. Economic losses due to ozone impacts on human health, forest productivity and crop yield across China. Environ. Int. 2019, 131, 104966. [Google Scholar] [CrossRef]
- Gil, M.; Pontin, M.; Berli, F.; Bottini, R.; Piccoli, P. Metabolism of terpenes in the response of grape (Vitis vinifera L.) leaf tissues to UV-B radiation. Phytochemistry 2012, 77, 89–98. [Google Scholar] [CrossRef]
- Pandey, N.; Pandey-Rai, S. Modulations of physiological responses and possible involvement of defense-related secondary metabolites in acclimation of Artemisia annua L. against short-term UV-B radiation. Planta 2014, 240, 611–627. [Google Scholar] [CrossRef]
- Smith, W.E.; Shivaji, R.; Williams, W.P.; Luthe, D.S.; Sandoya, G.V.; Smith, C.L.; Sparks, D.L.; Brown, A.E. A maize line resistant to herbivory constitutively releases (E)-β-caryophyllene. J. Econ. Entomol. 2012, 105, 120–128. [Google Scholar] [CrossRef] [PubMed]
- Valkama, E.; Koricheva, J.; Oksanen, E. Effects of elevated O3, alone and in combination with elevated CO2, on tree leaf chemistry and insect herbivore performance: A meta-analysis. Glob. Change Biol. 2007, 13, 184–201. [Google Scholar] [CrossRef]
- Kangasjärvi, J.; Jaspers, P.; Kollist, H. Signalling and cell death in ozone-exposed plants. Plant Cell Environ. 2005, 28, 1021–1036. [Google Scholar] [CrossRef]
- Kasote, D.M.; Katyare, S.S.; Hegde, M.V.; Bae, H. Significance of antioxidant potential of plants and its relevance to therapeutic applications. Int. J. Biol. Sci. 2015, 11, 982. [Google Scholar] [CrossRef]
- Silva, F.L.; Vieira, L.G.; Ribas, A.F.; Moro, A.L.; Neris, D.M.; Pacheco, A.C. Proline accumulation induces the production of total phenolics in transgenic tobacco plants under water deficit without increasing the G6PDH activity. Theor. Exp. Plant Physiol. 2018, 30, 251–260. [Google Scholar] [CrossRef]
- Debona, D.; Rodrigues, F.A.; Datnoff, L.E. Silicon’s role in abiotic and biotic plant stresses. Annu. Rev. Phytopathol. 2017, 55, 85–107. [Google Scholar] [CrossRef] [PubMed]
- Deng, G.B.; Zhang, H.B.; Xue, H.F.; Chen, S.N.; Chen, X.L. Chemical composition and biological activities of essential oil from the rhizomes of Iris bulleyana. Agric. Sci. China 2009, 8, 691–696. [Google Scholar] [CrossRef]
- Videira, R.; Castanheira, P.; Grãos, M.; Salgueiro, L.; Faro, C.; Cavaleiro, C. A necrodane monoterpenoid from Lavandula luisieri essential oil as a cell-permeable inhibitor of BACE-1, the β-secretase in Alzheimer’s disease. Flavour Fragr. J. 2013, 28, 380–388. [Google Scholar] [CrossRef]















Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Madheshiya, P.; Gupta, G.S.; Sahoo, A.; Tiwari, S. Role of Elevated Ozone on Development and Metabolite Contents of Lemongrass [Cymbopogon flexuosus (Steud.) (Wats.)]. Metabolites 2023, 13, 597. https://doi.org/10.3390/metabo13050597
Madheshiya P, Gupta GS, Sahoo A, Tiwari S. Role of Elevated Ozone on Development and Metabolite Contents of Lemongrass [Cymbopogon flexuosus (Steud.) (Wats.)]. Metabolites. 2023; 13(5):597. https://doi.org/10.3390/metabo13050597
Chicago/Turabian StyleMadheshiya, Parvati, Gereraj Sen Gupta, Ansuman Sahoo, and Supriya Tiwari. 2023. "Role of Elevated Ozone on Development and Metabolite Contents of Lemongrass [Cymbopogon flexuosus (Steud.) (Wats.)]" Metabolites 13, no. 5: 597. https://doi.org/10.3390/metabo13050597
APA StyleMadheshiya, P., Gupta, G. S., Sahoo, A., & Tiwari, S. (2023). Role of Elevated Ozone on Development and Metabolite Contents of Lemongrass [Cymbopogon flexuosus (Steud.) (Wats.)]. Metabolites, 13(5), 597. https://doi.org/10.3390/metabo13050597